

направляется
вперед и вверх от оливы и подходит к
задним буграм четверохолмия. Среди
волокон латеральной петли располагаются
группы клеток, которые составляют
вентральное, дорсальное и интермедиальное
ее ядра. Вентральное ядро неоднородно,
основная его часть состоит из небольших
клеток округлой формы с петлевидными
синапсами. Ближе к оливе располагается
группа крупных клеток с крупными
синапсами в виде модифицированных
чашечек Хельда. Эта область по типу
синаптических окончаний сходна с
передней областью переднего кохлеарного
ядра. Дорсальное ядро латеральной петли
составляют небольшие клетки с мелкими
петлевидными синапсами. Такое же
строение имеет и интермедиальное ядро,
но оно выражено не у всех млекопитающих.
На клетках ядер латеральной петли
оканчивается часть волокон от дорсальных
кохлеарных ядер противоположной
стороны, идущих в составе дорсальной
слуховой полоски; на клетках дорсального
ядра оканчивается также часть волокон,
идущих от латеральной и медиальной
олив, но основную часть синапсов этих
ядер составляют окончания коллатералей
слуховых волокон. Часть аксонов клеток
ядер латеральной петли оканчивается
на клетках заднего бугра, другая
направляется во внутреннее коленчатое
тело. Небольшое число волокон латеральной
петли по всей ее длине направляется
через медиальную часть ствола на
противоположную сторону, образуя
так называемую коммиссуру Пробста.
Латеральная петля представляет собой
общий путь для аксонов нейронов II, III и
отчасти IV порядков и представляет
улитку как своей, так и противоположной
сторон. В ростральном направлении
число аксонов нейронов II порядка
уменьшается в результате переключения
части волокон на клетках ядер латеральной
петли. Латеральная петля входит в глубь
ядра заднего бугра четверохолмия, часть
ее волокон проходит без переключения
во внутреннее коленчатое тело.
Задние
бугры четверохолмия, иначе называемые
нижними буграми четверохолмия
(colliculus inferior, corporis quadrige- mini) или нижними
или задними холмами, а также нижним
двухолмием, представляют собой в
основном скопление нейронов IV порядка,
но в нем в значительной степени
представлены также нейроны III и V
порядков. Структура этого образования
очень сложна. Основную его часть
составляет массивное овальной формы
центральное ядро. Дорсо-латерально
по отношению к нему расположена
относительно небольшая клеточная
группа — латеральная кора заднего
бугра. Наружная поверхность бугра
покрыта тонким слоем белого вещества.
Между обоими буграми лежит интернуклеарная
кора, образующая спайку задних бугров.
Центральное ядро представлено густым
скоплением клеток различной формы и
величины.
201Задние бугры четверохолмия
Среди
них выделяются длинноаксонные клетки
мультиполярной и веретенообразной
формы и короткоаксонные звездчатые
клетки (Бехтерев, 1895; Ramon у Cajal, 1909; Левин,
1961; Зворыкин, 1963).
Аксоны
клеток заднего бугра выходят на боковую
поверхность и образуют крупный пучок
волокон — ручку заднего бугра (brachium
corporis quadrigemini posterioris), который на-
Рис.
63. Ход волокон латеральной петли (Ramon у
Cajal, 1909).
Пунктиром
обозначены ядра.
правляется
ко внутреннему коленчатому телу (рис.
63). Часть аксонов идет через спайку в
противоположный бугор и либо переключается
там, либо проходит в* ручку и идет в
противоположное коленчатое тело.
Таким образом, задний бугор дает как
ипси-, так и контрлатеральные проекции.
Среди
волокон ручки находятся рассеянные
клетки, которые составляют интерстициальное
ядро ручки. , Медиальнее по отношению
к волокнам ручки расположено
парабрахиальное ядро. Часть волокон
ручки оканчивается в этих ядрах.
ВНУТРЕННЕЕ
КОЛЕНЧАТОЕ ТЕЛО
Следующее
место переключения в восходящем
слуховом пути — внутреннее коленчатое
тело (corpus geniculatum mediale). Оно делится прежде
всего на главную (pars principalis) мелкоклеточную
и крупноклеточную (pars magnocellularis) части?
202
Мелкоклеточная
часть, через которую проходит основной
слуховой путь, неоднородна и
подразделяется на дорсальную и
вентральную части (Ramon у Cajal, 1911;
Зворыкин, 1952; Левин, 1959; Sychowa, 1962; Morest,
1964).
Рис.
64. Схема фронтального среза внутреннего
коленчатого
тела (Morest, 1965).
d
—
дорсальная, v
—
вентральная, т
—
медиальная области внутрен-
него
коленчатого тела.
Волокна
ручки заднего бугра четверохолмия при
входе во внутреннее коленчатое тело
разделяются на два пучка: медиальный
и латеральный. Медиальный пучок состоит
в основном из волокон латеральной
петли, которые, не переключаясь в заднем
бугре, проходят в медиальной части
ручки и входят в крупноклеточную
203
медиальную
часть внутреннего коленчатого тела.
Латеральный пучок содержит волокна,
идущие от клеток заднего бугра, и входит
в мелкоклеточную часть. Вскоре после
входа он разделяется на дорсальный и
вентральный пучки. Дорсальный пучок,
меньший по объему, входит в дорсальную
часть внутреннего коленчатого тела.
Вентральный пучок содержит основную
часть афферентных волокон и входит в
вентральную часть.
Для
дорсальной области характерны крупные
нейроны с толстыми дендритами,
отходящие от тела клетки в различных
направлениях, образуя сферическое
дендритное поле. Для вентральной
области основными являются нейроны с
пучковидными дендритами, которые
отходят от тела клетки только в одной
плоскости, образуя дендритное поле
дисковидной формы. В медиальной части
встречаются нейроны обоих типов, а
также нейроны промежуточной формы
(рис. 64). Вентральная область имеет
наибольшие размеры, и в ней оканчивается
наибольшая часть входящих волокон. В
этой области была обнаружена ламинарная
структура (Morest, 1964, 1965). Пучковидные
нейроны, налагаясь друг на друга своими
дисковидными дендритными полями,
образуют клеточный слой. Параллельно
этому клеточному слою проходят входящие
в эту область афферентные волокна ручки
заднего бугра. Каждое волокно дает
многочисленные коллатеральные ветви,
которые разветвляясь в виде виноградной
кисти, образуют обильное сплетение в
месте перекрытия дендритных полей
соседних клеток (Majorossy, Rathelvi, 1968). В этом
месте формируется область синаптических
контактов, синаптическая кисть,
образование, близкое по своей структуре
гломерулам наружного коленчатого тела
(рис. 65). Такие слои, или пластинки,
располагаясь параллельно друг другу,
занимают весь объем вентральной области.
В латеральной части они идут параллельно
боковой поверхности коленчатого тела
(латеральная часть), затем они закручиваются
в вентро-медиальном направлении, образуя
два завитка (овальная часть), затем
вновь раскручиваются и образуют в
медиальной части вертикальные пластинки,
вогнутые медиально.
Аксоны
клеток внутреннего коленчатого тела
направляются вначале вперед медиально,
а затем, поворачивая латерально, входят
в слуховую радиацию (Меринг, 1953). Основную
часть волокон слуховой радиации
составляют аксоны вентральной части
внутреннего коленчатого тела и в
меньшей степени аксоны клеток дорсальной
области. В медиальной области находится
много короткоаксонных нейронов, которые
связывают различные отделы коленчатого
тела.
СЛУХОВАЯ
КОРА
Волокна
слуховой радиации оканчиваются на
клетках слуховой коры, расположенной
в височной доле (lobus temporalis) (рис. 66;
Блинков, 1949, 1955; Мохова и др., 1959; Поляков,
1959, 1961; Woolsey, 1961).
204
Рис.
65. Схема связей в вентральной области
внутреннего колен-
чатого тела
(Majorossy, Rathelvi, 1968).
а
— клетки с дисковидным дендритным
полем; Ъ
—
афферентное волокно; с—
клетка
Гольджи II типа; d
—
синаптическая кисть.
Согласно
существующим анатомическим представлениям
о ходе восходящего слухового пути,
слуховой корой следует считать те
области коры, которые являются прямой
проекцией восходящего слухового пути,
т. е. удаление которых вызывает
ретроградную дегенерацию клеток
медиального коленчатого тела (рис.
666). В соответствии с физиологическими
данными во всех областях слуховой коры
возникают первичные ответы при действии
звуко-
Рис.
66. Слуховая кора и ее связи с внутренним
коленчатым телом
(по: Woolsey, 1961).
А
—
слуховые корковые поля кошки; Б
—
схема связей внутреннего коленчатого
тела
с различными областями слуховой
коры. AI
—
центральное слуховое поле; All,
AIII,
Ер —
слуховые поля; ins
—
инсулярное поле, Т
—
темпоральное поле, Assoc — ассо-
циативная
кора.
вых
сигналов или при прямом электрическом
раздражении волокон слухового нерва.
Проекции из медиального коленчатого
тела неравноценны в этих областях:
область AI (или зона AI) получает наиболее
мощную проекцию из переднего отдела
медиального коленчатого тела. Область
инсуло-темпоральная получает меньшее
количество восходящих путей от заднего
отдела медиального коленчатого тела.
Области (или зоны) Ер и АП получают
только коллатеральную проекцию от
медиального коленчатого тела, так как
удаление этих областей не вызывает
заметной дегенерации в медиальном
коленчатом теле, но удаление областей
АП и Ер вместе с областью AI вызывает
более обширную дегенерацию, чем в одной
области AI. Цитоархитектоническая
характеристика слуховой области у
кошки позволяет выделить центральную
область, совпадающую с областью AI, и
периферический пояс, совпадающий с
положением областей АП и Ер (Rose, 1949).
Строение слухо- 206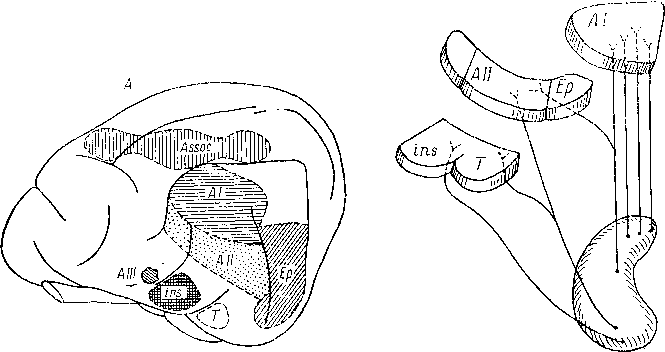
вой
коры у собак мало отличается от описанного
выше строения слуховой коры кошки,
тогда как слуховая кора у приматов
организована иначе, чем у хищных (Ades,
Felder, 1942; Pandya et al., 1969).
Для
слуховой шестислойной коры характерна
мелкоклеточ- ность всех слоев, отсутствие
крупных пирамид в 5-м и 3-м слоях, наличие
большого числа звездчатых короткоаксонных
клеток, которые в основном сконцентрированы
в 4-м слое, где они располагаются
колонками. Зведчатых клеток также много
во 2-м и 3-м слоях, в результате чего
происходит слияние 2-го — 4-го слоев.
Можно различить два вида зведчатых
клеток: крупные клетки с широко ветвящимся
аксоном, который оканчивается на
клетках, расположенных на значительном
расстоянии, и мелкие клетки с аксонами,
которые густо сконцентрированы возле
тела клетки. Мелкие пирамидные клетки,
широко представленные во всех слоях,
характеризуются обильными разветвлениями
дендритов как верхушечных, так и
базальных. Аксоны этих клеток дают
большое количество многократно
ветвящихся коллатералей. Для слуховой
коры характерны также мелкие пирамидные
клетки, расположенные на уровнях 4-го
и 5-го слоев, у которых аксон дугообразно
огибает тело клетки и направляется
вверх. Афферентные волокна проходят в
коре либо в виде перпендикулярных
волокон, сильно ветвящихся и оканчивающихся
в основном на клетках 3-го и 4-го слоев,
либо в виде наклонных маловетвящихся
волокон (Ramon у Cajal, 1911; Lorente de Nd, 1949; Попова,
1962). В периферических полях слуховой
коры клетки крупнее, 4-й слой слабее
выражен, чем в центральном поле,
звездчатых клеток меньше и они менее
разнообразны.
На
клетках слуховой коры встречаются два
вида синапсов; более крупные в виде
луковиц (bouton) и мелкие петлевидные.
Крупные синапсы располагаются только
на теле клетки, по 1—2 синапса на клетке,
петлевидные располагаются на дендритах
небольшими группами (Lorenzo, I960),
Кроме
этого, классического слухового пути,
существуют дополнительные слуховые
пути, идущие параллельно. Они проходят
через ретикулярную формацию продолговатого
и среднего мозга (Morest, 1964; Rasmussen, 1964) в
заднюю таламическую группу ядер. К этой
группе относятся: заднее ядро (nucl.
posterior thalami), подколенчатое (nucl.
suprageniculatus), заднее латеральное (nucl.
lateralis posterior thalami), подушка (pulvinar) и
крупноклеточная часть внутреннего
коленчатого тела (Rose, Woolsey, 1958; Moore,
Goldberg, 1961, 1963). Волокна от этих ядер идут
в корковые поля, окружающие слуховые
основные поля.
207
НИСХОДЯЩИЕ
СЛУХОВЫЕ ПУТИ
От
различных слуховых полей к нижележащим
образованиям слуховой системы
направляются довольно многочисленные
нисходящие волокна, которые, переключаясь
на разных уровнях, образуют нисходящий
слуховой путь (рис. 67). В слуховой коре
обнаружена определенная зона, от которой
идет наибольшее
число
волокон. Эта зона определена как начало
нисходящего слухового пути —
инсуло-темпоральная зона (Desmedt, Meche- Ise,
1959). Нисходящие волокна от коры
направляются ко внутреннему коленчатому
телу, главным образом к его крупноклеточной
части, которая является важным звеном
в нисходящей слуховой системе. Все
области внутреннего коленчатого тела,
за исключением каудальной части
дорсальной области, в определенной
пропорции получают нисходящие волокна
от всех слуховцх
208
областей
(Rose, Woolsey, 1958; Walther, Rasmussen, 1960). Нисходящие
волокна от внутреннего коленчатого
тела оканчиваются билатерально в ядрах
задних бугров четверохолмия, небольшое
число волокон проходит в дорсальное и
интерстициальное ядра латеральной
петли, от которых некоторая часть
волокон проходит без переключения
в задние холмы. (Гранстрем, 1963). От задних
бугров четверохолмия идут два нисходящих
пучка: латеральный и медиальный.
Латеральный пучок возникает в
клеточно-волокнистой капсуле заднего
бугра и его латеральной коре и затем
идет в латеральной части ствола мозга.
Часть волокон этого пучка оканчивается
на клетках периоливарных ядер своей и
противоположной сторон, а часть входит
в ипсилатеральное дорсальное кохлеарное
ядро (Rasmussen, 1964). От ядра заднего бугра
четверохолмия и от дорсального ядра
латеральной петли начинается медиальный
пучок. Его волокна входят в верхнеоли-
варную область, где часть их переключается.
В этой области от клеток ретрооливарной
группы начинается оливо-кохлеарный
пучок, или пучок Расмуссена (Rasmussen,
1946). Этот пучок проходит на
противоположную сторону и вместе с
волокнами вестибулярного нерва
выходит из ствола мозга, далее через
вестибулокохлеарный анастомоз
входит в кохлеарный нерв и достигает
улитки. Крупные синапсы на внутренних
и наружных волосковых клетках являются
окончаниями волокон оливо-кохлеарного
пучка. Из верхнеоливарной области идут
также пучки нисходящих волокон через
интермедиальную полоску и трапециевидное
тело в кохлеарные ядра своей и
противоположной сторон, при этом
особенно обильно оказывается снабженным
нисходящими волокнами дорсальное
кохлеарное ядро (Lorente de Nd, 1933b; Rasmussen,
1960, 1964).
Сенсорные
системы