

ОБОНЯНИЕ.
ВКУС
Г
лае а 14
ОБОНЯНИЕ
СТРУКТУРНАЯ
ОРГАНИЗАЦИЯ ПЕРИФЕРИЧЕСКИХ ОТДЕЛОВ
ОБОНЯТЕЛЬНОГО АНАЛИЗАТОРА И ОБОНЯТЕЛЬНОЙ
ЛУКОВИЦЫ
Периферические
отделы обонятельного анализатора
обычно называют органами обоняния. У
человека они представлены
эпителиоподобной выстилкой, или
слизистой обонятельной области носа
(regio olfactoria), которая занимает среднюю
часть верхней носовой раковины и
соответствующий ей участок слизистой
оболочки носовой перегородки (рис. 219,
а),
У многих млекопитающих с хорошо развитым
обонянием обонятельная область носа
резко увеличивается за счет развития
системы этмоидальных раковин —
ethmoturbinalia, представляющих собой выросты
решетчатой кости (рис. 219, б).
У круглоротых и рыб обонятельная
выстилка покрывает внутреннюю поверхность
складчатых обонятельных ямок или
подкожных мешков, которые располагаются
на голове в ближайшем соседстве с
ротовым отверстием и омываются
водой. У амфибий эти мешки одеваются
хрящевой капсулой и сообщаются не
только с окружающей средой, но и с
помощью первичных хоан с ротовой
полостью. У многих пресмыкающихся
(змеи, ящерицы, крокодилы) появляются
зачатки обонятельных раковин,
получивших дальнейшее развитие у птиц
и млекопитающих и делящих носовую
полость на переднюю — респираторную
и заднюю — обонятельную области.
К
органам обоняния обычно относят также
трубчатый или мешковидный якобсонов
орган наземных позвоночных, расположенный
в нижнемедиальной стенке их носовой
полости и сообщающийся с полостью
носа или рта. Он особенно хорошо развит
у некоторых пресмыкающихся и многих
млекопитающих. У птиц якобсонов орган
отсутствует, а у человека, как правило,
редуцируется еще в эмбриональном
периоде. Точная роль якобсонова органа
в обонятельной рецепции не установлена
(Moulton, Beidler, 1967).
33* 515Раздел III
а
б
Рис.
219. Схема строения носовой полости.
а
—
у человека (по: А. И. Бронштейн, 1950), б
— у
хомяка (по: Bang, Bang, 1959). На а: 1 — верхняя
носовая раковина, 2
—
средняя носовая раковина, 3
—
нижняя носовая раковина, 4 — верхний
носовой ход, б
—
средний носовой ход, 6
—
нижний носовой ход. Продольный срез.
На б
справа затемненным контуром
отмечены зоны распределения
обонятельного эпителия, волнистой
линией
показан респираторный эпителий.
Поперечный срез.

Гистологические
исследования органа обоняния с помощью
светового микроскопа были начаты во
второй половине прошлого столетия
(см.: Винников, Титова, 1957). Эти исследования
пока зали, что обонятельный эпителий
(= эпителиоподобный рецепторный слой
обонятельной выстилки), достигающий в
толщину 100—150 мкм, включает рецепторные
клетки, которые относятся к наиболее
древним — первичночувствующим
рецепторам. Они снабжены собственным
центральным отростком (рис. 220, а),
выполняющим роль аксона и передающим
полученную клеткой информацию в
обонятельные луковицы, которые являются
первичными центрами обонятельного
анализатора. Рецепторные клетки окружены
и изолированы друг от друга телами
опорных клеток, часть из которых,
располагающаяся вблизи базальной
мембраны рецепторного слоя, носит
название базальных. В соединительнотканном
слое обонятельной выстилки (tunica propria)
у всех наземных позвоночных, начиная
с амфибий, локализуются концевые отделы
боуменовых желез. В соединительнотканном
слое располагаются также многочисленные
кровеносные сосуды и пучки безмякотных
обонятельных нервов, образованные
центральными отростками обонятельных
клеток. У млекопитающих, в том числе и
у человека, эти отростки не формируют
обособленного нервного ствола, а в
виде многочисленных (до 20) коротких
и тонких обонятельных нитей — fila
olfactoria — проходят через отверстия
решетчатой кости к обонятельным
луковицам мозга.
Единый
обонятельный нерв отсутствует и у
некоторых видов рыб, у которых обонятельные
луковицы непосредственно прилегают к
обонятельным мешкам и связываются с
передним мозгом с помощью длинных
обонятельных трактов.
Строение
и цитохимия обонятельных рецепторов.
Количество
обонятельных рецепторов весьма велико,
но у различных животных не одинаково.
Оно в значительной мере определяется
площадью, занимаемой обонятельным
эпителием, и зависит также от размеров
этих рецепторов и особенно от плотности
их расположения в обонятельном эпителии.
Разными авторами (см. обзоры: Allison, 1953а;
Moulton, Beidler, 1967) подсчитано, что число
обонятельных клеток у миноги равно 800
тыс., у человека — 10 млн, у кролика —
100 млн и у немецкой овчарки — 224 млн. При
этом площадь обонятельного эпителия
составляет у человека — 10 см2,
у кролика — 9.3 и у немецкой овчарки —
196.46 см2.
Таким образом, у кролика на 1 мм2
обонятельного эпителия должно приходиться
120 тыс., а у человека 10 тыс. обонятельных
рецепторов. Обонятельные клетки имеют
веретеновидную форму и снабжены,
кроме центрального, коротким периферическим
отростком, заканчивающимся на поверхности
рецепторного слоя сферическим утолщением
— обонятельной булавой (рис. 220, б).
517
Электронномикроскопические
исследования обонятельного эпителия
у круглоротых (Бронштейн, Иванов, 1965;
Thornhill, 1967), костистых рыб (Trujllo-Cenoz, 1961;
Bannister, 1965; Бронштейн, Пяткина, 1966;
Wilson, Westerman, 1967; Филюшина, Бахтин, 1969, и
др.), земноводных (Bloom, 1954; Reese, 1965;
а
Рис.
220. Схема строения обонятельного
эпителия.
а
—
по данным световой микроскопии
(Винни-
ков, Титова, 1957).
Костанян,
1971), пресмыкающихся (Бронштейн, Пяткина,
1968), птиц (Graziadei, Bannister, 1967; Бронштейн,
Пяткина, 1969а) и млекопитающих (Gasser, 1956;
Lorenzo, 1957, 1963; Graziadei, 1965; Andres, 1966, 1969; Frish,
1967; Okano et al., 1967) показали, что вершины
обонятельных клеток устроены у них
сходным образом (рис. 221, а—в).
Несмотря на различный уровень
филогенетического развития, разную
среду обитания и неодинаковую роль
обоняния в жизни различных позвоночных
животных непременной структурой
воспринимающих отделов (вершин)
обонятель- 518
6
Рис.
220 (продолжение).
б
—
по данным электронной микроскопии
(оригинал). На б: БК — базальная клетка,
БМ
—
базальная мембрана, ВЛ1
—
обычные обонятельные волоски, BJIII
—
длинные обонятельные волоски, BJIIII
—
комплексные обонятельные волоски, ВЛIV
—
обонятельные волоски с дополнительными
периферическими фибриллами, МВО
—
микроворсинки опорных клеток, МБР
—
микроворсинки рецепторных клеток, ОБ
—
обонятельная булава, ОК — опорная
клетка, ЦО
— центральные
отростки обонятельных клеток.
ных
клеток являются киноцилиоподобные
волоски. Обонятельные волоски
начинаются в цитоплазме булавы от
характерных базальных телец (рис. 221,
а—в)
и содержат, по крайней мере в своем
проксимальном сегменте, набор трубчатых
фибрилл, состоящий из 2 центральных и
расположенных вокруг них в виде кольца
9 пар периферических фибрилл. Именно
такой набор фибрилл свойствен
локомоторным ресничкам и жгутикам
одноклеточных и многоклеточных
организмов и, как выяснилось в последнее
время, видоизмененным волоскам, ресничкам
или жгутикам, представленным в
воспринимающих отделах рецепторных
клеток большинства органов чувств
(Винников, 1965). В дистальном сегменте
обонятельные волоски утончаются и
постепенно утрачивают внутренние
фибриллы.
Количество
волосков в обонятельных клетках
колеблется от 1 у некоторых видов рыб
до 15—20 и более у собаки и белой крысы.
Длина большинства обонятельных волосков
составляет 15—30 мкм при диаметре, равном
вблизи основания 0.25—0.3 мкм. Отдельные
обонятельные клетки снабжены также
более длинными волосками (рис. 220,6),
размеры которых у некоторых видов
лягушек (Rana catesbiana, Rana ridibunda и др.) могут
достигать 150— 200 мкм (Hopkins, 1926; Reese, 1965;
Костанян, 1969). Интересно, что длина
ресничек мерцательного эпителия
позвоночных обычно не превышает 8—10
мкм.
Третьей
разновидностью являются более толстые
обонятельные волоски, или жгуты,
которые, как оказалось (Бронштейн,
Пяткина, 1966, 1968), состоят из целого
комплекса киноцилий, покрытых общей
плазматической оболочкой (рис. 220, 6).
Обонятельные
волоски могут чередоваться с короткими
(0.5— 1.0 мкм) пальцевидными выростами
цитоплазмы — микровиллами, вообще
не содержащими трубчатых фибрилл. У
некоторых рыб были обнаружены даже
отдельные клетки, сходные по форме и
ультраструктурной организации с
обонятельными рецепторами, но несущие
на поверхности булавы только одни
микровиллы (Bannister, 1965; Филюшина, Бахтин,
1969). Подобные же клетки были описаны в
якобсоновом органе пресмыкающихся
(Bannister, 1968). В органе обоняния такие
клетки, по-видимому, относятся к
неполностью дифференцированным
рецепторам (Trujillo-Genoz, 1961; Bannister, 1968).
Обонятельные
волоски погружены в жидкую среду,
которая у наземных позвоночных
продуцируется главным образом
боуменовыми железами. В волосках,
по всей видимости, осуществляются
первичные процессы обонятельной
рецепции (Винников, 1971).
Возможно,
что волоски обеспечивают увеличение
рецептивной поверхности обонятельной
клетки. Подсчитано, например, что у
кролика общая поверхность всех
обонятельных волосков составляет около
600 см2
при площади всей поверхности обонятельного
эпителия (включая не только вершины
рецепторных, но и
520
Рис.
221. Электронные микрофотографии вершины
обонятельной клетки.
а
*— речная минога, увел. 30 000 (Бронштейн,
Иванов, 1965), б — травяная лягушка, увел.
35 000 (Бронштейн, Пяткина, 19696), в
—
зеленая мартышка, увел. 40 000 (оригинал).
На а—в:
БТ —
базальное тельце обонятельного волоска,
ВЛ
—
обонятельный волосок, К — корешок, М
—
митохондрия, МТ
—
микротрубочки, PH
—
рецепторная клетка, СГ
—
секреторные гранулы опорнрй клетки.
Остальные обозначения см. рис. 220.
Рис.
221 (продолжение).
Рис.
221 (продолжение).
превосходящие
их по величине вершины опорных клеток)
около 10 см2.
Не исключено, что волоски выполняют и
более сложную функцию в рецепторном
акте. Согласно результатам прижизненного
изучения изолированных кусочков
обонятельного эпителия круглоротых,
рыб, земноводных и пресмыкающихся
(Hopkins, 1926; Бронштейн, 1964; Reese, 1965), обонятельные
волоски активно двигаются, проделывая
сложные асинхронные колебания, включающие
компоненты маятникообразного,
изгибательного и воронкообразного
движения. Однако значение этого движения
и его необходимость для работы
обонятельной клетки не установлены.
Известно, например, что у некоторых
видов лягушек — Rana catesbiana, Rana ridibunda, Rana
pipiens — наиболее длинные обонятельные
волоски в обычных условиях наблюдения
неподвижны (Hopkins, 1926; Reese, 1965; Костанян,
1969). До сих пор неясно также, двигаются
ли обонятельные волоски у теплокровных
животных.
Гистохимические
исследования (Baradi, Bourne, 1951, 1953; Amicis,
Zorzoli, 1957; Бронштейн, 1965, и др.) показали,
что в состав обонятельных волосков
входят липиды, белки, богатые
сульфгидрильными группами, и фосфатазы.
С помощью электронной гистохимии в
их оболочках выявлена высокая АТФ-азная
активность (Бронштейн, Пяткина, 1969 б).
В то же время обонятельные волоски
практически не содержат нуклеиновых
кислот, окислительных ферментов и
ацетилхолинэстеразы. Вершина обонятельных
волосков, как правило, заполнена
гомогенной или зернистой осмиофильной
субстанцией, химический состав которой
неизвестен.
Булавы
обонятельных клеток имеют диаметр
порядка 1—2 мкм. Кроме базальных телец
и связанных с ними микротрубочек или
поперечно исчерченных корешков,
обонятельные булавы содержат
некоторое количество мелких вакуолей,
а также митохондрии, число которых
заметно возрастает в месте перехода
булавы в периферический отросток.
Обонятельная булава является одним из
важных цитохимических центров рецепторной
клетки. В ней высока концентрация
белковых сульфгидрильных и карбоксильных
групп и липидов, а также весьма выражена
активность некоторых окислительных
ферментов и фосфомоноэстераз. В булаве,
по-видимому, вырабатывается АТФ, которая
необходима, в частности, для обеспечения
двигательной активности обонятельных
волосков (Бронштейн, 1964, 1966). Есть
основания полагать (Бызов, Флерова,
1964), что обонятельные булавы являются
местом генерирования рецепторных
потенциалов.
Периферический
отросток, соединяющий обонятельную
булаву с телом рецепторной клетки,
имеет длину порядка 20—90 мкм. В нем
содержатся митохондрии и проходят
многочисленные микротрубочки,
спускающиеся сюда из булавы. Форма и
толщина периферических отростков
варьируют, в связи с чем в световом
521
микроскопе
различают палочковидные (нитевидные)
и колбочко видные обонятельные клетки
(Догель, 1886, Винников, Титова, 1957). Неясно,
однако, имеют ли эти разновидности
обонятельных клеток какие-либо
функциональные отличия или форма их
периферических отростков обусловлена
только необходимостью наиболее
компактной упаковки рецепторов в
эпителиальном слое.
Тело
обонятельных клеток не содержит
каких-либо специфических для этих
рецепторов ультраструктур. Кроме ядра,
здесь локализуются многочисленные
митохондрии, лизосомы, элементы аппарата
Гольджи и достаточно хорошо развитые
цистерны эндоплазматической сети
с рибосомами (рис. 220, б).
Согласно гистохимическим данным
(Baradi, Bourne, 1953; Бронштейн, 1965; Цопова, 1966;
Щекочихина, 1967), в ядросодержащих отделах
обонятельных рецепторов концентрируется
цитоплазматическая РНК, различные
белки, в том числе ферменты, а также
гликоген. При воздействии на обонятельную
выстилку пахучими веществами распределение
РНК в них меняется (Бронштейн, 1965).
От
нижнего полюса обонятельной клетки
отходит тонкий центральный отросток
(рис. 220, б), имеющий диаметр около 0.2—0.5
мкм. Он содержит нейрофибриллы и
митохондрии и по ультраструктурной
организации вполне соответствует
аксонам нервных клеток. Центральные
отростки ряда обонятельных рецепторов
группируются в пучки, прободают базальную
мембрану обонятельного эпителия и
входят в состав безмякотных обонятельных
нервов. От окружающих элементов эти
пучки изолируются вначале выростами
базальных клеток, а затем мезаксонами
шванновских клеток, которые охватывают
цитоплазматическим футляром большое
число (до нескольких сот) волокон.
К
опорным элементам обонятельного
эпителия относятся длинные, цилиндрические,
собственно опорные клетки и расположенные
над базальной мембраной эпителия
базальные клетки, имеющие неправильную
кубовидную форму и не достигающие
поверхности рецепторного слоя (рис.
220, б). Как правило, свободная поверхность
опорных клеток покрыта микровиллами
(рис. 220, б), длина которых не превышает
3—5 мкм. Такие структуры обычно
свойственны клеткам, интенсивно
выделяющим или поглощающим какие-либо
вещества. Цитоплазма опорных клеток
достаточно богата ультраструктурами.
В них обнаруживаются каналы и цистерны
гладкой и шероховатой эндоплазматической
сети, рибосомы, структуры аппарата
Гольджи, митохондрии, лизосомы,
тонкие опорные фибриллы, а в апикальных
отделах множество секреторных гранул
(рис. 220, б; 221, б, в), выделяющихся в просвет
обонятельной полости. Гистохимическими
методами в опорных клетках показано
наличие ряда биологически важных
веществ, в том числе нуклеиновых кислот,
ферментов и полисахаридов. Есть основания
считать, что опорные клетки обонятельного
эпителия выполняют не только механичес
522
кую
функцию, но и активно участвуют в его
метаболизме. Они, в частности, секретируют
кислые мукополисахариды, добавляя их
к секрету боуменовых желез (Попова,
1966). Интересно, что подобная секреторная
функция свойственна также опорным
клеткам круглоротых и рыб. У этих
животных выделенный опорными и
бокаловидными клетками секрет
предохраняет, по-видимому, обонятельные
волоски от непосредственного
соприкосновения с водой. Таким образом,
внешние условия взаимодействия пахучих
веществ с обонятельными рецепторами
у водных и наземных позвоночных
оказываются аналогичными.
Обонятельные
клетки обеспечиваются питательными
веществами и, вероятно, кислородом
через сеть капилляров, расположенных
под рецепторным эпителием. Как уже
отмечалось, у наземных позвоночных
здесь же локализуются боуменовы железы.
Секрет их содержит главным образом
кислые и нейтральные мукополисахариды,
а также белки и липиды (de Vanna, Salonna, 1953;
Бронштейн, 1965; Попова, 1966). Гистохимически
в нём обнаружено значительное количество
натрия (Бронштейн, Пяткина, 1969 б).
В
соединительнотканном слое обонятельной
выстилки кроме пучков безмякотных
обонятельных нервов проходят отдельные
миэлинизированные волокна тройничного
нерва. Однако в обонятельном эпителии
не имеется никаких синаптических
образований или свободных нервных
окончаний.
При
биохимических исследованиях (Gerebtzoff,
Philippot, 1957; Briggs, Duncan, 1961, 1962; Kurihara, 1967, и др.)
в обонятельной выстилке некоторых
животных были обнаружены каротиноиды.
Однако до сих пор неясно, где локализованы
эти пигменты (в обонятельных рецепторах,
опорных клетках, боуменовых железах),
и имеют ли они непосредственное отношение
к обонятельной рецепции (Wright, 1957, 1964;
Jackson, 1960; Moulton, 1962). В последние годы
предпринимались попытки изолировать
обонятельные рецепторы или их фрагменты
дифференциальным центрифугированием
гомогенатов обонятельной выстилки
(Ash, Bransford, 1966). Были получены фракции
обонятельных булав и их плазматических
мембран (Koch, 1969), богатые
НАД-Н21-цитохром-с-редуктазой,
К+—
Na+—АТФ-аЗой
и нечувствительной к оубаину Mg2+—АТФ-азой.
Получение таких фракций позволило
приступить к поискам специфических
белков (Ash, 1968), которые могут быть
ответственны за первичный акт
взаимодействия обонятельных рецепторов
с молекулами пахучих веществ.
Морфология
обонятельной луковицы. Обонятельные
луковицы лежат у человека на базальной
поверхности лобных долей у переднего
конца обонятельной борозды (sulcus
olfactorius). Они имеют
1
НАД—Н2
— восстановленный никотин адениндинуклеотид.
523
Рис.
222. Строение обонятельной луковицы
млекопи-
тающих.
А
—
схематический рисунок (по: Moulton, Tucker,
1964); Б
—
при обработке горизонтального среза
по Гольджи (по: Valverde, 1965). На А:
1 —
обонятельные клубочки, 2
—
пучковые клетки, з
—
митральные клетки, 4
—
зерновидные клетки, 5
—
наружный сетевидный слой, 6
—
внутренний сетевидный слой, 7
—
пластинка решетчатой кости, 8
—
обонятельный эпителий, 9
—
обонятельные клетки. На Б: I—VI
—
различные слои луковицы (см. текст).
1—16
—
клубочки, 17
—
волокна обонятельного нерва, 18—21
—
волокна, оплетающие митральные
клетки, 22,
25—зо —
волокна, контактирующие с отростками
зерновидных клеток, a—g
—
митральные нейроны, h
—
горизонтальные клетки, г, j
—
пучковые клетки, fe, I
—
межклубочковые клетки, т, тг, о —
зерновидные клетки, 1а—lg
—
аксоны митральных клеток, 2а—За
—
возвратные коллатерали аксонов
митральных клеток, 1г—lj
—
аксоны пучковых клеток, Ik
—
аксоны межклубочковых клеток. На
Б вверху слева —
сплетение волокон во внутреннем
сетевидном слое.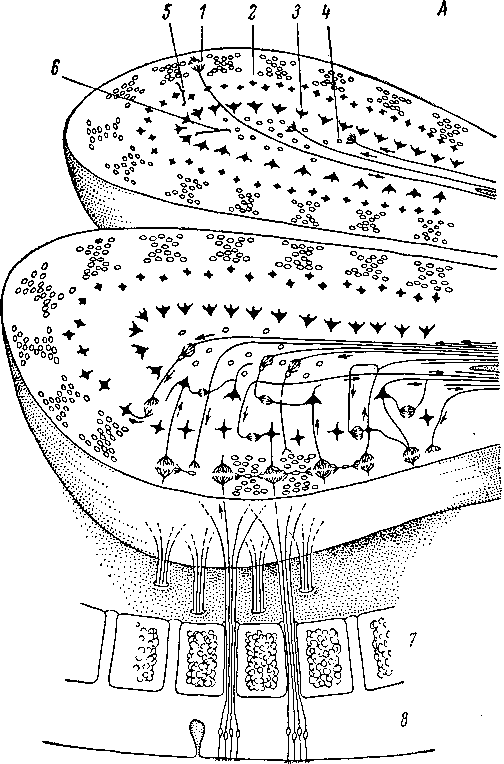
6
Рис.
222 (продолжение).
овальную
форму, длину 8—10. мм, ширину 3—4 мм и
толщину 2—3
мм (Кононова, 1957). Однако относительные
размеры обонятельных луковиц у
разных животных сильно варьируют,
главным образом в связи со степенью
развития у них обонятельной функции
(Negus, 1954; Nieuwenhuys, 1967, и др.). Так, у некоторых
сумчатых луковица составляет до
половины всей длины полушария (Ariens
Kappers et al., 1960), у приматов и птиц они
развиты слабо. У зубатых китов типичные
обонятельные луковицы вообще отсутствуют.
У позвоночных имеется две луковицы.
Однако у некоторых рыб, у бесхвостых
амфибий и некоторых птиц обонятельные
луковицы частично или полностью
сливаются по средней линии.
Обонятельные
луковицы построены по типу корковых
центров со сложным расположением
элементов и поэтому могут быть отнесены
к корковым структурам. Иногда их включают
в состав древней коры (Gastaut, Lammers, 1961).
Более принято, однако, рассматривать
обонятельные луковицы отдельно от
палеокортикальных областей, как особую
часть обонятельного мозга (Филимонов,
1949).
Со
времен классических исследований
Кахаля (Ramon у Cajal, 1911) в обонятельной
луковице млекопитающих обычно различают
шесть основных слоев (рис. 222, А,
Б),
которые расположены концентрически в
следующем порядке (считая от поверхности):
7— слой волокон обонятельного нерва;
II
—
слой клубочков; III
—
наружный сетевидный слой; IV
—
слой тел митральных клеток; V
—
внутренний сетевидный слой; VI
—
зернистый слой (последние два слоя
иногда объединяются). У некоторых
животных в центре обонятельной луковицы
располагается, кроме того, канал, или
желудочек, выстланный эпендимой.
Наружный
слой луковицы представляет собой густое
сплетение волокон обонятельного нерва.
В этом сплетении происходит интенсивное
перераспределение центральных отростков
обонятельных клеток (Allison, 1953, б),
которые на пути от обонятельного
эпителия до луковицы, по-видимому, идут
почти параллельно. Первое синаптическое
переключение для сигналов, приходящих
от обонятельных рецепторов, осуществляется
в клубочках луковицы (рис. 222, А, Б,
7—76), составляющих ее второй слой. Каждый
клубочек включает окончания большого
числа центральных отростков
рецепторных клеток (рис. 222, А, Б, 77),
образующих синаптические контакты
с дендритами значительно меньшего
числа клеток луковицы (Lorenzo, 1963; Andres,
1965).
.
Обонятельный клубочек имеет округлую
форму. Средний диаметр его составляет
у кролика 185 мкм, у крысы — 115 мкм (Allison,
1953а). У млекопитающих каждый клубочек
заключен в хорошо развитую глиальную
капсулу (Винников, Титова, 1957). Наиболее
крупными клетками, посылающими дендриты
в клубочки, являются митральные клетки
луковицы (рис. 222, Б,
526
a—g).
Они имеют характерную форму тела и у
млекопитающих располагаются на одном
уровне и выделяются в самостоятельный
IV
слой луковицы. Аксоны митральных клеток
(рис. 222, 5, 1а—2
g) образуют основной, а возможно и
единственный проекционный путь из
обонятельной луковицы к центрам
следующего порядка (см. ниже). У
млекопитающих, как правило, каждая
митральная клетка имеет только один
главный (апикальный) дендрит, контактирующий
только с одним клубочком, хотя встречаются
клетки и с несколькими апикальными
дендритами (Ramon у Cajal, 1911; Allison 1953а;
Valverde, 1965). У других позвоночных митральная
клетка обычно связана с несколькими
клубочками. Боковые дендриты митральных
клеток ветвятся в наружном сете7
видном
слое (III
слой луковицы). Описаны тонкие волокна,
оплетающие тела митральных клеток
в виде характерных корзинок (рис. 222, 5,
18—21),
(Valverde, 1965). Происхождение этих волокон
неясно.
Кроме
митральных клеток с клубочками связаны
так называемые пучковые клетки (рис.
222, Б,
I—])
и довольно многочисленные мелкие
клетки (наружные зерна, или межклубочковые
клетки) (рис. 222, Б,
к, I),
локализующиеся во II
слое луковицы. Дендриты межклубочковых
клеток оканчиваются в одном или
нескольких рядом расположенных
клубочках, аксоны же их (рис. 222, Б,
1к)
разветвляются в слое клубочков и в
наружном сетевидном слое (Ramon у Cajal,
1911; Allison, 1953а), контактируя, по некоторым
данным, с дендритами и телами пучковых
клеток (Andres, 1965). Тела пучковых клеток
(рис. 222, Б,
I—/)
разбросаны в наружном сетевидном слое.
По своим очертаниям и способу контакта
с клубочками они напоминают митральные
клетки, но меньше их по размерам и более
многочисленны. Помимо главного дендрита
они имеют по несколько боковых
дендритов, которые также ветвятся в
наружном сетевидном слое. Слабо
миэлинизированные аксоны пучковых
клеток (рис. 222, Б,
I.—I^
отдают много коллатералей, спускаясь
преимущественно во внутренний сетевидный
слой (слой V), где образуют, по мнению
Кахаля, весьма запутанное переплетение.
Некоторые из этих аксонов идут
горизонтально в поверхностных слоях,
разветвляясь ореди клубочков (Valverde,
1965).
Центральная
часть луковицы заполнена многочисленными
зерновидными клетками, или
клетками-зернами (рис. 222, Б,
т, п,о), по
виду которых назван весь VI
слой. Для большинства клеток- зерен
характерны короткие ветвящиеся дендриты,
направленные внутрь, и длинный радиальный
отросток, ответвления которого
располагаются в наружном сетевидном
слое.
Отростки
зерновидных клеток усеяны шипиками и
контактируют с большим числом волокон
луковицы (рис. 222, Б,
25—30), имеющих
как местное происхождение, так и с
центробежными, о которых речь пойдет
ниже. В зернистом слое описано также
несколько типов звездчатых клеток с
короткими аксонами.
527
По
данным электронной микроскопии в
наружном сетевидном слое обнаружены,
кроме обычных синаптических контактов,
своеобразные реципрокные синаптические
комплексы (Hirata, 1964; Andres, 1965; Price, 1968а,
1968b), связывающие, как полагают, дендриты
митральных и пучковых клеток с отростками
клеток- зерен. Предполагают, что в таком
комплексе один синапс (направленный
от митральной или пучковой клетки к
отростку зерновидной) является
возбуждающим, а другой с противоположной
направленностью — тормозящим.
Поскольку отросток зерновидной клетки
связан с дендритами многих митральных
клеток, такая структурная организация
создает возможность осуществления
латерального торможения без обязательного
участия активности тел зерновидных
клеток (Rail et al., 1966; Ottoson, Shepherd, 1967). Аксоны
митральных клеток, проходя через слой
клеток-зерен, отдают многочисленные
возвратные коллатерали (рис. 222, Б,
2а. За), которые
образуют синаптические контакты в
различных слоях луковицы. Слой
клеток-зерен непосредственно переходит
в клеточные массы так называемого
переднего обонятельного ядра (nucleus
olfactorius), которое относят к обонятельным
центрам третьего порядка. У многих
животных передняя часть этого ядра
довольно глубоко вдается в центральную
часть луковицы (Ariens Kappers et al., 1960), окружая
ее желудочек.
Помимо
перечисленных 4 основных групп клеток
(митральные, межклубочковые, пучковые,
зерновидные), в луковице имеется еще
несколько типов нейронов (например,
горизонтальные клетки) (рис. 222, Б,
h).
Эти клетки сравнительно малочисленны.
Описаны также различные глиальные
элементы луковицы (Ramon у Cajal, 1911).
Следует,
наконец, упомянуть о центробежных
волокнах луковицы. Они поступают из
переднего обонятельного ядра,
обонятельного бугорка, препириформной
коры и других отделов обонятельного
мозга и могут быть гомолатеральными
или гетеролатераль- ными. Пути центробежных
волокон в луковице изучены еще
недостаточно, хотя известно, что они
могут оканчиваться в слое клубочков,
в зернистом и во внутреннем сетевидном
слоях (Ariens Kappers et al., 1960; Valverde, 1965, и др.).
Значение центробежных связей в луковице
весьма велико, особенно в связи с
отмечавшимся уже отсутствием эфферентной
иннервации обонятельного эпителия.
Приводившиеся
данные свидетельствуют об очень сложной
структурной организации обонятельной
луковицы. Как показали опыты с локальными
повреждениями (Le Gros Clark, Warwick, 1946; Le Gros
Clark, 1957; Винников, Титова, 1957), не существует
топической пространственной проекции
обонятельных рецепторов на элементы
луковицы. Можно говорить лишь о довольно
грубом соответствии между макрообластями
обонятельного эпителия и отдельными
сегментами луковиц. Вместе с тем,
строение
528