

Фотосинтез
.pdf11
Фотодыхание и метаболизм гликолевой кислоты. Фермент РУБИСКО может функционировать не только как карбоксилаза, но и как оксигеназа и катализировать окислительное расщепление 1,5-рибулозобисфосфата (РуБФ) до фосфоглицериновой (ФГК) и фосфогликолевой кислоты. Этот процесс называют С2-путь фотосинтеза (поскольку в результате оксигеназной реакции Рубиско образуется С2-кислота) или фотодыхание (поскольку в нем происходит поглощение О2 и выделение СО2).
С2-путь фотосинтеза осуществляется с участием трех органелл — хлоропластов, пероксисом и митохондрий. Расщепление РуБФ до гликолевой и фосфоглицериновой кислот происходит в хлоропластах. Затем гликолат поступает в пероксисомы и там окисляется гликолатоксидазой до глиоксилевой кислоты. Появляющийся при этом пероксид водорода (Н2О2) расщепляется каталазой, а глиоксилат аминируется и превращается в глицин, который поступает в митохондрии. Здесь из двух молекул глицина образуется аминокислота серин и выделяется СО2. Серин далее транспортируется в пероксисому и превращается вначале в гидроксипируват, затем в глицериновую кислоту и, наконец, в фосфоглицериновую кислоту (ФГК).
Однако С2-путь фотосинтеза не всегда является циклом, т. к. иногда процесс идет только в одну сторону и завершается образованием серина и СО2 в митохондриях. Фотодыхание усиливается, когда синтезируется слишком много углеводных продуктов фотосинтеза, а транспортные системы и потребление ассимилятов органами-акцепторами не справляются с их оттоком и утилизацией. В этих условиях углеводы начинают окисляться с образованием аминокислот, которые используются в самой клетке.
У Рубиско способность связывать СО2 значительно выше, чем кислород. Поэтому при естественных концентрациях этих газов в атмосфере (кислорода 21%, а углекислоты 0,036%) доля фотодыхания в фотосинтетическом газообмене листа составляет 25—30%. Однако если концентрация СО2 уменьшается, а кислорода увеличивается, то роль фотодыхания в газообмене листа возрастает. В экстремальной ситуации (полное отсутствие СО2 и 21% концентрации кислорода) фотодыхание превышает фотосинтез и на свету наблюдается не поглощение, а выделение СО2.
Синтез крахмала и сахарозы. Конечными, химически стабильными продуктами фотосинтеза, которые накапливаются в листьях в больших количествах, являются сахароза и крахмал. Впервые образование зерен крахмала при фотосинтезе продемонстрировал в 1864 г. немецкий ботаник Юлиус Сакс. Он помещал зеленые листья на несколько часов в темноту для того, чтобы они израсходовали накопленный в них крахмал. Затем он выносил листья на свет, но при этом освещал только половину каждого листа, оставляя другую половину листа в темноте. Спустя некоторое время весь лист обрабатывался парами йода. В результате освещенная часть листа становилась темно-фиолетовой (в результате образования комплекса крахмала с йодом), в то время как окраска другой половины листа не изменялась.
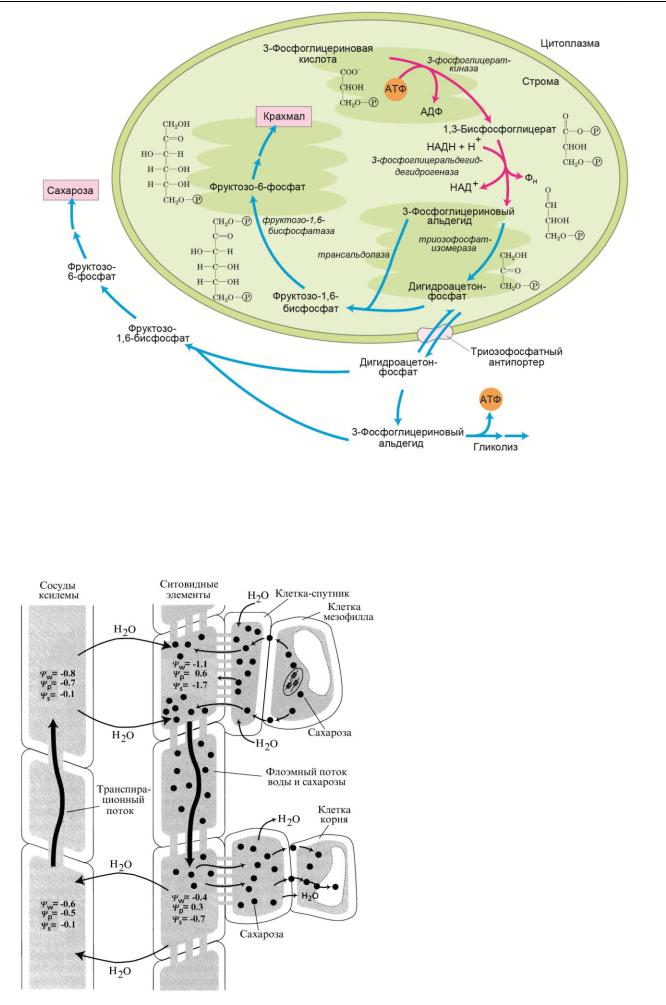
Различают первичный крахмал, который синтезируется в хлоропластах, и вторичный, синтезируемый из сахарозы в лейкопластах запасающих вегетативных органов и плодов. Крахмал, накапливающийся в хлоропластах, не транспортируется. Основной транспортной формой углерода у большинства растений является сахароза, которая синтезируется из фруктозы и глюкозы в цитоплазме. Распределение триозофосфатов между процессами накопления крахмала в хлоропластах и синтеза сахарозы в цитоплазме зависит от обеспеченности этих клеточных компартментов неорганическим фосфатом (ФН). Когда концентрация ФН в цитоплазме велика, триозофосфаты из хлоропластов идут в цитоплазму в обмен на ФН и синтезируется сахароза. Если содержание ФН в цитоплазме падает, триозофосфаты остаются в хлоропласте и синтезируется крахмал.
12
Процесс загрузки сахарозы во флоэмные окончания. Ассимиляты в листе транспортируются от клеток мезофилла по клеткам-спутникам в ситовидные трубки и далее по всему растению. Протонный нанос, функционирующий в плазмалемме клеток-спутников, выкачивает Н+ из клеток. Подкисление апопласта способствует поглощению сахарозы переносчиком, который в режиме симпорта переносит внутрь клеток сахарозу и ионы Н+.
Вода и растворенные в ней вещества передвигаются в соответствии с градиентом водного потенциала.
Возрастание концентрации сахарозы приводит к притоку воды по градиенту осмотического давления и созданию высокого тургорного давления в ситовидных трубках.
Процесс разгрузки сахаров происходит в обратном порядке. Сахароза в симпорте с протоном активным путем выводится из клеток флоэмы. Поскольку при этом происходит снижение осмотического давления, вода выходит из клеток.
Таким образом, движущей силой для потока ассимилятов от мест синтеза к гетеротрофным тканям служит градиент осмотического давления, формируемый вдоль ситовидной трубки.