

Структура главных липидов биологических мембран имеет одну общую черту, которая определяет их способность к образованию двойного липидного слоя в водной среде. Все эти соединения амфифильны по своей природе, то есть содержат как гидрофильные, так и гидрофобные участки.
Название липида |
Гидрофобные участки |
Гидрофильные |
|
|
участки |
Фосфоглицероли- |
цепи жирных кислот |
Фосфорилированный |
пиды |
|
спирт |
Сфингомиелин |
цепь жирной кислоты и угле- |
Фосфорилированный |
|
водородная цепь сфингозина |
холин |
Гликолипиды |
цепь жирной кислоты и угле- |
Углеводы |
|
водородная цепь сфингозина |
|
Холестерин |
углеродный скелет |
ОН-группа при С3- |
|
|
атоме углерода |
В бислое молекулы липидов уложены в виде двух параллельных монослоев, обращенных друг к другу своими гидрофобными сторонами. Полярные группы липидных молекул образуют соответственно две гидрофильные поверхности, отделяющие внутреннюю неполярную область бислоя от водной среды. Толщина липидного бислоя определяется, прежде всего, длиной углеводородных цепей радикалов жирных кислот, а также плотностью упаковки липидных молекул и варьирует в пределах 5-10 нм.
Условия, в которых находятся молекулы липидов, по разные стороны бислоя могут значительно различаться по таким параметрам, как ионный состав среды, рН, наличие или отсутствие мембрано-активных веществ и др. В результате состав липидов на наружной стороне бислоя может быть иной, чем на внутренней. Так, в цитоплазматической мембране эритроцитов фосфатидилэтаноламин, фосфатидилсерин и фосфатидилинозит локализованы преимущественно на внутренней стороне мембраны, а фосфатидилхолин и сфингомиелин – на внешней. Холестерин также неравномерно распределен между внутренним и внешним листками липидного бислоя. Его содержание во внешнем монослое мембраны почти в два раза больше, чем во внутреннем.
Бислой может находиться в двух основных состояниях – кристаллическом (гелевом, «твердом») и жидкокристаллическом («текучем», «расплавленном»). Состояние геля образуется, когда жирнокислотные «хвосты» липидов находятся в полностью упорядоченной трансконформации. Переход из одного состояния в другое происходит при определенной температуре, которая зависит как от строения углеводородных цепей жирных кислот в составе липидных молекул, так и от природы их полярных головок. Во многих случаях фазовые изменения
71

могут происходить и при постоянной температуре за счет изменений значения рН, ионного состава среды, присутствия мембранотропных веществ и т. д. В условиях нативной клетки липиды в мембранах ведут себя подобно жидким кристаллам. Именно в таком «расплавлен-
ном» состоянии реализуется сочетание упорядоченности с лабильностью. Жидкокристаллическое состояние липидов в бислое обеспечивается молекулярной подвижностью его компонентов, и прежде всего подвижностью углеводородных цепей липидных молекул. Наличие большого числа С–С-связей в углеводородной цепи позволяет ей принимать разнообразные конформации за счет процесса транс-гош-изомеризации. При переходе двух соседних транс-конформаций в гошпроисходит излом цепи и в гидрофобной части бислоя образуется небольшая свободная полость – кинк (от англ. kink – петля).
Однако, вследствие близкого расположения соседних цепей, изомеризация происходит в виде сопряженных поворотов в смежных сегментах С–С-связей. В результате на углеводородной цепи жирной кислоты появляется ряд кинков.
Возникающие кинки постоянно мигрируют вдоль цепи, что приводит к разрыхлению бислоя, появлению в нем подвижных дефектов упаковки липидных молекул, через которые возможно проникновение в мембрану гидрофильных веществ.
Аналогичное влияние на упаковку оказывают двойные цис-связи ненасыщенных жирных кислот в составе фосфолипидов. Их присут-
72

ствие приводит к образованию постоянного кинка в углеводородной цепи.
Подвижностью могут обладать не только отдельные участки углеводородной цепи липидной молекулы, но и вся молекула в целом. Так, она может вращаться вокруг продольной оси, совершать различные перемещения. Миграция молекул вдоль поверхности бислоя называется латеральной диффузией. В жидкокристаллическом состоянии скорость латеральной диффузии молекул липидов достаточно высока. В то же время, переход липидов с одной стороны бислоя на другую – поперечная диффузия (флип-флоп) обычно происходит крайне медленно – фосфолипидной молекуле для пересечения бислоя толщиной в 4-5 нм может понадобиться несколько часов, тогда как в ходе латеральной диффузии она преодолевает это расстояние примерно за 2,5 мкс. Однако в ряде случаев скорость флип-флопа может значительно возрастать под действием некоторых факторов, например таких, как присутствие в бислое молекул, облегчающих перенос через его гидрофобную область полярной головки липидной молекулы.
Рис. Виды подвижности липидов мембраны:
а – латеральная диффузия; б – вращение молекулы вокруг оси; в – поперечная диффузия
или флип-флоп.
В зависимости от состава липидов, условий окружающей среды и других факторов, распределение липидных молекул в плоскости бислоя неоднородно. Липиды способны образовывать упорядоченные области
– кластеры (домены), в которых плотность упаковки нескольких десят- ков-сотен липидных молекул может существенно отличаться от соседних с ними участков. Время жизни кластеров мало и, вследствие неодновременности протекания процессов их распада и образования по всей мембране, более упорядоченные области сосуществуют с уже «расплавленными». В этих условиях для мембран характерно наличие разного рода «дефектов» бислоя, которые могут индуцировать встраивание в
73
мембрану различных соединений, способствовать увеличению ее проницаемости для гидрофильных молекул и ионов, оказывать влияние на взаимодействие белковых молекул друг с другом и т. д.
Таким образом, наличие бислоя липидов, его состояние имеет большое значение для функционирования мембраны.
Функции липидов:
1.Формируют барьер проницаемости для полярных молекул.
2.Обеспечивают жидкокристаллическое состояние (текучесть) мембраны, которое зависит от степени насыщенности радикалов жирных кислот в составе липидов, содержания холестерина, температуры и других факторов. Стабилизация мембран уменьшает функциональную активность клетки.
3.Являются «растворителем» для интегральных белков мембраны и играют важную роль в поддержании их нативной конформации, а, следовательно, и функциональной активности, через изменение состава липидного окружения и его микровязкости (текучести).
Белки мембран
Белки – обязательный компонент мембранных структур. Именно с наличием белковых молекул связано большинство функций, выполняемых мембранами. По степени влияния на структуру бислоя и силе взаимодействия с ним белки делятся на интегральные и периферические.
Интегральные белки – это обычно глобулярные амфифильные структуры, погруженные внутрь липидного бислоя. Среди них выделяют прошивающие белки, которые пронизывают бислой насквозь. Интегральные белки достаточно прочно связаны с липидами за счет гидрофобного взаимодействия, поэтому для их выделения необходимо сначала разрушить двойной слой липидов с помощью детергентов (поверх- ностно-активных веществ) – соединений, солюбилизирующих нерастворимые в воде вещества.
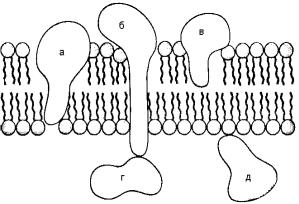
Рис. Варианты расположения белков в мембране:
а, б, в – интегральные белки (а, б – прошивающие); г, д – периферические белки
74

Глубина внедрения интегральных белков в бислой липидов зависит от соотношения гидрофобных и гидрофильных областей в молекуле белка. Гидрофильные участки располагаются на поверхности мембраны (на уровне полярных головок молекул липидов), а гидрофобные – в толще бислоя. В результате взаимодействия белка с липидами может происходить изменение градиента кривизны и деформация бислоя.
Внедрение белка в липидный матрикс упорядочивает последний, делая его структуру более жесткой за счет «прилипания» и ориентации молекул липидов, примыкающих к поверхности белка, где их подви ж- ность затрудняется. То есть, не только липиды влияют на конформацию мембранного белка, но и сам белок модифицирует липидный бислой.
Белки в составе мембраны, также как и липиды, обладают способностью к миграции. Подвижность белка определяется не только его свойствами, но и состоянием липидного окружения. Чем больше микровязкость липидов (ниже текучесть), тем меньше у молекулы белка возможностей для движения.
Периферические белки отличаются от интегральных значительно меньшей глубиной проникновения в бислой. Они, как правило, связаны с мембраной за счет полярных и ионных взаимодействий и не контактируют с гидрофобной частью бислоя липидов. Периферические белки относительно легко экстрагируются из мембраны в мягких условиях, например при промывании буферными растворами с различным значением рН.
На внешней поверхности плазматической мембраны белки часто связаны с углеводами, то есть являются гликопротеинами.
Функции мембранных белков:
1.Регулируют структурное состояние мембраны.
2.Осуществляют трансмембранный перенос веществ.
3.Обеспечивают соединение мембраны с компонентами цитоскелета (белки анкирин, спектрин в эритроцитах).
4.Являются ферментами. Некоторые ферменты локализованы в определенных мембранах и могут служить маркерами этих мембран.
75
Ферменты – маркеры мембран
Мембрана |
Фермент |
Плазматическая |
аденилатциклаза, |
|
γ-глютамилтрансфераза |
Эндоплазматический ретикулум |
Глюкозо-6-фосфатаза |
Аппарат Гольджи |
Галактозилтрансфераза |
Внутренняя мембрана митохондрий |
АТФ-синтаза |
5.Участвуют в межклеточном взаимодействии.
6.Обеспечивают рецепцию внешних сигналов.
7.Выступают в роли антигенов (белок гликофорин в эритроцитах является носителем антигенов групп крови).
Углеводы мембран
Углеводы в составе мембран обнаруживаются только в соединении с белками (гликопротеины) или липидами (гликолипиды). В биологических мембранах может быть гликозилировано до 10 % белков и до 25 % липидов.
Углеводные компоненты преимущественно находятся на внешней стороне плазматической мембраны, встречаются также внутри полости эндоплазматического ретикулума и аппарата Гольджи.
По строению углеводные цепи колеблются от простых моносахаридов до сложных, разветвленных олигосахаридов. В их составе наиболее часто встречаются следующие соединения: галактоза, глюкоза, аминосахара, нейраминовая кислота и ее производные.
Функции углеводов в составе гликопротеинов и гликолипидов мембран:
1.Участвуют в процессах межклеточного взаимодействия.
2.Определяют антигенную специфичность.
3.Обусловливают различия групп крови.
4.Являются рецепторами для связывания различных соединений (гормонов, токсинов и т. д.).
5.Стабилизируют положение белковых и липидных молекул в мембране.
Модели строения мембран
Первые представления о молекулярной организации биологических мембран были высказаны еще в двадцатые годы прошлого столетия. В 1925 году голландские исследователи Э.Гортер и Ф. Грендель высказали гипотезу, что клеточная мембрана представляет собой двойной липидный слой, в котором гидрофильные группы липидных моле-
76

кул локализованы на поверхности бислоя, а углеводородные цепи жирных кислот образуют его гидрофобную внутреннюю область.
Мысль о том, что в состав мембран входят белки впервые выск а- зали в 1935 году Д.Даниелли и Х.Давсон. В соответствии с их гипотезой об общем принципе структурной организации клеточной мембраны, последняя представляется как трехслойная структура, где двойной слой липидов заключен между двумя слоями белка.
В течение следующих десятилетий представления о структурномолекулярной организации мембран сильно менялись. Хотя не возникало никаких сомнений в том, что основными компонентами являются липиды и белки, вопрос об их взаимном расположении стал предметом многочисленных дискуссий.
Современный взгляд на молекулярную организацию биологических мембран начал формироваться в 70-х годах ХХ века. К этому моменту накопилось достаточно много новых экспериментальных фактов, на основании которых американские ученые С.Синджер и Г.Николсон предложили в 1972 году жидко-мозаичную модель строения мембраны.
Рис. Жидко-мозаичная модель Синджера- Николсона
В соответствии с этой моделью, структурной основой биологических мембран является липидный бислой, в котором молекулы липидов находятся в жидкокристаллическом состоянии. Неполярные углеводородные цепи жирнокислотных остатков направлены внутрь бислоя, а полярные головки молекул липидов находятся на его поверхности, контактируя с водой. В липидный бислой, имеющий вязкость растительного масла, погружены молекулы белков. В гидрофобной толще бислоя находится та часть белковой молекулы, которая содержит преимущественно неполярные аминокислоты, а участки, где преобладают полярные аминокислоты располагаются на поверхности липидного бислоя. В противоположность прежним моделям, которые рассматривали мембраны как системы, состоящие из жестко фиксированных элементов, жидко-мозаичная модель представляет мембрану как липидное «море», в котором «плавают айсберги» белков.
77

Таким образом, одним из основных постулатов этой модели является предположение о свободном движении молекул липидов и белков в фазе липидного бислоя. Однако позднее оказалось, что не все белки и липиды способны к свободному перемещению, в некоторых случаях их подвижность сильно ограничена.
Факторы, ограничивающие подвижность липидов и белков в мембране:
1.Образование липидных доменов с более упорядоченной, плотной упаковкой молекул липидов.
2.Способность мембранных белков к агрегации, образованию мультиферментных комплексов.
3.Гликопротеины и гликолипиды образуют на наружной поверхности мембраны ажурную сетку – гликокаликс.
4.Белки со стороны внутренней поверхности мембраны связаны с белковыми нитями цитоскелета.
5.Прошивающие белки образуют сквозную решетку в мембране.
Это привело к модификации жидко-мозаичной модели. Итак, биологическая мембрана – это липидный бислой, начиненный молекулами белка и заключенный в ажурный каркас – решетку.
Рис. Схематическое изображение плазматической мембраны клетки
Решеточно-мозаичная модель отражает одно из главных свойств мембраны – гетерогенность ее механических свойств. Молекулы белка, которые связаны с каркасом, малоподвижны, те же белки, которые с каркасом не связаны, могут относительно свободно перемещаться в плоскости мембраны.
Следует помнить, что современные модели строения мембраны – это лишь упрощенное и схематичное отражение столь сложной и разносторонней системы, какой является биологическая мембрана.
78
Свойства мембран
Общий принцип построения биологических мембран вовсе не означает их однородность. Мембранные системы не только у клеток разного типа, но и в пределах одной и той же клетки отличаются друг от друга как по содержанию и составу основных компонентов, так и по выполняемым функциям. Однако, для всех мембран характерны следующие общие свойства:
1.Мембраны – это ультратонкие пленочные структуры, образующие сплошную перегородку. Протяженность мембран значительно превосходит их толщину.
2.Мембраны состоят в основном из липидов и белков. Углеводы, входящие в состав мембран, связаны с белками или липидами.
3.Мембраны – это надмолекулярные структуры. Входящие в их состав молекулы липидов и белков удерживаются вместе множеством взаимодействий, кооперативных по своему характеру.
4.Мембраны имеют амфифильную природу, так как входящие в их состав липиды и белки содержат как гидрофильные, так и гидрофобные участки. Обе поверхности мембраны гидрофильны, в то время как ее внутренняя область – гидрофобна.
5.Липиды в мембранах находятся в жидкокристаллическом состоянии.
6.Мембраны характеризуются не жесткой, статичной структурой,
адинамичной, подвижной. Подвижность компонентов мембраны определяется двумя видами движения: внутримолекулярным (например, вращение вокруг каждой С–С-связи в цепи остатка жирной кислоты) и межмолекулярным (латеральная диффузия и «флип-флоп»).
7.Мембраны асимметричны функционально и структурно. Функциональная асимметрия проявляется в разделении мембранами внутриклеточного пространства на компартменты. Структурная
– в расположении ее основных компонентов:
•углеводы – преимущественно на внешней поверхности мембраны;
•липиды – фосфатидилхолин и сфингомиелин преобладают в наружном молекулярном слое, а фосфатидилэтаноламин и фосфатидилсерин преимущественно во внутреннем монослое. Содержание холестерина больше в наружном слое мембраны, чем во внутреннем;
•белки – пространственная ориентация белковой молекулы
влипидном бислое, как правило, является однозначной и различные белки занимают строго определенное положение в мембранах. Например, периферические белки, связанные с компонентами
79
цитоскелета локализованы на внутренней стороне плазматической мембраны; переносчики дыхательной цепи расположены зигзагообразно во внутренней мембране митохондрий (цитохром а3 – со стороны матрикса, цитохром с – со стороны межмембранного пространства).
Асимметрия мембраны создается под влиянием различных факторов, например за счет действия ферментов липидного обмена и липидпереносящих белков, участвующих в метаболизме мембранных липидов; различий ионного состава среды по обе стороны бислоя в условиях нативной клетки; особенностей строения молекул фосфолипидов; асимметричной локализации белков в липидах мембраны и т. д. Асимметрия бислоя является важным фактором, обеспечивающим создание градиента кривизны поверхности мембраны, образование складок, везикул.
Биогенез мембран
Динамичность мембранных структур проявляется не только в способности к движению их отдельных компонентов (латеральная и поперечная диффузия молекул липидов и белков), но и в поддержании динамичного равновесия в процессе онтогенеза. Формирование мембраны идет непрерывно, путем введения в ее новых составных частей, обновления компонентов, прежде всего липидов и белков. Биосинтез мембран, как правило, начинается в эндоплазматическом ретикулуме, где образуется бόльшая часть фосфолипидов, холестерина, белков. Затем образовавшиеся мембранные компоненты перемещаются к месту назначения, например в плазматическую мембрану. В этом случае они последовательно проходят через аппарат Гольджи и цитоплазму, модифицируясь в соответствии со своим функциональным назначением (гликозилирование, процессинг и т. п.).
В течение всего времени существования живой клетки все молекулы, входящие в состав мембран многократно обновляются. Время жизни и скорость обновления различных компонентов мембран неодинакова и зависит от интенсивности функционирования клетки. В среднем, из липидов медленнее всего обновляется сфингомиелин – полупериод жизни t0,5 (время, в течении которого заменяется половина исходного содержания молекул) для него составляет около 38 часов. Несколько быстрее обновляется фосфатидилсерин – для него t0,5 равно примерно 23 часа. Для фосфатидилэтаноламина и фосфатидилхолина t0,5 составляет 15 часов. К наиболее быстро обновляемым липидам относится фосфатидилинозит – его обмен может проходить в течение нескольких минут. Это объясняется тем, что фосфатидилинозит принимает участие в трансмембранной передаче сигнала.
80