

АКАДЕМИЯ
НАУК СССР
ОБЪЕДИНЕННЫЙ
НАУЧНЫЙ СОВЕТ «ФИЗИОЛОГИЯ ЧЕЛОВЕКА И
ЖИВОТНЫХ»
ПО ФИЗИОЛОГИИ
Физиология
СЕНСОРНЫХ СИСТЕМ
ЧАСТЬ
ВТОРАЯ
в
ИЗДАТЕЛЬСТВО
«НАУКА» ЛЕНИНГРАДСКОЕ
ОТДЕЛЕНИЕ
ЛЕНИНГРАД
• 19 7 2
УДК
612.85/.88
Физиология
сенсорных систем. Ч. 2. В
серии «Руководство по физиоло гии».
1972. Изд-во «Наука», Ленингр. отд., Л.
1—703.
В
томе рассматривается широкий круг
вопросов, касающихся деятельности
органов чувств: органа слуха, равновесия
(вестибулярный аппарат), обоняния,
вкуса, осязания, болевой и температурной
чувствительности. В каждом из этих
разделов излагаются сведения о физических
и химических свойствах внешнего
сигнала, с которым связана деятельность
данного органа чувств, о структуре
периферических и центральных отделов,
данные о психофизиологических
характеристиках деятельности органов
чувств, биофизических и нейрофизиологических
основах их дея- тельности по обнаружению
и различению свойств внешнего сигнала.
Освещаются также физиологические
основы восприятия наиболее важных для
человека звуковых сигналов, какими
являются звуки речи. Илл.—241, табл. —
13, библ.—2271 назв.
Редакционная
коллегия руководства:
акад.
П.
К. Анохин (зам.
отв. редактора),
чл.-корр. АН СССР Э.
А. Асратян,
акад.
П.
С. Бериташвили, чл.-корр.
АН СССР
Л. Г. Воронин,
акад.
Е.
М. Крепе, К. А. Ланге (отв.
секретарь),
акад. М.
Н. Ливанов,
акад.
В.
В.
Парии
(зам.
отв, редактора),
чл.-корр. АН СССР
А.
М. Уголев, акад.
В.
Н. Черниговский (отв.
редактор).
В.
В. Парии
Редакционная
коллегия тома:
д.
м. н. Я. А. Альтман, к.
м. н. А.
В. Бару, к.
м. н. И.
А. Вартанян (отв.
секретарь),
чл.-корр. АН СССР Г.
В. Гершуни (отв.
редактор), д.
б. н. О.
Б. Ильинский, к.
б. н. В.
А. Кисляков, д.
б. н. Л.
А. Чистович.
5-3-1;
5-3-11
928-72
ПРЕДИСЛОВИЕ
Первая,
уже вышедшая из печати часть «Физиологии
сенсорных систем» посвящена зрению.1
В настоящей, второй части рассматриваются
общая физиология рецепторов, физиология
кожной чувствительности, вестибулярного
аппарата (органа равновесия), слуха,
обоняния и вкуса. Излагаются также
вопросы восприятия звуков речи и
речеобразования, включение которых в
книгу по физиологии сенсорных систем
не является случайным. При современном
уровне развития науки изучение слуха
и речи у человека неотделимы друг от
друга в своих общих основах, точно так
же как изучение звуковосприятия и
звукоизлучения в животном мире в целом.
В
Руководстве по физиологии, в томе по
физиологии сенсорных систем в его
настоящем объеме может быть представлен
материал, ограниченный строго определенным
кругом вопросов. Этот круг вопросов
охватывает физиологию тех основных
сенсорных систем, которые обозначались
как органы внешних чувств, а в более
позднее время в связи с появлением
новых понятий такими терминами, как
«экстероцептивные системы», «внешние
анализаторы». В настоящей книге не
рассматриваются вопросы физиологии
собственных чувствительных систем
организма, а именно двигательного
аппарата (мышц и суставов) и внутренних
органов, обозначаемых такими терминами,
как «проприоцептивные» и «интероцептивные
аппараты», а также «внутренние
анализаторы». Эти вопросы излагаются
в других томах Руководства по
физиологии.
В
настоящий том включена физиология
вестибулярного прибора; этот прибор
в равной мере может быть отнесен к
органам и экстероцепции, и проприоцепции.
Как
указывалось в предисловии к первой
части, в физиологии органов чувств
уже со второй половины прошлого века
выделились различные направления
исследования. Эти направления схематически
разделялись на следующие: 1) биофизическое
(и биохимическое), рассматривающее
процессы, происходящие в периферических
аппаратах органов чувств; 2) собственно
физиологическое, посвященное
процессам в чувствительных нервах и
связанных с ними центральных отделах
данной сенсорной систе
1 Физиология
сенсорных систем. Ч. I. Физиология
зрения-(В серии: Руководство по
физиологии). «Наука», Л., 1971.
1*
3
мы;
3) психофизиологическое, имеющее
предметом изучения восприятие
объектов внешнего мира в целом. Это
последнее направление в своем развитии
наиболее тесно связано с общими вопросами
изучения высших функций центральной
нервной системы, которые обеспечиваются
интегративной деятельностью всего
мозга, а не только ограниченных его
отделов (структурно образующих так
называемые специфические центральные
сенсорные аппараты). Именно эти отделы
рассматриваются при изолированном
изучении отдельных сенсорных систем.
Во
второй части в основном излагается
круг вопросов, который ограничивается
первыми двумя направлениями изучения
сенсорной функции. Третье направление
— изучение интегративной деятельности
мозга, обеспечивающей процессы восприятия
внешних объектов в целом, — не могло
быть включено в настоящем его объеме.
Некоторые
из вопросов, связанных с общими
механизмами высшей нервной деятельности,
относящиеся^ восприятию признаков
внешних объектов, рассматриваются
очень кратко в томе Руководства,
посвященном физиологии высшей нервной
деятельности (см. главы 6 и особенно
7).2
Несомненно,
круг вопросов, касающихся изучения
общих (интегративных) механизмов
деятельности мозга, лежащих в основе
восприятия объектов внешнего мира,
весьма широк. Его изложение требует
рассмотрения не изолированной функции
отдельных сенсорных систем, а
закономерностей совместного
функционирования различных сенсорных
аппаратов соответственно множественности
действующих факторов внешней и
внутренней среды при осуществлении
организмом определенных задач.
Следует
указать, что учитель многих авторов
настоящей книги Л. А. Орбели, так много
сделавший для развития изучения органов
чувств в нашей стране, многократно"подчеркивал
в своих работах важность задачи
специального рассмотрения взаимоотношения
сенсорных (афферентных) систем для
понимания общих закономерностей их
деятельности.3
Рассмотрение
накопившихся к настоящему времени
данных о механизмах деятельности мозга,
при помощи которых может строиться
восприятие (представление) объектов
внешнего мира на основе информации,
поступающей от множества сенсорных
систем активно действующего организма,
представляет значительные трудности
для систематического изложения,
соответствующего современному уровню
развития разных направлений изучения
нервной системы и органов чувств. Можно
надеяться, что в дальнейшем все же
удастся рассмотреть в отдельном томе
Руководства и этот круг вопросов.
2 Физиология
высшей нервной деятельности. 4.1. (В
серии: Руководство до физиологии).
«Наука», М., 1970.
3 Фдздодогидеский
журнал СССР им. Сеченова, 17, 1105, 1934.
Р
Л 3Д Е Л I
ОБЩАЯ
ФИЗИОЛОГИЯ РЕЦЕПТОРОВ. КОЖНАЯ
ЧУВСТВИТЕЛЬНОСТЬ. ВЕСТИБУЛЯРНАЯ СИСТЕМА
ОБЩАЯ
ФИЗИОЛОГИЯ РЕЦЕПТОРОВ
ВИДЫ
РЕЦЕПТОРНЫХ СТРУКТУР
Живые
организмы не могут существовать, не
получая информации о состоянии внешней
и внутренней сред и об изменениях,
происходящих в них. Чем быстрее и точнее
организм получает такую информацию,
тем выше при прочих равных условиях
его шансы в борьбе за существование.
Поэтому в ходе эволюции животные
организмы выработали различные рецепторы
— специальные образования,
предназначенные для трансформации
энергии раздражающего стимула в
специфическую активность нервной
системы, в сигналы, несущие нервным
центрам информацию о действующем
агенте.
Существует
огромное разнообразие рецепторов,
позволяющих животным очень точно
воспринимать стимулы самых разных
модальностей: механические, химические,
температурные, световые, электрические.
Одни из сенсорных структур — мономодальные
— оказались приспособленными для
восприятия лишь одного вида раздражения
(светового, механического, температурного
и т. д.), другие же — полимодальные —
нескольких (механического и температурного;
механического и химического и т. п.).
Часть рецепторов (дистантные) служат
для получения информации на некотором
расстоянии от источника раздражения,
остальные же (контактные) — лишь при
непосредственном соприкосновении с
ним. Основная масса рецепторных приборов,
особенно высокоспециализированных,
обращена во внешнюю среду (экстеро-
цепторы). Однако не менее важную роль
играют рецепторы, сигнализирующие о
раздражителях внутренней среды и
двигательной системы организма (интеро-
и проприоцепторы).
Как
показали исследования последних
десятилетий, все рецепторные приборы
независимо от их модальности, назначения,
расположения и т. д. по своей функциональной
организации и морфологическому строению
могут быть разделены всего лишь
5Глава 1
Рис.
1. Схематическое изображение^ответов
первичного (Л) и вторичного
(Б)
рецепторов на раздражение.
А
—
тельце Пачини (ориг.); Б
—
волосковый механорецептор боковой
линии (по: Flock, 1965). На А:
1 —
вспомогательные структуры (капсула)
рецептора; 2
—
немиелинизиро- ванная часть нервного
окончания; з
—
миелинизированное нервное волокно; РП
— рецепторный
потенциал; ПД
—
потенциал действия. На Б:
РК —
рецептирующая клетка и ее афферентное
(АН)
и эфферентное (ЭН)
нервные волокна; РП
—
рецепторный потенциал; С
—
синапс; стрелками
сверху
показано направление раздражения.

на
две группы: на первичные (первичночувствующие)
и вторичные (вторичночувствующие). К
первичным относятся такие рецепторные
аппараты, у которых воспринимающий
внешнее воздействие субстрат заложен
в самом сенсорном нейроне, который
поэтому непосредственно (первично)
возбуждается раздражителем (рис. 1, А).
К вторичным же рецепторам относятся
те, у которых между действующим агентом
и сенсорным нейроном располагаются
дополнительные, специализированные
(рецептирующие) клетки (рис, 1, Б),
В этом случае процесс преобразования
энергии внешнего раздражения
протекает уже не в сенсорном нейроне,
а в ре- цептирующих клетках. Последние
в свою очередь активируют сенсорный
нейрон, который, следовательно,
возбуждается уже не первично внешним
стимулом, а опосредованно (вторично),
благодаря воздействию рецептирующих
клеток. К первичным рецепторам у
высокоразвитых организмов относятся,
например, обонятельные и тактильные
рецепторы, мышечные веретена, а к
вторичным — рецепторы органов слуха,
зрения, вкуса, боковой линии,
вестибулярного аппарата.
ПОСЛЕДОВАТЕЛЬНОСТЬ
ПРОЦЕССОВ, ПРОТЕКАЮЩИХ В РЕЦЕПТОРАХ
Между
раздражителем и рецептирующим субстратом
практически у всех сенсорных аппаратов
располагаются специализированные
вспомогательные структуры, а часто,
кроме того, и обычные элементы ткани,
в которой локализуются рецепторы. Все
они существенно изменяют характеристики
раздражающих воздействий на их пути к
рецептирующему субстрату и поэтому
играют важную роль в деятельности
рецепторов.
Пройдя
через вспомогательные структуры,
адекватный раздражитель взаимодействует
с рецептирующим субстратом, определяющим
модальность рецептора, и вызывает
развитие в нем специфических для данного
типа рецепторов трансформационных
процессов, следствием которых является
изменение проницаемости поверхностной
мембраны клетки. В результате возникает
ионный ток, о котором обычно судят по
развитию локальной электрической
реакции рецептора, так называемого
рецепторного потенциала (РП). Изменение
проницаемости клеточной мембраны под
влиянием специфического раздражителя
происходит лишь в области собственно
воспринимающей структуры рецептора,
которая, по мнению многих исследователей,
является электрически невозбудимой
или мало возбудимой (Grundfest, 1961а, Ь, 1965;
Loewenstein, 1961, 1965; Bennett, 1961, 1967; Ильинский,
1967, и др.).
При
возбуждении первичного рецептора
ионный ток, лежащий в основе РП, оказывает
деполяризующее действие на электрически
7

возбудимые,
регенеративные элементы рецептора.1
В случае, если деполяризация достигает
некоторого критического уровня, в зоне,
наиболее приспособленной к возникновению
спайков, появляются нервные импульсы,
распространяющиеся к центру. Таким
образом, в первичном рецепторе
специфические трансформационные
процессы и возникновение импульсной
активности происходят в одной и той же
сенсорной клетке.
Во
вторичном рецепторе трансформационные
явления протекают в рецептирующей
клетке, где и возникает РП. Под влиянием
РП из рецептирующей клетки обычно
выделяется медиатор, который действует
на нервное окончание сенсорного нейрона
и вызывает появление в нем локального
электрического ответа — постсинаптического
потенциала. Если последний оказывает
деполяризующее действие на отходящее
нервное волокно, в нем возникает
импульсная активность. Следовательно,
у вторичных рецепторов место протекания
специфических трансформационных
явлений и область генерации нервных
импульсов находятся в разных клетках.
В отличие от первичного во вторичном
рецепторе локальная деполяризация
возникает дважды: как в рецептирующей
клетке, так и в сенсорном нейроне. Чтобы
избежать терминологической путаницы,
было предложено называть градуальный
электрический ответ рецептирующей
клетки «рецепторным потенциалом»,
а локальную деполяризацию сенсорного
нейрона — «генераторным потенциалом»
(ГП), имея в виду, что он «генерирует» в
отходящем от рецептора нервном волокне
бегущие волны возбуждения. В первичных
рецепторах эти понятия совпадают
(Davis, 1961).
В
последние годы было показано, что при
действии адекватных раздражителей
в рецепторах может возникать не только
возбуждение, но и торможение. Первому
соответствует появление локальных
деполяризационных, а второму —
гиперполяризационных реакций.
Особенно легко тормозные процессы
обнаруживаются У рецепторов, обладающих
фоновой активностью (рис. 1, Б).
Гиперполяризация
рецепторов может быть не только
вторичной, т. е. следующей за деполяризацией,
но и первичной (Ильинский. 1965, 1966а; Flock,
1965; Nishi, Sato, 1968, и др.).
В
своем первоначальном значении РП
означал всегда локальный
деполяризационный ответ рецептора.
Если понятие «генераторный потенциал»
продолжает и сейчас являться синонимом
деполяризационной реакции, то термин
«рецепторный потен
1
Регенеративными структурами называют
те элементы рецептора, которые
ответственны за возникновение
регенеративного процесса генерации,
т. е. самоускоряющегося процесса активной
деполяризации, ведущего к возникновению
ответа «все или ничего». Нерегенеративными
же структурами называют те элементы
рецептора, которые ответственны за
появление нерегенеративного процесса
генерации. Результатом последнего
является возникновение РП. -
8
циал»
требует теперь уточнения. Было бы
наиболее разумно называть по аналогии
с возбуждающими и тормозными постси-
нангическими потенциалами (ВПСП и ТПСП)
локальные деполяризационные ответы
рецепторов — возбуждающими РП, а
гиперноля ризационные реакции —
тормозными РП (Ильинский, 1965).
В
цепи явлений, предшествующих возникновению
импульсной активности, особое место
занимают трансформационные процессы.
вспомогательные
структуры
внешняя
энергия
Защищает
Проводит
Концентрирует
Анализирует
Тормозит
Повышает
чувствительность
Чувствительный
орган
Сенсорный
нейрон
_
„ Начальный сегмент
Дендрит q0MQ
рксон
Терминаль
Постсцнаптический
(генераторный) ,•
потенциал
Нервный импульс
Процесс\
генера-
j
ции
I *
Рецептирующая
клетка
Рецепторный
потенциал
Процесс
Процесс
транс- генера-
(рормации
ции
Преобразует
Усиливает
Секретирует
Синапс
Медиатор
Передает
Гоадуальный
электрический и химический ответ
Нерв
Центр
Синапс
Медиатор
Процесс
транс-
формации
Преобразует
| |
Усиливает :
■
Генерирует I
|
Интегрирует
i
: Проводит
I
I Секретирует
Градуальный i
„ л
„ I I
электрический
I Ответ
все или ничего
> I
ответ
; и
a |
<
Кодирует Декодирует
\^Периферическое
взаимодействие
Передает
-ит.д.
Эфферентный,
нейрон
Рис.
2. Общая схема процессов, происходящих
в сенсорном органе
(по: Davis, 1961).
Их
следует четко разграничивать от
процессов генерации, связанных с
перемещением ионов через поверхностную
мембрану сенсорной клетки. Процессы
трансформации у подавляющего большинства
рецепторных структур изучены крайне
недостаточно. Только в отдельных
случаях, например у фоторецепторов,
исследователи знают о них несколько
больше.
Заканчивая
перечисление явлений, протекающих в
рецепторных аппаратах, следует еще
упомянуть о том, что нервная система,
получая от рецепторов сигналы о
действующем раздражителе, часто в
свою очередь оказывает на них регулирующее
(эфферентное) влияние (см. стр. 23).
В
общем виде последовательность явлений,
протекающих
в рецепторах при
возбуждении, может быть представлена
в виде
следующей схемы (рис. 2). Конечно,
далеко не все элементы
9![]()
схемы
присущи каждому чувствительному органу.
У одних рецепторов не обнаружены
вспомогательные структуры, у других
не найден эфферентный нейрон, у третьих
предполагается возможность
эфаптической, а не синаптической
передачи возбуждения с рецептирующей
клетки на сенсорный нейрон (пунктирная
линия на схеме) и т. д. Наконец, у первичных
рецепторов отсутствуют рецептирующие
клетки.
Постсинаптические
мембраны нейронов фактически являются
высокоспециализированными хеморецепторными
поверхностями, поэтому последовательность
протекающих в них процессов принципиально
такая же, как и в любом первичном
хеморецепторе (Grundfest, 1959, 1965), что и
отражено на рис. 2.
ОБЩАЯ
ХАРАКТЕРИСТИКА МЕСТНЫХ И РАСПРОСТРАНЯЮЩИХСЯ
ОТВЕТОВ РЕЦЕПТОРОВ
Основные
сведения о РП были получены в опытах
на одиночных первичных механорецепторах:
мышечных веретенах, рецепторах
растяжения, тельцах Пачини (подробнее
см.: Katz, 1950; Granit, 1955; Kuffler, 1958; Gray, 1959 a, b;
Loewenstein, 1959, 1961, 1965; Ильинский, 19666—г,
1967).
Наиболее
отличительными чертами РП по сравнению
с потенциалом действия являются
зависимость его параметров (амплитуды,
длительности, скорости нарастания и
спадения и т. д.) от различных
характеристик раздражающего воздействия,
а также декрементный характер
распространения.
Амплитуда
и скорость нарастания РП по мере
увеличения силы раздражения градуально
возрастают до некоторого максимального
значения. При прочих равных условиях
они зависят и от скорости нарастания
раздражения. У телец Пачини такая
зависимость наблюдается при скоростях
ниже 1—2 мкм/мсек., а у мышечного веретена
— при скоростях менее 200 мкм/мсек.
Количественная
зависимость амплитуды деполяризационного
РП от силы раздражения у многих рецепторов
выводится из соотношения между
частотой импульсации и силой воздействия,
так как было показано, что между величиной
стационарной деполяризации и
спайковой активностью имеются линейные
отношения (Katz, 1950; Fuortes, 1959; Rushton, 1959;
Terzuolo, Washizu, 1962 и др.). Было установлено,
что у мышечных веретен, рецеп-
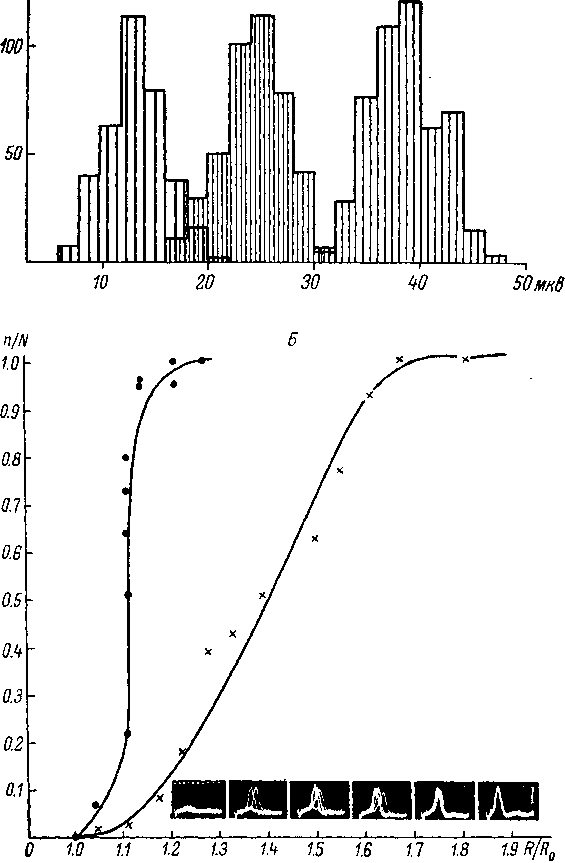
Рис.
3. Флуктуация амплитуды ГП (А) и порога
возникновения импульсной
активности
(2?) в тельцах Пачини (А — Loewenstein, 1961; В
—
Ильинский,
Фикс, 1963).
На
А: по
оси абсцисс —
амплитуда ГП (в мкв) при трех различных
силах раздражения; по
оси ординат —
число зарегистрированных ответов. На
J5: по
оси абсцисс — R/Ro,
R —
сила раздражения, Ro
—
максимальная сила раздражения, при
которой ответы еще отсутствуют; по
оси ординат —
вероятность появления ответа (n/N),
где п—
число
раздражений, а N
—
число ответов; кружки
—
реакция нервного волокна при раздражении
его электрическими импульсами; крестики
—
реакция тельца Пачини при раздражении
его механическими толчками; внизу
справа —
осциллограммы ответов телец Пачини;
отметка времени 2.0 мсек.; калибровка
650 мкв,
10
150г
А
торов
растяжения ракообразных, фоторецепторов
беспозвоночных и некоторых других
рецепторов между силой раздражения и
частотой импульсации (в среднем диапазоне
нагрузок) имеется логарифмическая
зависимость, что соответствует известному
закону Вебера—Фехнера (см.: Granit,
1955). Однако встречаются рецепторы,
например тельца Пачини, у которых кривая
зависимости амплитуды локальной
деполяризации от величины воздействия
имеет скорее S-образный вид.
Амплитуда
РП, несмотря на постоянство адекватного
раздражения, меняется. Это обусловлено
явлением флуктуации возбудимости
нерегенеративных структур рецептора
(рис. 3, А). Суммарная флуктуация
возбудимости рецептора определяется
в основном этими изменениями РП, а также
колебаниями порога возникновения
спайковой активности (рис. 3, Б),
т. е. флуктуациями возбудимости
регенеративной системы генерации
(Loewenstein, 1961; Ильинский, Фикс, 1963).
Максимальная
амплитуда возбуждающего РП, как это
следует из экспериментов на рецепторе
растяжения ракообразных, близка к
величине потенциала покоя и равняется
40—70 mb
(Terzuolo, Washizu, 1962; Obara, 1968). В отношении телец
Пачини было показано, что максимальное
значение ГП близко к величине потенциала
действия, возникающего в рецепторе
(Gray, 1959а; Ильинский, 1963). Однако нужно
учесть, что амплитуда потенциала
действия в районе немиелинизированного
нервного окончания меньше, чем на
некотором расстоянии от рецептора.
Можно полагать, что максимальная
величина ГП телец Пачини порядка 60 мв
(Nishi, Sato, 1968), а следовательно, она также
близка к значению потенциала покоя
рецептора (в последнее время в опытах
с внутриклеточным отведением от
волосковых механорецепторов боковой
линии было установлено, что максимальная
амплитуда РП может быть очень небольшой
— всего лишь 800 мкв (Harris et al., 1970).
В
РП обычно можно различить два компонента:
быстрый и медленный (рис. 4, А). Быстрый
возникает в момент нанесения раздражения
(on-ответ), а медленный — в период его
стационарного1
действия.
Соотношение между ними, равно как и
временное течение РП, определяется
при прочих равных условиях адаптационными
свойствами рецепторов. У быстро
адаптирующихся рецепторов, таких
как тельца Пачини, медленный компонент
ответа практически отсутствует (рис.
5, А). В свою очередь у более медленно
адаптирующихся структур, например, у
мышечного веретена, быстрый компонент
может быть сведен к минимуму при
небольших скоростях нарастания
раздражения (рис. 4, Б). По своему знаку
on-ответ может быть как де-, так и
гиперполяриза- ционным* (рис. 5).
Помимо
динамической реакции на включение
раздражения у рецепторов часто возникает
и ответ на выключение воздействия —
12
off
ответ. Он также может^быть как Де-, так
и гиперполяризацион- ним (рис. 5). Механизмы
возникновения off-реакции изучены еще
Рис.
4. РП мышечного веретена (Otto-
son,
Shepherd, 1965).
А
—
ответы рецептора до (а) и после (б)
дей-
ствия 0.2%-го лигнокаина,
блокирующего
импульсную активность;
масштаб времени —
50 мсек. Б
—
изменения динамической и
статической
компоненты РП при различных
скоростях
растяжения [от 70 мм/сек. (а)
до
0.55 мм/сек. (г)]. Конечная величина
растяжения
во всех записях 170 мкм, масштаб
времени
10 мсек, (для а, б) и 100 мсек, (для
в,
г); калибровка 0.5 мв. На А и Б вверху
—
запись
ответов рецептора, внизу
—■
запись
раздражения.
Рис.
5. Ответы телец Пачини
(А,
В)
и мышечного веретена
(Б)
на включение и выключе-
ние р
аздражения.
А
—
деполяризационные I on- и
off-РП. Ответ
отсутствует в момент
действия
статической составляющей
раздражения
(Ильинский, 1966а).
Б — деполяризационный
оп-РП
(отклонение луча вниз) и
гипер-
поляризационный off-P I
(Katz,
1950). В
—
гиперполяризационный
on-РП и
деполяризационный off-PH
и спайк
(Ильинский, 1966в).
На всех кадрах еверху
запись
формы механического
стимула.
Масштаб времени (в мсек.):
А, В —
4.0; Б — 100; калибровка (в мкв):
А
— о; В — 30.
недостаточно.
У механорецепторов они, например, могут
быть п значительной степени обусловлены
механическими свойствами пспомогательных
структур (см. стр. 41).
13
После
того как ГП достигает некоторой
критической амплитуды, возникают
спайки. У медленно адаптирующихся
рецепторов растяжения ракообразных
это значение критической деполяризации,
например, составляет 8—12 мв, т. е. 10—15%
от величины потенциала покоя (Eyzaguirre,
Kuffler, 1955). Обычно местом генерации ответов
«все или ничего» является начальный
участок аксона, где порог возникновения
импульсной активности наименьший.
Однако в принципе спайки могут возникать
и в других, менее чувствительных к
электрическому току областях рецептора
(Hunt, Takeuchi, 1962; Ozeki, Sato, 1964; Grampp, 1966, и др.). В
некоторых случаях электрогенные,
регенеративные структуры, по-видимому,
могут быть мозаично распределены среди
собственно рецептирующих, нерегенёративных
элементов рецепторной мембраны.
Рис.
6. Изменение РП и импульсной активности
медленно адаптирующегося рецептора
растяжения ракообразных при шагообразном
варьировании силы раздражения
(Eyzaguirre, Kuffler, 1955).
Масштаб
времени 1.0 сек., калибровка 20 мв; стрелками
показаны соответственно
начало
возрастания и уменьшения раздражения.
По
мере возрастания амплитуды ГП наблюдается
увеличение числа и частоты разрядов,
но это происходит только до определенного
предела, выше которого наступает полное
торможение импульсной активности
(католический блок) (рис. 6). В зависимости
от аккомодационных особенностей
регенеративной системы генерации этот
блок может возникнуть как при очень
небольших величинах и длительностях
действия локальной деполяризации
(тельца Пачини), так и при весьма
значительных амплитудах и продолжительностях
стационарной деполяризации (рецепторы
растяжения). Если рецептор в течение
продолжительного времени генерирует
импульсную активность, то часто после
выключения раздражения наблюдается
посттетаническое (анодическое) угнетение
(Nakajima, Takahashi, 1966). При ритмическом,
прерывистом раздражении рецепторов
изменение импульсной активности
происходит по тем же правилам, как и в
других возбудимых образованиях.
Также имеются оптимальные и пессимальные
для возбудимого субстрата частоты
раздражения, наблюдается явление
усвоения ритма и т. д.
14
ИОННЫЕ
МЕХАНИЗМЫ И ДЕЙСТВИЕ НЕКОТОРЫХ ВЕЩЕСТВ
Ионные
механизмы генерации активности в
рецепторах до настоящего времени
изучены крайне недостаточно, что в
значительной мере объясняется
методическими трудностями. Кроме
того, анализ действия ионов и различных
ядов даже на первичные рецепторы весьма
затруднен, так как местом их
приложения
могут оказаться разные образования:
1) сосуды данной рецепторной области;
2) вспомогательный аппарат рецептора,
а также окружающие его тканевые
структуры; 3) собственно рецептирующий
субстрат сенсорного прибора и связанная
с ним система генерации РП; 4) регенеративная
система генерации пиковых потенциалов.
У вторичных рецепторов наличие контактов
между рецептирующей клеткой и сенсорным
нейроном создает еще дополнительные
точки действия химических агентов.
Экспериментов
с регистрацией РП пока еще немного. В
основном они проведены на
механорецепторах (подробнее см.: Paintai,
1964; Ильинский, 1967). Возникновение
возбуждающего РП в механорецепторах
зависит от нескольких ионов. Так, было
установлено, что равновесный уровень
РП у рецептора растяжения ракообразных
колеблется, по данным разных авторов,
от 0 до —30 мв, что говорит об участии
нескольких ионов в формировании
деполяризационного РП (Terzuolo, Washizu, 1962;
Obara, 1968). По-видимому, можно рассматривать
рецепторную мембрану рецептора
растяжения в активном состоянии как
относительно неспецифическую
катион-селективную структуру. Основную
часть заряда при активации рецепторной
мембраны переносят ионы Na+.
В безнатриевых растворах амплитуда ГП
рецептора растяжения и тельца Пачини
падала до 90%, а мышечного веретена —•
до 70—80%. Аналогично амплитуде изменялась
и скорость нарастания ГП. При замене
безнатриевых растворов на обычные
активность рецепторов хорошо
восстанавливалась. Ион Li+
мог заместить ион Na+,
однако амплитуда возбуждающего РП в
этом случае была меньше нормальной
(Sato et al., 1968; Obara, Grund’fest, 1968).
В
генерации ГП принимают участие и ионы
К+.
Ионы же С1", по-видимому, в формировании
ГП не участвуют (Obara, 1968).
Удаление
из раствора ионов Са++
ведет к уменьшению амплитуды
деполяризационного РП мышечного
веретена и рецептора растяжения.
Одновременно с этим понижается порог
возникновения спайков, причем второе
явление превалирует над первым, в
результате чего фоновая активность
рецепторов увеличивается и
пороги
на адекватное раздражение снижаются.
На фоне действия бескальциевых
растворов резко увеличивается
возбуждающее действие ионов К+.
Избыток Са++
либо не сказывается на РП,
15
либо
при больших концентрациях ведет к
прогрессивному снижению его амплитуды.
Пороги же возникновения спайков
возрастают. Ионы Mg++
уменьшают или полностью препятствуют
влиянию бескальциевых растворов.
Действию
различных веществ на рецепторы посвящено
огромное число работ (см. обзоры:
Черниговский, 1960; Paintai, 1964; Лебедева,
1965; Ильинский, 1967, и др.), и систематическое
их рассмотрение выходит за рамки
этой статьи. Здесь мы коснемся только
влияния некоторых агентов.
Такое
биологически активное вещество, как
ацетилхолин, часто вызывает возбуждение
рецепторов или же снижение их порогов
к адекватному раздражению. У таких
образований, как первичные механорецепторы,
этот эффект обусловлен в значительной
степени деполяризацией регенеративной
системы. У вторичных же рецепторов
действие ацетилхолина может быть
связано с его участием в передаче
возбуждения от рецептирующей клетки
к сенсорному нейрону.
Адреналин
и норадреналин увеличивают амплитуду
и скорость нарастания ГП и снижают
пороги возникновения потенциалов
действия. Большие дозы адреналина или
длительное раздражение симпатического
нерва после периода возбуждения
механорецепторов затем их угнетают.
В значительной мере такой эффект
обусловлен влиянием адреналина на
вспомогательные структуры рецепторов,
а также на сосуды данной области.
Анестетики
в умеренных концентрация;х полностью
угнетают импульсную активность
рецепторов, сохраняя при этом РП. Так
же действует и тетродотоксин. При этом
он сохраняет не только деполяризационные,
но и гиперполяризационные РП (Nishi, Sato,
1968).
РИТМИЧЕСКАЯ
АКТИВНОСТЬ
За
исключением чрезвычайно быстро
адаптирующихся нервных окончаний,
для всех рецепторных приборов является
очень характерной генерация ритмического
разряда импульсов в ответ на действие
раздражителя, ибо только таким способом
большинство из них может информировать
центральную нервную систему о
различных параметрах внешнего
воздействия (см. ниже — «Периферическое
кодирование»). Кроме того, многим
рецепторам свойственна ритмическая
активность даже в состоянии покоя
(«спонтанная» импульсация). Поэтому
ритмическую активность можно считать
одной из фундаментальных особенностей
деятельности рецепторных приборов.
Изучение
природы ритмической импульсации было
начато в классических работах Эдриана
(Adrian, 1928, 1932) и в дальнейшем шло по
двум основным направлениям: по пути
анализа меха
16
низмов
аккомодации нерва и исследования
механизмов, определяющих интервал
между спайками.
Многочисленными
исследованиями (Erlanger, Blair, 1938; Hodgkin,
1948, и др.) было показано, что далеко не
все нервные волокна являются быстро
аккомодирующимися структурами. Поэтому
при действии постоянного тока во многих
из них может возникать ритмический
разряд, частота и длительность которого
воз-
Рис.
7. Ответы одиночного нервного волокна
краба при действии катода
постоянного
тока разной силы (Hodgkin, 1948).
Цифры
—
величина тока в реобазах; моменты
включения и выключения раздражения
показаны артефактами. Внизу
—
время в сек.
растают
при увеличении амплитуды тока (рис. 7).
Эти данные позволили предположить, что
механизм ритмической активности может
быть понят на основе общих свойств
нервных волокон. Разрабатывая эту идею,
шведские нейрофизиологи (Bernhard et al.,
1942; Granit, 1947) сформулировали общепринятую
сейчас и рассмотренную выше концепцию
«генераторного потенциала».
Для
объяснения механизмов, определяющих
величину межспайковых промежутков,
т. е. частоту разряда, первоначально
было выдвинуто представление о ведущей
роли изменений рефрак- терности субстрата
после возникновения импульса (Adrian,
1928). Однако с позиции этой гипотезы было
трудно объяснить низкочастотную
активность рецепторов. В этом случае
межспайковый интервал был много дольше
времени полного восстановления
возбудимости.
2
Сенсорные системы
17
Эксперименты
на нервных волокнах ракообразных
позволили выдвинуть другое представление,
согласно которому интервал между
спайками определяется теми ясе причинами,
что и латентный (скрытый) период,
необходимый для возникновения нервного
импульса в момент включения раздражения
(Hodgkin, 1948). Оба эти промежутка времени
определялись развитием локального
ответа: когда последний достигал
критического уровня, возникал спайк.
Указанная
гипотеза хорошо объясняла характер
изменения частоты при низких силах
раздражения. При больших же раздражающих
воздействиях частота разряда зависела
скорее от рефрактерного периода,
так как латентный период реакции мог
быть в этом случае очень маленьким, а
межспайковый интервал никогда не был
менее нескольких миллисекунд.
Эксперименты
на мышечном веретене (Katz, 1950) и на
рецепторе растяжения ракообразных
(Eyzaguivve, Kuffler, 1955) в целом подтвердили
изложенные выше представления о
механизмах генерации импульсной
активности в рецепторах.
СПОНТАННАЯ
ИМПУЛЬСАЦИЯ
Как
уже отмечалось, для многих рецепторных
структур, например таких, как
волосковые механорецепторы полукружных
каналов и боковой линии, некоторые
хемо- и электрорецепторы и т. д.,
характерно наличие фоновой активности.
Она имеет важное значение в деятельности
сенсорных систем, так как позволяет
центрам с помощью одних и тех же
рецепторов оценивать действие как
возбуждающих, так и тормозных
раздражителей (рис. 1, Б), а также
поддерживать свою высокую возбудимость.
Спонтанная
активность отражает ряд важных
особенностей в деятельности рецепторных
приборов. Она, по-видимому, связана
с флуктуационными изменениями мембранного
потенциала, спонтанно происходящими
в рецепторных клетках, что находит свое
отражение в описанных ранее колебаниях
амплитуды РП. Аналогичные изменения
потенциала хорошо известны в различных
нервных структурах (Derksen, 1965; Firth, 1966).
Для того чтобы спонтанные изменения
ГП могли вызвать появление импульсных
разрядов, порог возникновения возбуждения
в рецепторах должен быть очень
низким, что и наблюдается в действительности.
Чрезвычайно низкие пороги возникновения
возбуждения, существующие практически
у всех высокоорганизованных рецепторов,
а также наличие у многих из них спонтанной
активности заставляют предполагать
стационарно неустойчивое состояние
всех возбудимых систем этих сенсорных
аппаратов.
В
целом спонтанная активность рецепторов
изучена еще совершенно недостаточно,
АДАПТАЦИЯ
Процесс
адаптации, выражающийся в уменьшении
активности рецепторов по мере действия
стационарного раздражителя, является
очень характерной чертой сенсорных
структур (Гершуни, 1940). По своей
адаптационной способности все рецепторы
независимо от их строения и модальности
могут быть подразделены на быстро
и медленно адаптирующиеся (соответственно
фазные и тонические). Можно говорить
также и о промежуточном типе:
фазно-тонических рецепторах. Проще
всего проследить сущность адаптационных
явлений на примере первичных рецепторных
приборов, таких как рецепторы растяжения,
тельца Пачини, мышечные веретена, что
и было осуществлено в последние годы
(см. подробнее: Ильинский, 1966а, 1966г,
1967).
Адаптационные
процессы в первичных рецепторах, о
развитии которых обычно судят по
исчезновению или уменьшению импульсной
активности, могут определяться
теоретически тремя основными
факторами: 1) свойствами вспомогательного
аппарата; 2) особенностями собственно
воспринимающих структур рецептора;
3) свойствами регенеративных элементов
нервного окончания, ответственных
за возникновение спайков (у вторичных
рецепторов, кроме того, могут иметь
значение и те явления, которые
развертываются в месте синаптического
контакта между рецепти- рующей клеткой
и окончаниями сенсорного нейрона).
Поэтому анализ явления адаптации
фактически сводится к оценке удельного
веса каждого из этих факторов в развитии
суммарной ответной реакции сенсорного
прибора на действие адекватного
раздражителя.
В
зависимости от характера воздействия
можно различить два вида адаптационных
процессов. В одном случае раздражителем
является стационарное, непрерывистое
воздействие (например, для механорецепторов
— это стационарное растяжение, сдавление
и т. д.). В другом случае раздражителем
является постоянное, но прерывистое
воздействие (например, вибрация).
Резкое
растяжение как быстро, так и медленно
адаптирующихся рецепторных нейронов
ракообразных ведет к всплеску импульса-
ции в момент включения раздражения.
При этом чем больше скорость нарастания
стимула и его амплитуда, тем выше частота
импульсной активности. Возросшая в
первый момент активность очень быстро
затухает и у быстро адаптирующихся
рецепторов скоро совсем исчезает. У
медленно же адаптирующихся нейронов
импульсация устанавливается на некотором
постоянном уровне, зависящем от величины
стационарного растяжения, и может
сохраняться в таком состоянии часами
(рис. 8, А).
Удалось выяснить, что первоначальное
уменьшение импульсации у обоих видов
рецепторов было обусловлено исключительно
механическими свойствами вспомогательных
структур; частота разрядов падала
2*
19

параллельно
уменьшению напряжения рецепторной
мышцы после ее растяжения (Wendler, 1963).
Различие же в поведении быстро и медленно
адаптирующихся рецепторов в период
действия стационарной составляющей
раздражения оказалось связанным с
особенностями их регенеративных
структур (Nakajima, 1964; Nakajima, Onodera, 1969а). Так,
при действии постоянного тока медленно
адаптирующийся рецептор генерировал
импульсы в течение всего времени
приложения раздражителя, в то время
как быстро адаптирующийся нейрон
отвечал разрядами не более 30 сек.
Локальная же деполяризация (ГП), вызванная
растяжением, в обоих случаях могла
длиться одинаково долго (рис. 8, Б).
Таким образом, собственно воспринимающий
субстрат рецептора и связанная с ним
нерегенеративная система генерации
оказались медленно адаптирующимися к
действию раздражителя. Поэтому феномен
адаптации у быстро адаптирующихся
рецепторов, фактически полностью
определялся свойствами вспомогательных
структур и регенеративной системы
генерации. Вместе с тем те адаптационные
явления, которые наблюдаются у медленно
адаптирующихся нейронов, по-видимому,
полностью зависят от процессов,
развивающихся именно в нерегенеративной
системе генерации (Nakajima, Onodera, 1969b).
Принципиально
такой же вывод можно было сделать и в
отношении других механорецепторов.
Так, у крайне быстро адаптирующихся
телец Пачини феномен адаптации к
действию стационарного, непрерывистого
раздражителя определялся исключительно
свойствами их вспомогательных структур
(капсул рецепторов), которые не пропускают
к нервному окончанию стационарной
составляющей стимула (Hubbard, 1958;
Loewenstein, 1965; Ильинский, 1965, 1966г). После
же удаления капсулы рецептор начинает
генерировать РП в течение всего времени
действия стимула (рис. 9, Л). Однако и у
декапсулированных рецепторов импульсная
активность продолжает возникать только
в самый первый момент включения
раздражения, что связано с высокой
аккомодационной способностью
регенеративных структур (рис. 9, В, а).
Несколько иная картина наблюдается
при действии на интактные
Рис.
8. Процессы адаптации в рецепторах
растяжения ракообразных.
А
— изменение напряжения рецепторной
мышцы и импульсной активности медленно
адаптирующегося рецептора при включении
(а) и выключении (б) длительного
механического раздражения (Wendler,
1963). По
оси абсцисс —
время в сек.; по
оси ординат — частота
импульсации и величина напряжения в
процентах к исходному уровню. Б
— изменение
импульсной активности и ГП в рецепторах
растяжения при действии продолжительных
стимулов (Nakajima, 1964). Слева
—
изменение импульсной активности
медленно (а,, а2)
и быстро (бх—б4)
адаптирующихся рецепторов под действием
постоянного деполяризующего тока; а,
— длительность тока 140 сек.; а2
—
большой ток действовал 26 сек. Справа
—
изменение ГП, вызванного действием
продолжительного растяжения. Импульсная
активность блокирована тетродотоксином
(10~7
г/мл). Подъем ГП соответствует началу
раздражения, падение ГП — концу
воздействия. Три медленно (а,—а3)
и
три быстро (б,—б3)
адаптирующихся нейрона. Масштаб времени:
внизу
слева —
1 сек.; внизу
справа —
5 сек. Калибровка (внизу)
—
20 мв; масштаб силы раздражающего тока
10~8
А.
21
тельца
Пачини длительных ритмических
раздражителей. В этом случае основной
причиной адаптации являются уже не
свойства вспомогательных структур, а
только аккомодационные особен-
Рис.
9. Адаптация телец Пачини к действию
длительного непрерывного
и прерывистого
раздражения.
А
—
ответы интактного (а) и декапсулированного
(б, в) телец Пачини на действие длительного
механического толчка (Ильинский, 1966а).
На а
верхний луч,
на б,
в нижний луч — запись
раздражающего воздействия. Б
—
изменение ГП интактного рецептора при
раз- < личных (а—г)
частотах ритмического раздражения
(Loewenstein, 1958). Верхний
луч — ГП:
второй
луч сверху —
отметка раздражения; на г
нижний луч —
отметка времени. В
—
ГП и импульсная активность (верхняя
запись)
декапсулированного (а)
и интактного (б) телец Пачини на разные
виды механических стимулов (нижняя
запись)
(Loewenstein, Mendelson, 1965). Г
—
изменение импульсной активности (нижний
луч)
интактного тельца Пачини при действии
длительного прерывистого раздражения
(Ильинский, 1966а). Сила раздражения всюду
одинаковая; частота раздражения: на 1
—
100 в 1 сек.; на 2—9
— 650
в 1 сек.; кадры 2-й—6-й
соответствуют 2-й, 7-й, 10-й, 15-й, 20-й сек.
раздражения. Кадры 7-й—9-й
соответствуют 5-й, 40-й, 120-й сек. после
окончания тетанизации. Раздражение
на 7—9
производилось только в момент съемки.
На Г: отметка раздражения — верхний
луч\
отметка времени — второй
луч сверху.
Масштаб времени (в мсек.): на А
— 6.0; В —
5.0; Г
—
20.0; отметка времени (в мсек.) на Б
—
1.0; калибровка (в мкв): А
— 40.0;
Б
—
25.0; В
— 50.0; Г
— 200.0.
пости
его нервного волокна. При больших
частотах раздражения отдельные ГП
сливаются в сплошное плато (рис. 9, В,
г) и
регенеративные элементы рецептора
инактивируется практически
22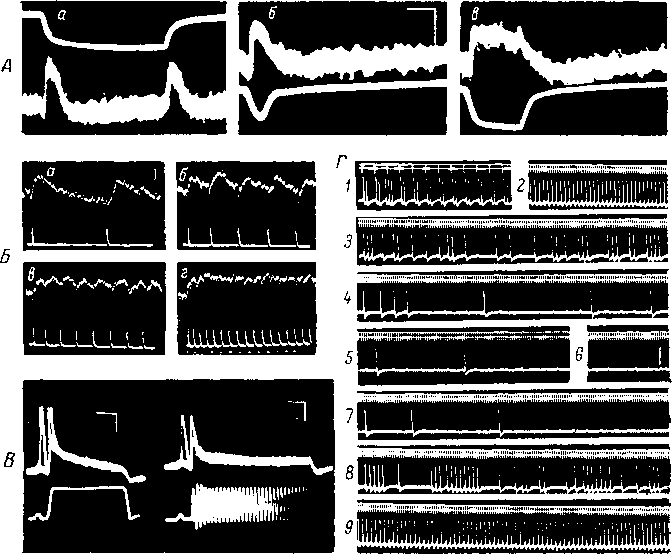
сразу
(рис. 9, Б, б).
При меньших частотах отдельные ГП
суммируются лишь частично (рис. 9, Б,
а—в),
поэтому угнетение импульсной
активности развивается постепенно
(рис. 9, Г).
Важную
роль в процессах адаптации могут играть
и эфферентные влияния от расположенных
выше нервных центров.
ЭФФЕРЕНТНАЯ
РЕГУЛЯЦИЯ
Установлено,
что большинство рецепторных структур
находится под контролем центральных
нервных образований (Granit, 1955; Livingston,
1959; Hagbarth, 1960; Снякин, 1961; Hensel, 1966; Есаков,
1966). Этот эфферентный контроль над
рецепторными аппаратами осуществляется
в разных случаях по- разному. Чаще всего
сенсорные приборы находятся под
некоторым тоническим тормозным влиянием.
Механизм
эфферентной регуляции на уровне
одиночных рецепторов изучен еще
очень мало. Одними из наиболее
обследованных в этом плане оказались
рецепторы растяжения ракообразных, у
которых .деятельность эфферентных
нервных волокон была продемонстрирована
как в опытах на изолированных препаратах,
так и в экспериментах на целых животных
(Kuffler, 1958; На- giwara et al., 1960; Eckert, 1961a, b; Fields
et al., 1967, и др.). Тормозные волокна рецептора
растяжения оканчиваются в основном
на дендритах сенсорного нейрона, т. е.
в области, где происходит восприятие
механического раздражения и генерируется
РП. Раздражение тормозного нерва
вызывает небольшие деполяризационные
изменения потенциала в покоящемся
рецепторном нейроне. По мере растяжения
рецептора и снижения величины мембранного
потенциала этот деполяризационный
потенциал уменьшается, а затем
извращается, превращаясь в
гиперполяризационный, амплитуда
которого возрастает при увеличении
растяжения. Таким образом, действие
тормозного нерва заключается в
стабилизации мембранного потенциала
около некоторой величины (равновесного
тормозного потенциала), значение которой
варьирует у разных препаратов. Благодаря
такой стабилизации потенциала клетки
ГП не достигает критического значения
и спайки в рецепторе не возникают.
Тормозные
волокна стационарно выделяют из своих
окончаний медиатор, что обнаруживается
по наличию миниатюрных потенциалов
в рецепторной клетке (Iwasaki, Florey, 1969).
Природа тормозного медиатора пока не
ясна. Наличие ионных механизмов генерации
тормозного потенциала показало, что в
основе его лежит изменение проницаемости
мембраны рецептора к ионам К +
и G1".
Наличие
эфферентного контроля над кожными
рецепторами было показано рядом авторов.
В частности, оказалось, что раздражение
симпатических нервов, идущих к коже,
снижает пороги, уменьшает утомление и
замедляет скорость адаптации тактильных
??
рецепторов
(Loewenstein, 1956; Chernetski, 1964; Дмитриева, 1967, и
др.). При раздражении же симметричных
участков кожи обычно наблюдалось
угнетение тактильных рецепторов
(Дмитриева, Еса- ков, 1966; Есаков, Дмитриева,
1968). Это торможение имело рефлекторную
природу и осуществлялось при помощи
как соматических, так и симпатических
путей.
Наряду
с центральными образованиями на
деятельность различных рецепторов
(например, фоторецепторов, тактильных
рецепторов и др.) могут оказывать влияние
и расположенные непосредственно рядом
с ними другие рецепторные структуры.
Это явление получило название
периферического взаимодействия.
Механизмы такого взаимодействия
могут быть различными. В случае, например,
тактильных рецепторов (Есаков,
Дмитриева, 1967) оно скорее всего возникает
в результате антидромного распространения
импульсов от возбужденных рецепторов
к невозбужденным.
Периферическое
взаимодействие, эфферентные влияния
от центров и «спонтанные» колебания
возбудимости самих рецепторов,
по-видимому, и являются причиной
непрерывного изменения числа работающих
элементов сенсорной поверхности
(явления функциональной мобильности
чувствительных приборов — см. Снякин,
1959).
ПЕРИФЕРИЧЕСКОЕ
КОДИРОВАНИЕ
Вопрос
о связи различных параметров раздражающего
воздействия с импульсной активностью,
генерируемой рецепторами, является
одним из наиболее важных при изучении
деятельности сенсорных аппаратов. Ведь
в конечном счете только анализируя
последовательность поступающих от
рецепторов импульсов, центральная
нервная система составляет представление
о характеристиках падающих на организм
раздражителей. Среди изученных рецепторов
наибольшее число способов периферического
кодирования обнаружено у электрорецепторов
(Hagiwara, Morita, 1963; Bullock, Chichibu, 1965; Suga, 1967, и
др.).2
Наиболее
распространенным среди различных
рецепторов способом кодирования
интенсивности адекватного стимула
является изменение частоты импульсации
(Adrian, 1928; Granit, 1955).
Существует
большое количество разновидностей
частотного кодирования. Это определяется
разнообразием функциональных особенностей
рецепторов. Одни из них, например,
обладают фоновой активностью, а
другие нет. Первые могут оценивать не
только возбуждающий раздражитель
(увеличением частоты импульсации), но
и тормозное воздействие (уменьшением
частоты разрядов).
2
Основные характеристики этого класса
рецепторов, открытого сравнительно
недавно у рыб и не рассматриваемого в
данном руководстве, изло- женыТв ряде
обзорных работ (Machin, 1962; Lissmann, 1963; Bennett,
1965, 1967/1970; Murray, 1967; Lissmann, Mullinger, 1968, и др.).
24
Вторые
же способны оценить только лишь
возбуждающие стимулы. В зависимости
от того, является ли рецептор быстро
или медленно адаптирующейся структурой,
реагирует ли он на включение или
выключение раздражения и т. д., рисунок
ответной реакции может быть у разных
сенсорных приборов совершенно различным,
Примером
одной из разновидностей частотного
кодирования, обнаруженной сравнительно
недавно, может быть так называемое
обусловленное частотное кодирование
(Bullock, Chichibu, 1965). Оно заключается в том,
что фоновая ритмическая импульсация
рецепторов плавно возрастает или
снижается и затем остается на новом
уровне в зависимости от увеличения или
ослабления раздражения.
Другим,
довольно распространенным способом
кодирования является численное
кодирование. В этом случае сила стимула
оценивается числом возникающих в
рецепторе импульсов. Чем больше
максимальное число генерируемых
разрядов, тем более тонко рецептор
может оценивать внешнее воздействие.
Наряду с числом импульсов (или
длительностью вспышки), по-видимому,
может иметь значение и временная
микроструктура последовательности
спайков (межспайковые вариации).
Весьма
своеобразный способ кодирования
наблюдается у электрорецепторов
Gymnotus carapo (Suga, 1967). Ряд электрорецеп-
торных волокон этих животных обладает
в состоянии покоя стационарной
ритмической активностью, которая может
синхронизоваться разрядами
электрического органа. В случае, если
амплитуда разрядов падает или же
изменяется их частота, наблюдается
десинхронизация фоновой активности.
Внешний объект, попадая в электрическое
поле рыбы, меняет величину тока,
протекающего через рецепторы и вызывает
тем самым явление десинхронизации, что
и является сигналом для центральной
нервной системы. Наличие волокон с
различной частотой фоновой активности
и с различными порогами приводит к
тому, что раздражитель, вызывая
десинхронизацию у одних рецепторов,
наоборот, синхронизует ак тивность
других. Это, естественно, расширяет
информационные возможности системы.
Другим
способом оценки силы воздействия,
встречающимся у некоторых слабоэлектрических
рыб, и в частности у рыб, обладающих
высокочастотными разрядами электрических
органов (у некоторых Gymnotid число разрядов
может доходить до 1600 в 1 сек.), является
вероятностное кодирование. В покое
электрорецепторы таких рыб в ответ
на разряды электрического органа
генерируют импульсы очень нерегулярно,
с некоторой вероятностью. По мере
же увеличения силы воздействия
вероятность появления спайков плавно
увеличивается, что и позволяет животному
точно оценивать величину раздражающего
стимула.
У
некоторых слабоэлектрических рыб
обнаружены электрорецепторы,
генерирующие всего лишь один-два
импульса на каждый
25
разряд
электрического органа. В этом случае
изменения, вызываемые в электрическом
поле около рыбы, кодируются скрытым
периодом реакции (Bullock, Chichibu, 1965; Szabo,
1967, и др.).
В
состоянии покоя ответы элек-
трорецепторов
на разряды элек-
трического органа
возникают
с определенным,
неизменным
латентным периодом. При
по-
падании же в электрическое
поле
рыбы какого-либо предмета
плотность
токов, текущих че-
рез рецепторы,
меняется, что
и вызывает
соответствующие
изменения скрытого
периода
реакции (рис. 10). Центральная
нервная
система, зная момент
возникновения
разряда элект-
рического органа,
может точно
оценивать характер
воздейст-
вия по величине скрытого
пе-
риода ответов электрорецепто-
ров.
По-видимому, подобное
латентное
кодирование может
встречаться и в
других сен-
сорных системах, имеющих
сво-
его рода точки отсчета (нали-
чие
пейсмекерных структур).
Различные
способы кодиро-
вания часто сочетаются.
На-
пример, электрорецепторы Gym-
notus
наряду с варьированием
числа импульсов
могут исполь-
Рис.
10. Кодирование скрытым пе-
риодом у
одиночного электрорецеп-
тора (Szabo,
1967).
Каждый
разряд электрического органа вызывает
появление двух импульсов в рецепторе.
По мере продвижения пластинки из
пластика параллельно длинной оси тела
рыбы латентный период реакции плавно
меняется. Величина перемещения пластинки
равняется расстоянию между
вертикальными линиями. Слева
вверху —
исходное положение (линии совмещены).
Справа
внизу —
масштаб расстояния 5 см. Развертка луча
осциллографа запускается разрядом
электрического органа. Слева
внизу —
масштаб времени 2 мсек.
зовать
также изменения межспайковых интервалов
и латентных периодов (Suga, 1967).
Вероятностное кодирование в свою .
очередь может сочетаться с численным
кодированием (Hagiwara, Morita, 1963) и т. д.
Сенсорные системы одного
26