

4 курс / Общая токсикология (доп.) / Toxikologia_ryb_Lukyanenko
.pdfВо второй части опытов мы решили детально сопоставить изменение устойчивости сеголетков и 2+ в растворах фенола различной концентрации (от 15 до 800 мг/л). Проведено по семь дублированных серий опытов. Данные этих исследований приведены в табл. 44. Почти во всех растворах более устойчивыми оказались сеголетки. Особенно показательно различие в области концентраций от 15 до 200 мг/л.
Средняя продолжительность жизни сеголетков в растворе фенола 50 мг/л была значительно выше не только по сравнению с карасями 2+ (137,4 ч против 82,5), но и превосходила продолжительность жизни этих карасей в растворе с концентрацией фенола 25 мг/л. Еще большим оказалось различие устойчивости карасей обеих групп в растворе 100 мг/л (рис. 35). Средняя продолжительность жизни сеголетков была выше, чем у карасей в возрасте 2 + , в 35 раз (142 ч против 4,0).
Рис. 35. Динамика гибели карасей разных возрастных групп в растворе фенола 100 мг/л: 1 - возраст 2+ ; 2 - сеголетки.
Существенно отличные результаты были получены в опытах с концентрацией фенола 200 мг/л, которая за 10 дней вызвала гибель только 20% сеголетков, в то. время как караси в возрасте 2+ погибли в среднем за 43 ч
91

(рис. 36). Две последующие концентрации фенола 400 и 800 мг/л оказались по токсичности примерно равными для карасей обеих возрастных групп (рис. 37 и 38). При этих концентрациях испытуемые рыбы погибли в течение первых часов наблюдений.
-
Рис. 36. Динамика гибели карасей разных возрастных групп в растворе фенола 200 мг/л. Условные обозначения те же, что на рис. 35.
Рис. 37. Динамика гибели карасей двух возрастных групп в растворе фенола 400 мг/л. Условные обозначения те же, что и на рис. 35,
Рис. 38. Динамика гибели карасей двух возрастных групп в растворе фенола 800 мг/л. Условные обозначения те же , что и на рис. 35.
92
Анализ приведенных кривых летальности показывает, что сеголетки карася проявляют высокую устойчивость к фенолу при концентрациях до 50 мг/л. При более высоких концентрациях устойчивость значительно понижается и рыбы гибнут, хотя и живут дольше, чем караси в возрасте 2+. Особенно велико это различие в диапазоне концентраций фенола от 50 до 200 мг/л. Только при концентрациях 400-800 мг/л возрастные различия в устойчивости исчезают и время выживания рыб становится приблизительно равным. В опытах, полученных и на форели, и на карасях, видах, резко отличающихся по устойчивости к ядам промышленных сточных вод, сеголетки были более устойчивы, чем рыбы старших возрастных групп. Можно предположить, что обнаруженный факт имеет более широкое значение, и его следует учитывать в опытах с другими видами рыб. Совокупность имеющихся данных показывает, что устойчивость рыб к ядам неодинакова на разных этапах онтогенеза. Периоды высокой устойчивости (икра на стадии пульсирующего сердца, личинки на этапе С2 и сеголетки) чередуются с периодами низкой устойчивости (икра на стадии гаструляции, мальки в начале малькового периода, неполовозрелые особи) к ядам, Уровень токсико-резистентности определяется не только особенностями жизнедеятельности рыб на различных этапах онтогенеза, но и природой действующего токсического агента. В наших опытах устойчивость различных видов рыб к ядам органического ряда падала по мере онтогенетического развития и достигала минимума у половозрелых особей. Этот вывод подкрепляется Данными, полученными в ряде Других Лаборатории. Д. Бурц-Арле (Wurtz-Arlet, 1959) в опытах с развивающейся
. икрой и мальками форели обнаружил, что их устойчивость к двум детергентам (алкилсульфаты натрия) с возрастом падает. М. Катц и Хэдвик (Katz a. Chadwiek, 1961) установили, что развивающаяся икра и недавно выклюнувшиеся личинки колюшки оказались устойчивее, чем взрослые рыбы к токсическому действию эндрина. Е. А. Веселое, и др. (1965), изучая токсичность гексахлорана, пришли к выводу, что развивающаяся икра и личинки несколько устойчивее взрослых рыб (семга, плотва, уклея, окунь, ерш, щука). Сюда следует также отнести упоминавшиеся ранее данные Н. А. Мосевича и др. (1952), а также М. Ф. Вернидуб (1962).
Сформулированный нами вывод о снижении уровня устойчивости рыб к ядам органического ряда по мере созревания различных функциональных систем сочетания, деятельность которых определяет уровень - реактивности организма, следует считать обоснованным. Важная роль при этом принадлежит центральной нервной системе и ее синаптическим структурам, поскольку токсическое действие многих ядов органического ряда проявляется путем нарушения ее деятельности. Определенное значение при этом может иметь и состояние гипаталамо-гипофизарно-адреналовой системы, играющей важную роль в поддержании постоянства внутренней среды (гомеостаза). На ранних этапах онтогенеза постоянство внутренней среды и механизмы, ее регулирующие, не получают должного развития, вследствие чего организм способен переносить значительные отклонения oi оптимального уровня. В этом также можно усмотреть одну из причин пониженной реактивности организма к ядам в раннем онтогенезе.
Следует отметить, что при более тщательном изучении уровня устойчивости рыб к фенолу мы обнаружили, что в конце личиночного периода развития и начале малькового периода устойчивость резко падает, приближаясь к таковой половозрелых особей и даже несколько уступая ей.
Накопленные к настоящему времени данные по относительной устойчивости ранних этапов онтогенеза к ядам органического ряда не согласуются с мнением Н. С. Строганова и А. Т. Пажиткова (1941), что «наиболее уязвимыми стадиями онтогенеза рыбы для действия токсических веществ являются стадии личинки и малька» (с. 68). Это расхождение в оценке устойчивости отдельных этапов онтогенеза, вероятно, следует отнести за счет различной химической природы исследуемых веществ и различного механизма их воздействия на организм. Мы полагаем, что в зависимости от природы изучаемого токсического агента и вида испытуемых рыб устойчивость того или иного этапа онтогенеза может меняться. Это подтверждается имеющимися данными.
Поэтому необходимо более дифференцированно подходить к оценке роли возрастного фактора в определении устойчивости рыб к различным компонентам промышленных сточных вод.
Каждый раз, когда необходимо провести биологическое нормирование какой-либо новой группы токсикантов, объединенных химИ' ческим родством, необходимо специально изучать устойчивость рыб к этим компонентам промышленных сточных вод на различных этапах онтогенеза, ориентируясь при этом на наименее устойчивую стадию жизненного цикла рыб.
ХАРАКТЕР ЗАВИСИМОСТИ УСТОЙЧИВОСТИ РЫБ К ЯДАМ ОТ ВОЗРАСТА И МАССЫ
В процессе индивидуального развития наряду с возрастом меняются масса и размеры тела рыб. Поэтому при экспериментальных работах по изменчивости токсикорезистентности рыб в онтогенезе должны быть учтены и возраст и масса рыбы. Конечно, сопоставление устойчивости различных стадий эмбрионального развития, а также начальных периодов постэмбриональной жизни не встречает никаких осложнений. При постановке токсикологических экспериментов по определению устойчивости рыб к ядам на более поздних этапах онтогенеза естественно возникает вопрос, в какой мере устойчивость рыб зависит от возрастных особенностей исследуемой
93

группы рыб, а в какой от размерно-весовых?
Для решения этого вопроса можно сопоставлять устойчивость рыб разного возраста, но равной массы, либо определять токсикорезистентность у рыб разной массы, но одного возраста.
Втех немногих работах, где упоминается значение возраста в определении степени устойчивости рыб к ядам, совершенно не обращают внимания на размерно-весовые особенности испытуемых рыб. А в работах, где авторы (Cole, 1941; Neuhold a. Sigler, 1960; Sedsi, Sumio, Kadsunobu, 1964) отмечают зависимость устойчивости рыб к ядам от размеров, не указано, о каких возрастных группах идет речь. Поэтому трудно судить, в какой мере результаты этих опытов определяются весовыми, а в какой возрастными особенностями токсикорезистентности.
Мы предприняли специальное исследование для выявления уровня устойчивости рыб к фенолу у различных весовых групп одного возраста, чтобы тем самым определить, в какой мере токсикорези-стентность связана с массой, а в какой с возрастом испытуемых рыб. Опыты проводили на сеголетках и трехлетках радужной форели. Для определения роли массы в устойчивости рыб к фенолу использовали пять весовых групп (от 16 до 352 г) форели 2+ и три весовые группы (от 1,1 до 24,5 г) сеголетков. Показателем устойчивости каждой весовой группы служили процент и время гибели подопытных рыб в растворах фенола 10; 12,5 и 15 мг/л. В опытах с каждой концентрацией фенола использовалось 12 взрослых особей и 20 сеголетков. Испытание проводилось в 300-литровых аквариумах при температуре воды 12-14°С, рН 6,8-7,2, содержании кислорода 9,6-10,2 мг/л. Данные опытов подвеогали статистической обоаботке.
Впервой серии опытов, выполненной на форели трехлетнего возраста, при постепенном увеличении массы тела рыб время выживания в растворе фенола 10 мг/л сокращалось (табл. 45).
Обращает на себя внимание различная степень связи между массой и уровнем устойчивости рыб к фенолу, оцениваемой по времени выживания. При увеличении массы тела в два раза (с 16 до 34 г) время выживания практически не изменилось (367 мин и 360 мин). При увеличении массы в 4,5 раза (с 16 до 73 г) время выживания уменьшилось почти в шесть раз. Новое четырехкратное увеличение массы (с 73 до 302 г) не вызвало существенного изменения устойчивости к яду. Проверка достоверности различий полученных результатов в растворе 10 мг/л показала, что только при 4,5-кратном различии массы рыб имеет место статистически достоверное снижение устойчивости рыб. Интересно, что при одинаковом четырехкратном увеличении массы в двух весовых интервалах устойчивость изменялась неравноценно. В то время как в первом интервале весов различие в устойчивости достигало шестикратной величины и падало с увеличением массы рыб, во втором интервале весов устойчивость практически не изменялась.
Определение устойчивости форели различных весовых групп (от 16 до 352 г) в растворе фенола 12,5 мг/л показало (табл. 46), что с увеличением концентрации яда зависимость устойчивости рыб от массы становится менее выразительной. Только 22-кратное изменение массы испытуемых рыб привело к статистически достоверному снижению (в 2,6 раза) времени выживания форели массой 352 г в сравнении с 16 г. При 2, 3, 5, 7 и 10-кратном различии массы подопытных рыб имела место лишь статистически неоправданная тенденция уменьшения времени выживания более крупных рыб. В графической форме зависимость времени выживания от массы тела представлена на рис. 39. Полученная кривая при возрастании массы тела имеет тенденцию к прямой линии, параллельной абсциссе массы. Следовательно, в достаточно широком интервале устойчивость рыб остается практически неизменной и не зависит от массы тела.
94
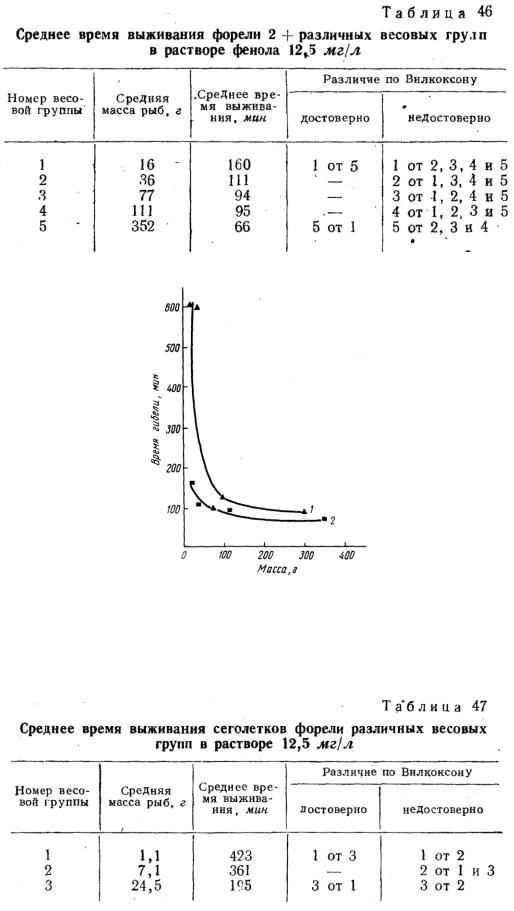
Рис. 39. Динамика гибели форели (2+) различных весовых групп
врастворах фенола 10 мг/л (1) и 12,5 мг/л (2).
Вопытах на сеголетках форели подобраны три группы с интервалом массы от 1,1 до 24,5 г. Результаты определения устойчивости различных весовых групп сеголетков форели даны в табл. 47.
При семикратном различии массы у сеголетков форели время выживания практически не изменилось и
95

находилось в пределах ошибки опыта. Только 24-кратное увеличение массы приводит к сокращению времени выживания в два раза. Построенный на основе этих данных график (рис. 40) выявляет отличие зависимости между массой тела сеголетков и временем выживания от аналогичной кривой, полученной для трехлетней форели. Эти факты указывают так же на зависимость характера связи между устойчивостью форели и ее массой от возраста.
Рис. 40. Динамика гибели сеголетков форели различных весовых групп в растворах фенола 12,5 мг/л (1) и 15 мг/л (2).
При определении устойчивости сеголетков форели к концентрации фенола 15 мг/л оказалось, что трехкратное различие массы (от 1,5 до 5,3 г) не оказывает существенного влияния на устойчивость рыб. Последующее трехкратное увеличение массы (от 5,3 до 15,4 г) приводит к отчетливому снижению времени выживания (табл. 48). Меньшие по массе рыбы гибнут в 2,1 раза медленнее, чем рыбы с большей массой. Примерно такая же степень различий по времени выживания между сеголетками, отличающимися по массе в 10
раз (1,5 и 15,4 г).
Сопоставляя результаты опытов, проведенных на сеголетках и форели 2 + , видно, что связь между массой тела и временем выживания сеголетков значительно слабее, чем у форели трехлетнего возраста. Необходимо отметить также наличие возрастных особенностей ускорения времени гибели рыб при многократном увеличении массы тела.
При одинаковой кратности изменения массы продолжительность жизни форели трехлетнего возраста изменилась в шесть раз, а сеголетков форели только в 2,1 раза.
Нельзя обойти вниманием и тот факт, что в опытах на трехлетней (2 +) форели связь между массой и временем выживания была более выразительна в растворе с минимальной летальной концентрацией фенола - 10 мг/л и значительно нивелировалась при увеличении концентрации до 12,5 мг/л. В противоположность этому опыты на сеголетках выявили более тесную связь между массой и устойчивостью рыб в растворе с более высокой концентрацией яда (в растворе 15 мг/л достоверная связь между массой и временем выжимания была получена при трехкратном различии массы, а в растворе 12,5 мг/л - только при 24,5-кратном). Таким образом, возрастной фактор оказывает заметное влияние на выраженность и характер связи между массой тела рыбы и временем выживания в токсическом растворе.
96
Мы не смогли провести опытов для выявления уровня устойчивости рыб разных возрастных групп, но одинаковой массы, однако можно сопоставить (для предварительной ориентировки) данные из предыдущей серии опытов. Если сопоставить время выживания 36-граммовой форели возраста 2+ и 24,5-граммовых сеголетков в растворе фенола 12,5 мг/л, то окажется, что устойчивость сеголетков почти в 1,8 раза выше (время выживания 195 мин), чем форели 2+ (время выживания 111 мин). Этот факт подкрепляет ранее сформулированный вывод о зависимости устойчивости от возраста испытуемых рыб и более высокий ее уровень к ядам органического ряда у младших возрастных групп по сравнению со старшими. Следует отметить, что выявленные различия токсикорезистентности сеголетков и трехлетней форели примерно равных весовых групп менее выражены (время выживания 195 и 111 мин, соответственно) по сравнению с токсикорезистентностью этих же возрастных, но отличных весовых групп (время выживания 336 и 95 мин, соответственно).
Суммируя данные наших исследований, направленных на выявление зависимости между массой тела и временем выживания форели двух возрастных групп, можно сделать вывод, что эта зависимость имеет весьма сложный характер и определяется многими переменными, среди которых следует указать концентрацию яда, возраст рыб и разницу масс испытуемых рыб. Зависимость между массой тела и устойчивостью более выражена у форели трехлетнего возраста в сравнении с сеголетками. С увеличением концентрации яда различие в устойчивости рыб разных весовых групп одного возраста падает. Значение фактора массы становится реальным и статистически оправданным только при различиях в массе более чем в три раза, а у сеголетков только по достижении 19-22-кратных различий. Помимо указанных факторов зависимость устойчивости от массы определяется также и видовыми особенностями испытуемых рыб (Burdick, Dean, Howard, 1964). Изучая зависимость токсического действия аквалина (гербицид) от размера рыбы на молоди гольцов и ушастого окуня, авторы обнаружили, что устойчивость ушастых окуней повышается с увеличением размера, а у гольцов такой зависимости выявить не удалось.
Наши данные по зависимости устойчивости радужной форели к фенолу от массы тела нельзя перенести на другие виды рыб и даже другие возрастные группы этого вида.
С. Гуднайт (Goodnight, 1942) считает, что масса и размеры играют незначительную роль в определении устойчивости рыб к яду, а японские исследователи А. Седзи, Н. Сумио и К. Кадзунобу (1964) установили линейную зависимость между логарифмом массы тела вьюна и логарифмом времени выживания, т. е. пришли к прямо противоположным выводам. Это можно объяснять тем, что авторы не учитывали другие факторы, оказывающие влияние на эту связь.
Необходимо уточнить, что рассматривая вопрос о роли возрастного фактора в определении устойчивости рыб к ядам и его связи с размером тела и массой, мы ведем речь лишь о возрастных группах, начиная с сформировавшихся неполовозрелых особей. Что касается ранних этапов онтогенеза, то этот вопрос мы уже рассмотрели выше.
Можно считать, что возрастной фактор имеет существенное значение и при сопоставлении устойчивости взрослых рыб (неполовозрелых и половозрелых), однако необходимы дальнейшие исследования. Работа в этом плане важна в методическом и практическом отношении. Если окажется, что фактор массы играет ведущую роль в определении чувствительности и устойчивости рыб к ядам по сравнению с возрастным, то испытание токсичности следует проводить на рыбах, руководствуясь размерно-весовыми показателями. Роль и удельный вес каждого из двух рассматриваемых факторов необходимо учитывать при межвидовых сопоставлениях токсикорезистентности рыб, а также при выборе тест-объекта. Наши эксперименты склоняют к выводу, что ведущими являются возрастные особенности токсикорезистентности рыб. Это подтверждается данными Р. Ейслера и Д. Дейла (Eislera. Deuel, 1964), а также Б. Рейфа (Reiff, 1964). В первой из этих работ, выполненной на трех видах эстуарных рыб: кефаль (Mugil cephalus), фундулус (Fundulus heteroCLitus) и мендия (Mendia menidia),
авторы показали, что время выживания в растворах различных мыл определяется возрастными особенностями и не зависит от размера рыб. Б. Рейф считает, что концентрации ядов, вызывающие гибель 50% испытуемых рыб
.(CL50), У больших и маленьких рыб весьма сходны и не определяются размерами рыб. Следует иметь в виду, что работы в этом плане только начинаются и в дальнейшем могут быть получены данные, которые внесут важные коррективы к высказанной точке зрения. Априорно можно предполагать, что в зависимости от химической природы вещества и механизма его действия на рыб, при прочих равных условиях, размер рыб может иметь существенное значение (особенно в опытах с группой контактных ядов) в определении времени выживания и уровня устойчивости. Но и возрастной фактор будет иметь определенное значение, ибо поглощение токсического агента на единицу массы не всегда соответствует размеру рыб, а чувствительность и устойчивость их значительно меняется с возрастом.
ВИДОВЫЕ ОСОБЕННОСТИ ЧУВСТВИТЕЛЬНОСТИ И УСТОЙЧИВОСТИ РЫБ К ЯДАМ
Определение пороговой токсичности вредных веществ и предельно допустимых концентраций немыслимо без учета видовых особенностей рыб, обитающих в конкретном водоеме. Между тем испытание токсичности
97
различных компонентов сточных вод осуществляется, как правило, на случайных, оказавшихся доступными видах рыб, без учета уровня их чувствительности и устойчивости к ядам. В результате предельно допустимые концентрации, полученные, скажем, в опытах с такими устойчивыми видами рыб, как карась или золотая рыбка, оказываются чрезвычайно завышенными для других рыб. В методическом отношении принципиально важен вопрос: какие именно виды следует брать в качестве тест-объекта для определения пороговых и предельно допустимых концентраций токсических веществ.
Изучение видовых особенностей чувствительности и устойчивости рыб к ядам имеет важное значение и для понимания адаптационных возможностей разных видов рыб, а также для направленного изменения видового состава рыбохозяйственного водоема под влиянием ядов промышленных сточных вод. Исследования видовых особенностей устойчивости рыб к ядам имеют первостепенное значение и для разработки теоретических основ химического метода преобразования ихтиофауны того или иного водоема.
Литература рассматриваемого вопроса весьма обширна, но фрагментарна и различна по своей ценности. Целенаправленное изучение видовых особенностей устойчивости рыб с использованием широкого ассортимента видов проводилось крайне редко (Строганов и Пажит-ков, 1941; Таусон, 1947; Hart, 1947).
В ранних работах, решай вопрос о степени токсичности различных компонентов промышленных сточных вод, использовали 1-3 вида, а критерием токсичности служила величина смертельной концентрации (Hein, 1908; Steimann, Surbeck, 1920; Holzinger, 1927, Belding, 1927; Демьяненко, 1931 и др.). Когда перед исследователями возникла необходимость определять видовые особенности устойчивости рыб к ядам, они продолжали использовать показатели гибели животных, но учитывали не время гибели, при одинаковой концентрации, как это принято в патофизиологическом эксперименте, а различие смертельных концентраций (Ebeling, 1939; Cole,
1941; Чистяков, 1941; Hart, 1946; Thrams, 1950; Веселов, 1957; Lammering, Bur-bank, 1960 и др.). При оценке степени устойчивости рыб к яду на основе величины смертельной концентрации яда нередко видовые особенности токсикорезистентности проявлялись в стертой форме или не выявлялись совсем (Ebeling, 1939; Sierp, 1940; Cole, 1941). В этих работах результаты испытания токсичности фенола давались в неоправданно обобщенном виде и создавалось впечатление, что устойчивость рыб к ядам в малой степени определяется видовой принадлежностью. Например, Г. Эбелинг (1939) считает, что для угря, линя и карпа смертельные концентрации лежат в области 10-15 мг/л. По мнению Ф. Сиерпа (1940), летальные концентрации фенола для столь разных видов рыб, как форель, щука и карп, относящихся к различным семействам, почти совпадают и находятся в интервале от 5 до 10 мг/л. Более поздние работы не принесли необходимое ясности в этот вопрос. К. Вурман и X. Вокер (Wuhrmann, V/oker, 1950) считают, что пороговые токсические концентрации фенола для форели и голавля равны 9-10 мг/л, для окуня - 12 мг/л. В сообщении X. Бандта (Bandt, 1958) указывается, что пороговая повреждающая концентрация гидрохинона (в течение 3-4 дней) для карпа, окуня и колюшки оказалась равной 0,2 мг/л. В обширном исследовании Н. С. Строганова и А. Т. Пажиткова (1941), также получены данные, показывающие, что пороговые концентрации меди и аммиака для многих видов довольно близки или полностью совпадают.
Наряду с этим имеются данные (Winterstein, 1908; Reuss, 1909, 1910; Ebeling a. Schrader, 1929; Belding, 1929; Jones, 1938; Hurt, 1945; Wuhrmann, Woker, 1948), свидетельствующие о важной роли видовых особенностей испытуемых рыб в определении уровня устойчивости ко многим токсическим компонентам промышленных сточных вод: хлору, ядам тяжелых металлов, аммиаку, углекислоте и др.
Исследователи, которые судят об уровне устойчивости или чувствительности рыб разных видов на основе сопоставления величин смертельных концентраций яда, по существу не дифференцируют совершенно различные по своему физиологическому содержанию понятия «чувствительность» и «устойчивость». Они используют их несоответственно той степени токсического воздействия, исход которой регистрировался (Shelfort, 1917; Wells, 1918; Cole, 1941; Чистяков, 1941; Таусон, 1947 и др.). В этих работах не только отсутствует попытка анализа и конкретизации этих двух понятий, но и делаются выводы о «большей устойчивости» или, что то же, «меньшей чувствительности» того или иного вида рыб исключительно на основе выявляемых различий между летальными концентрациями. Считают при этом, что высокой чувствительности соответствует низкая устойчивость и, наоборот. М. Велс (1918), например, пишет о том, что «рыбы более чувствительны или менее резистентны» к различным факторам внешней среды, а А. О. Таусон (1947) повторяет ту же ошибку и считает, что «...представители семейства окуневых (окунь, ерш, судак) являются наименее стойкими, наиболее чувствительными ко всем исследованным органическим соединениям» (с. 36). Такой подход полностью игнорирует физиологические особенности организма, существование компенсаторных и приспособительных механизмов, направленных на поддержание гомеостаза в переменных условиях среды. Предполагается, что токсический процесс у рыб развивается только по схеме: первичная токсикологическая реакция - конечный эффект. В специальной литературе по фармакологии, иммунологии и общей токсикологии разграничение этих понятий так же встречается весьма редко. Только благодаря усилиям Н. Н. Сиротинина (1951, 1952) было привлечено внимание к необходимости разграничивать выносливость и чувствительность организма при характеристике его реактивности к тому или иному веществу. Причину сложившегося положения следует усмотреть в том, что физиологи недостаточно уделяют внимания двум важнейшим физиологическим феноменам; устойчивости и чувствительности организма к разнообразным альтерирующим воздействиям.
98
При рассмотрении видовых особенностей чувствительности и устойчивости рыб к ядам необходимо обратить внимание и на содержание, которое вкладывают различные авторы в понятие «видовые особенности». Абсолютное большинство исследователей для выявления видовых особенностей используют рыб, относящихся подчас к различным родам, семействам и даже отрядам. Такие «видовые» различия могут быть обусловлены особенностями устойчивости, свойственной данному отряду или семейству. В строгом смысле слова видовые различия токсикорезистентности могут быть обнаружены только в опытах на систематически близких видах рыб. Работы на систематически близких видах необходимы и для целенаправленного поиска таксономически важных функциональных критериев вида, которые, как правило, менее устойчивы, чем морфологические, и потому трудно поддаются учету. Существует точка зрения (Fry, 1957; Ушаков, 1959), что в качестве функциональных критериев вида целесообразно использовать показатели, характеризующие пределы устойчивости организма или его отдельных систем к различным факторам внешней среды, причем в дозировках, превышающих нормальные. Ф. Фрай (1957), например, в качестве видового критерия использовал температуру гибели и пришел к выводу, что этот показатель, характеризующийся большой стабильностью, может быть использован при описании функциональных особенностей вида. Б. П. Ушаков и его сотрудники (1965) считают возможным использовать теплоустойчивость клеток и белков в качестве цитофизиологического критерия вида.
Мы предприняли сравнительное изучение чувствительности и устойчивости 21 вида рыб к фенолу, с тем чтобы в сходных методических условиях получить данные относительно особенностей этих двух показателей. В опытах были использованы представители 14 родов, объединенных в семь семейств.
Осетровые: осетр (Acipenser guldenstadtl Brandt), севрюга (Acipenser stellatus Pall) и стерлядь (Acipenser ruthenus).
Лососевые: ручьевая форель (Salmo trutta L.) и радужная форель (Salmo irrldeus Gibb).
Щуковые: щука (Esox lucius L).
Карповые: лещ (Abгamis brama L.), синец (Abramis ballerus, L.), белоглазка (Abramis sapa Pall), красноперка (Scardinius erythroph-talmus), чехонь (Pelecus cultratus), плотва (Rutilus rutilus), вобла (Rutilus rutilus caspicus), тарань (Ratilus rutius heckeli), жерех (Aspius aspius L.) и карась (Carassius carassius).
Сомовые: сом (Silurus glanis). Тресковые: налим (Lota lota L.).
Окуневые: судак (Lucioperca lucioperca), окунь (Perca fluviati-lis) и ерш (Acerina cernua).
Таким образом, в опытах использован достаточно широкий набор видов рыб, резко отличающихся по систематическому положению. Вместе с тем сопоставление чувствительности и устойчивости к ядам трех видов рода Acipenser (осетр, севрюга и стерлядь), двух видов рода Salmo (ручьевая и радужная форель) и трех видов рода Abramis (лещ, синец, белоглазка) позволяло ответить на вопрос об уровне токсикорезистентности у систематически близких видов рыб.
Часть опытов была выполнена совместно с Б. А. Флеровыми зимне-весенний период 1962/63 г. на рыбах, выловленных в Рыбинском водохранилище (лещ, синец, карась, плотва, ерш, окунь, налим, щука). Другая часть была поставлена в июле - августе в дельте Волги (вобла, жерех, тарань, белоглазка, лещ, синец, чехонь, судак, сом, красноперка, осетр, стерлядь, севрюга).
Для сопоставимости данных эксперименты ставились при температуре 6-8° С, содержании кислорода 8-10 мг/л, рН 6,8-7,2. Опыты ставились в 150-литровых плексигласовых аквариумах в течение 24 ч. Летом температура раствора достигала 19-23° С, а содержание кислорода находилось в пределах 5,5-6,5 мг/л. Для максимальной стабилизации условий проведения опытов использовали рыб, выловленных в одном водоеме, примерно одинакового возраста и биологического состояния. Сравнивались результаты, полученные в один сезон и при одинаковых температурах. В качестве показателя уровня токсикорезистентности рыб использовали время гибели животных в растворах фенола различных концентраций: 5, 10, 15, 20, 25, 30 и 40 мг/л. Это позволяло выявить уровень устойчивости даже в том случае, если минимальные смертельные концентрации были равными.
Определение чувствительности испытуемых рыб к фенолу проводилось на основе сопоставления величин латентного периода первой фазы фенольной интоксикации - бурной двигательной активности. Мы исходили при этом из представления, что понятие «чувствительность» по физиологическому содержанию близко или совпадает с понятием «возбудимость», уровень которой определяет порог раздражения. Применительно к действию токсических веществ порог раздражения или уровень чувствительности можно выразить либо минимальной концентрацией, вызывающей специфическую для данного вещества реакцию со стороны одной из функциональных систем организма, либо временем развития этой реакции в условиях действия определенной концентрации токсического агента.
Результаты измерения латентного периода проявления первой фазы фенольной интоксикации - двигательной активности - представлены в табл. 49. Оказалось, что различные виды рыб весьма существенно отличаются по чувствительности к ядам. Латентный период двигательной реакции в растворе фенола 10 мг/л варьировал в зависимости от вида испытуемых рыб от 30-35 мин (налим и лещ) до 1,5-2 мин (радужная и ручьевая форель), т.е. приблизительно в 15 раз. Равномерное увеличение концентрации фенола на 5 мг/л (с 10 до 25 мг/л) приводило к резкому сокращению латентного периода двигательной активности, особенно в опытах с теми видами рыб, у которых был отмечен высокий исходный латентный период. При 2,5-кратном увеличении концентрации фенола латентный период сократился у налима почти в шесть раз, у леща в 10 раз, у радужной
99

форели в девять раз, у ручьевой форели в четыре раза, а у окуня только в два раза. Причина такой неравномерности не совсем ясна и может быть обусловлена как различной чувствительностью нервной системы к величине градиента концентрации, так и функциональными особенностями локомоторного аппарата. Вероятно, уровень чувствительности того или иного вида рыб к различным ядам промышленных сточных вод будет меняться в зависимости от того, какая функциональная система избрана в качестве показателя для суждения о степени чувствительности. В каждом отдельном случае, в зависимости от природы вещества и механизма его токсического действия, придется выбирать те функциональные системы (двигательная, дыхательная, сердечнососудистая и т.д.), в деятельности которых в первую очередь отмечаются Сдвиги, и использовать их в качестве показателей чувствительности. Принимая во внимание уровень чувствительности, изученные виды можно условно разбить на две группы: высокочувствительные - радужная и ручьевая форель, плотва и ерш и низкочувствительные виды - лещ, налим, синец и карась. Щука и окунь по своей чувствительности занимают промежуточное положение.
Опыты, проведенные летом, показали, что наиболее чувствительны осетр и стерлядь, наименее - жерех и севрюга. Необходимо отметить, что выраженность двигательной активности у рыб летом при высоких температурах стерта (по сравнению с зимой). При сопоставлении латентного периода реакции рыб на токсическое вещество у видов, близких в систематическом отношении, определенной связи не обнаружено. Латентный период двигательной реакции у осетра и стерляди оказался близким 3,5 и 4 мин (при 20 мг/л фенола), а у севрюги 10 мин (более чем в два раза). Лещ и синец имеют сходный латентный период реакции на фе-нольное раздражение - 3-4 и 3 мин (при 20 мг/л). Ручьевая и радужная форель характеризуются сходным латентным периодом реакции в растворе фенола 10 мг/л- 2 и 1,5 мин, но заметно раз-I; личаются (в три раза) при использовании более концентрированных растворов: 15 и 20 мг/л. Наконец, в растворе фенола 25 мг/л Ц различие между латентными периодами реакции этих двух видов вновь сходны: 6,5 и 0,3 мин. Таким образом, уровень чувствительности рыб к фенолу широко варьирует от вида к виду.
Обзор табличных данных, характеризующих устойчивость рыб (табл. 50), показывает, как зажно в качестве показателя устойчивости использовать время гибели рыб, поскольку минимальные летальные концентрации для систематически удаленных видов могут оказаться одинаковыми. Так, например, минимальная летальная концентрация фенола для щуки и налима оказалась одинаковой и равна 10 мг/л. Вместе с тем время гибели щуки и налима при этой концентрации различается в 3,5 раза, а по мере увеличения концентрации фенола с 10 до 25 мг/л среднее время гибели обоих видов рыб различается уже в 10 раз. Следовательно, особенности реакции этих видов рыб более выражены при высоких концентрациях яда. Скорость развития интоксикации у
100