

Відділ діатомові водорості – bacillariophyta
За морфологічною будовою представники відділу Bacillariophyta досягли великого еволюційного успіху, при цьому залишаючись одноклітинними організмами. Відомо понад 285 родів, що включають від 10 000 до 20 000 видів. За кількістю видів вони конкурують тільки з зеленими водоростями. Деякі вчені вважають, що різноманітність діатомових водоростей ще слабо досліджена і припускають, що фактична кількість видів може перевищити мільйон. Крім цього, представники надзвичайно широко розповсюджені та найбільш чисельні серед еукаріотичних організмів. Вони є домінантними як за біомасою, так і за продуктивністю серед інших водоростей, особливо в прохолодних водах. Велика чисельність при наявності оболонки, що містить кремнезем, призводить до інтенсивного накопичення осадових порід. "Цвітіння" води в морях, викликане масовим розмноженням діатомових водоростей, відоме як коричневий прибій.
Діатомові належать до водоростей-страменопілів. До складу пігментів у них входять фукоксантин та хлорофіли "a" і "c", їх пластиди мають двомембранний хлоропластний ендоплазматичний ретикулум, розташований над двома внутрішніми мембранами оболонки хлоропласту. Запасні продукти – хрізоламінарін і ліпіди. На джгутиках сперматозоїдів (у тих видів, де вони наявні) розташовуються тричленні мастигонеми – ретронеми. Разом з тим, діатомові водорості мають ряд унікальних, характерних тільки для них ознак. Насамперед це стосується будови клітинного покриву, який являє собою твердий кремнеземовий панцир. Панцир діатомей сильно орнаментований. Друга характерна особливість полягає в наявності гаметного мейозу. Джгутиковий апарат у діатомових, як правило, дуже редукований.
Діатомові водорості часто поділяють на дві або три основні групи, в першу чергу за особливостями будови панцира. Центричні діатомеї мають типово дископодібні або циліндричні клітини з радіальною симетрією. Пенатні діатомеї мають більш-менш білатеральну симетрію панцира. Деякі пенатні мають поздовжні шви на панцирі, що пов’язані зі здатністю клітин до руху. Такі діатомеї називають шовними, на відміну від тих пенатних, що не мають швів і які називають безшовними. На основі таких особливостей будови, живі та викопні діатомові водорості були розділені на три групи: центричні, безшовні пенатні та шовні пенатні. Центричні діатомеї, здебільшого, мають багато дископодібних хлоропластів, пенатні – від одного до кількох великих, переважно пластинчастих хлоропластів. Всі пластиди мають піреноїди. Центричні та пенатні відрізняються за типом статевого процесу.
Панцир діатомових водоростей стійкий до руйнування в навколишньому середовищі і тому, за палеонтологічними даними, філогенія діатомових водоростей досить добре досліджена, починаючи з Ранньої Крейди (150 млн. років тому). Найдревніші скам’янілі діатомеї відносяться до центричних, в той же час, як безшовні форми пенатних відомі з Пізньої Крейди (близько 70 млн. років тому). Шовні пенатні з’явилися біля 60 млн. років тому і зайняли панівне положення у фітопланктоні у Міоцені (близько 24 млн. років тому).
Молекулярно-філогенетичні дослідження (рис. 88) свідчать про наявність двох філогенетичних гілок в еволюції відділу. Перша гілка включає центричні діатомеї, більшість з яких позбавлена додаткових вставних обідків. Друга гілка включає центричні діатомеї з додатковими вставними обідками, а також пенатні діатомові водорості. Молекулярні дані підтверджують також монофілетичне походження шовних пенатних.
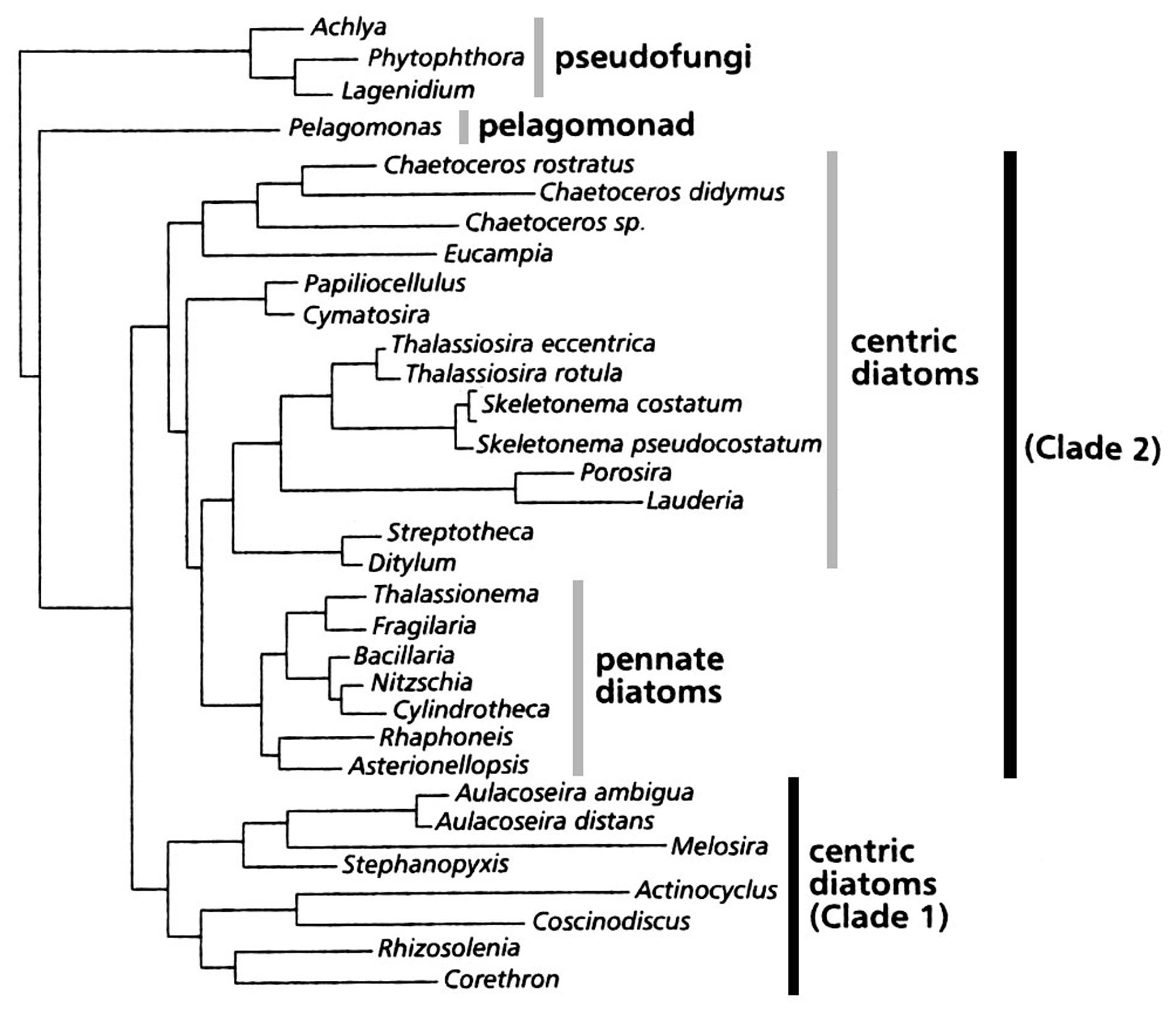
Рис. 88. Філогенетичне дерево Bacillariophyta, побудоване за даними аналізу послідовностей макромолекул (за: Medlin et al., 1997).
У діатомових водоростей джгутики зустрічаються тільки у сперматозоїдів центричних водоростей і зовсім не характерні для пенатних. Найімовірніше вони були втрачені в процесі еволюції. Еволюційне походження Bacillariophyta досі дискусійне. Донедавна вважалося, що вони походять від джгутикових Chrysophyta. Сучасні дослідження показують, що предками діатомей могли бути джгутиконосці, подібні до нещодавно описаного таксону Bolidophyta з бурої пігментної групи. Ці водорості мають дрібні (1,2 мкм. у діаметрі) безпокривні клітини та два гетероморфні джгутики, один з яких перистий, а другий – гладкий. Молекулярно-біологічні дані також підтверджують спорідненість цих таксонів. Розвиток кремнієвого покриву, як вважається, забезпечив адаптивні переваги. Кремній інертний до ферментативного впливу і, таким чином, діатомеї більш стійкі до мікробних атак та гірше засвоюються тваринами, ніж безпокривні клітини або такі, що мають оболонку з полісахаридів. Крім того кремній є дуже доступним елементом у природі.
Клітинний покрив утворений прозорим кремнієвим панциром. Структура панцира, форма, співвідношення осей і площин симетрії лежать в основі системи діатомових водоростей (рис. 89). Панцир складається із аморфного кремнію, що за складом нагадує опал (Si2 x xH2O) з домішкою металів: алюмінію, заліза, магнію. Панцир складається із двох частин-стулок, епітеки і гіпотеки. Епітека – більша і насувається краями на гіпотеку. Епітека складається із плоскої або опуклої стулки епівальви і пояскового обідка – епіцингулюма; гіпотека – із гіповальви і гіпоцингулюма. Епі- і гіпоцингулюм складають поясок панцира. Клітину можна бачити або з боку стулки, або з боку пояска. У деяких Bacillariophyta утворюються додаткові пояскові кільця або вставні обідки (копули).
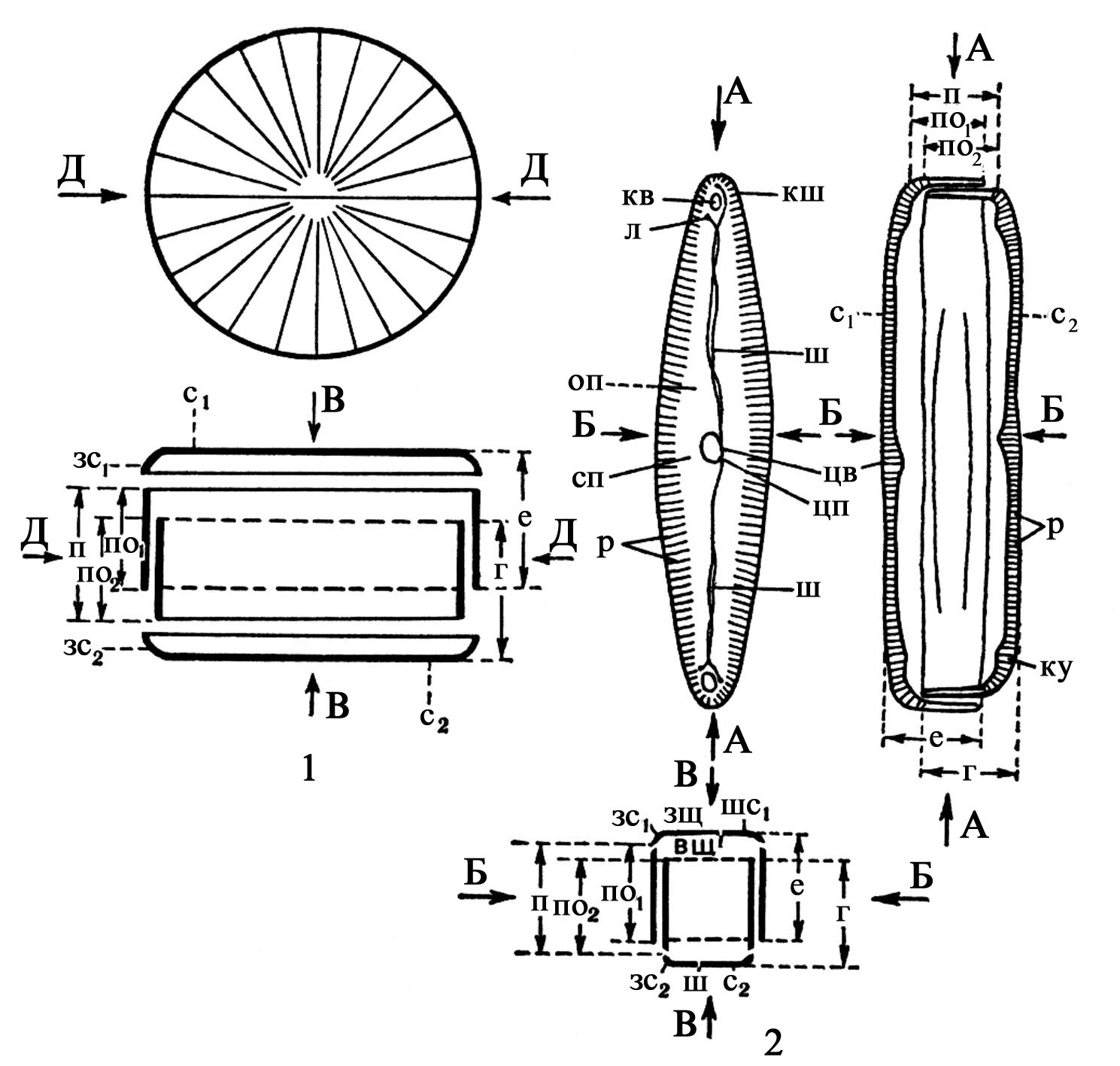
Рис. 89. Схема будови панцира Bacillariophyta:
1 – центричного типу: вверху – вигляд зі стулки, знизу – вигляд з пояска; ДД – діаметр панцира, ВВ – центральна вісь панцира; 2 – пенатного типу: вверху зліва – вигляд зі стулки, вверху справа – вигляд з пояска, знизу – переріз панцира впоперек; АА – поздовжня вісь, ББ – поперечна вісь, ВВ – вертикальна (первольварна) вісь панцира, е – епітека, г – гіпотека, с1 – стулка епітеки, с2 – стулка гіпотеки, зс1 – загин стулки епітеки, зс2 – загин стулки гіпотеки, по1 – поясковий обідок епітеки, по2 – поясковий обідок гіпотеки, р – ребра (ряди альвеол), цв – центральний вузол, цп – центральна пора, ш – шов, кв – кінцевий (полярний) вузол, кщ – кінцева (полярна) щілина, л – лійка, оп – осьове поле, сп – середнє поле, вщ – внутрішня щілина шва, зщ – зовнішня щілина шва.
Розрізняють два основні типи стулок: актиноморфні, через які можна провести три або більше площин симетрії і зигоморфні – продовгуваті, з бісиметричною (перистою) структурою, через які можна провести не більше двох площин симетрії. Перші виділяють у клас Centrophyceae, а другі – у клас Pennatophyceae.
Структура панцира специфічна для різних таксонів у межах класів і утворена різними структурними елементами, з яких загальними і найбільш важливими є перфорації – система отворів різної будови, що розміщені на стулках, через які відбувається зв’язок протопласта із зовнішнім середовищем. Під мікроскопом їх можна бачити у вигляді крапок, штрихів, ребер, комірок. Для багатьох планктонних форм характерні вирости на панцирі. Бентосні діатомеї із Pennatophyceae здатні до активного руху. Вміст клітин сполучається із зовнішнім середовищем поздовжньою щілиною або швом, що проходить безпосередньо зі стулкового боку або по особливих виростах - кілях, що розміщені по довжині довгої осі стулки або її краях. З рухом зв’язані вузлики, що є внутрішніми потовщеннями стінок стулки, які мають вигляд трьох невеликих круглих опуклостей: одна в центрі і по одній на кінчиках зі стулкового боку панцира. Шви бувають щілиноподібні і каналоподібні. Вони забезпечують зв’язок протопласта із зовнішнім середовищем і здатність до руху.
У філогенетичному відношенні наявність шва – прогресивна ознака, яка характерна для еволюційно більш молодих видів. За допомогою швів здійснюється унікальний рух клітин. Рух – складний процес і є наслідком переміщення цитоплазми у шві і вертикальних каналах, що проходять у вузликах. У русі приймають участь дві системи органел: фіброзні тяжі і, можливо, світлозаломлюючі частинки, які є біля шва у кожному кінці клітини. Вони виділяють локомоторну речовину через кінцеві пори і по всій довжині шва. Вважають, що рух обумовлений струменями цитоплазми в щілині (каналі шва) або струменями води у порожнині шва. Є думка, що фібрили скорочуються і з порожнини шва назовні виходить речовина, що ніби приклеює клітину до субстрату і залишає на ньому слід. Довгі фіброзні тяжі, що знаходяться під швом, рухаються завдяки локомоторній речовині, що виділяється світлочутливими частинками через пори і по всій довжині шва.
Будова клітини. Цитоплазма розміщена тонким пристінним шаром, а у деяких – і в середині клітини, де знаходиться ядро (рис. 90). В клітині є одна або декілька вакуолей. Хлоропласти у Centrophyceae – дрібні, багаточисленні, у вигляді дисків. Такі ж хлоропласти у безшовних Pennatophyceae. У більш високоорганізованих – крупні, у вигляді двох стрічок або перфоровані. В хлоропластах міститься хлорофіл “а” і “с”, - і е-каротин, п’ять ксантофілів, в тому числі діадиноксантин, діатоксантин, фукоксантин, нефукоксантин А і В. У стромі розміщені тилакоїди, вони не утворюють грани, а утворюють пачки із трьох цистерн (тритилакоїдні ламели), є один оперізуючий тилакоїд. Хлоропласт містить один або декілька піреноїдів. Іноді піреноїд має ламелярну структуру. Продукти асиміляції – волютин, хризоламінарин, ліпіди. Діктіосомний апарат добре розвинений.
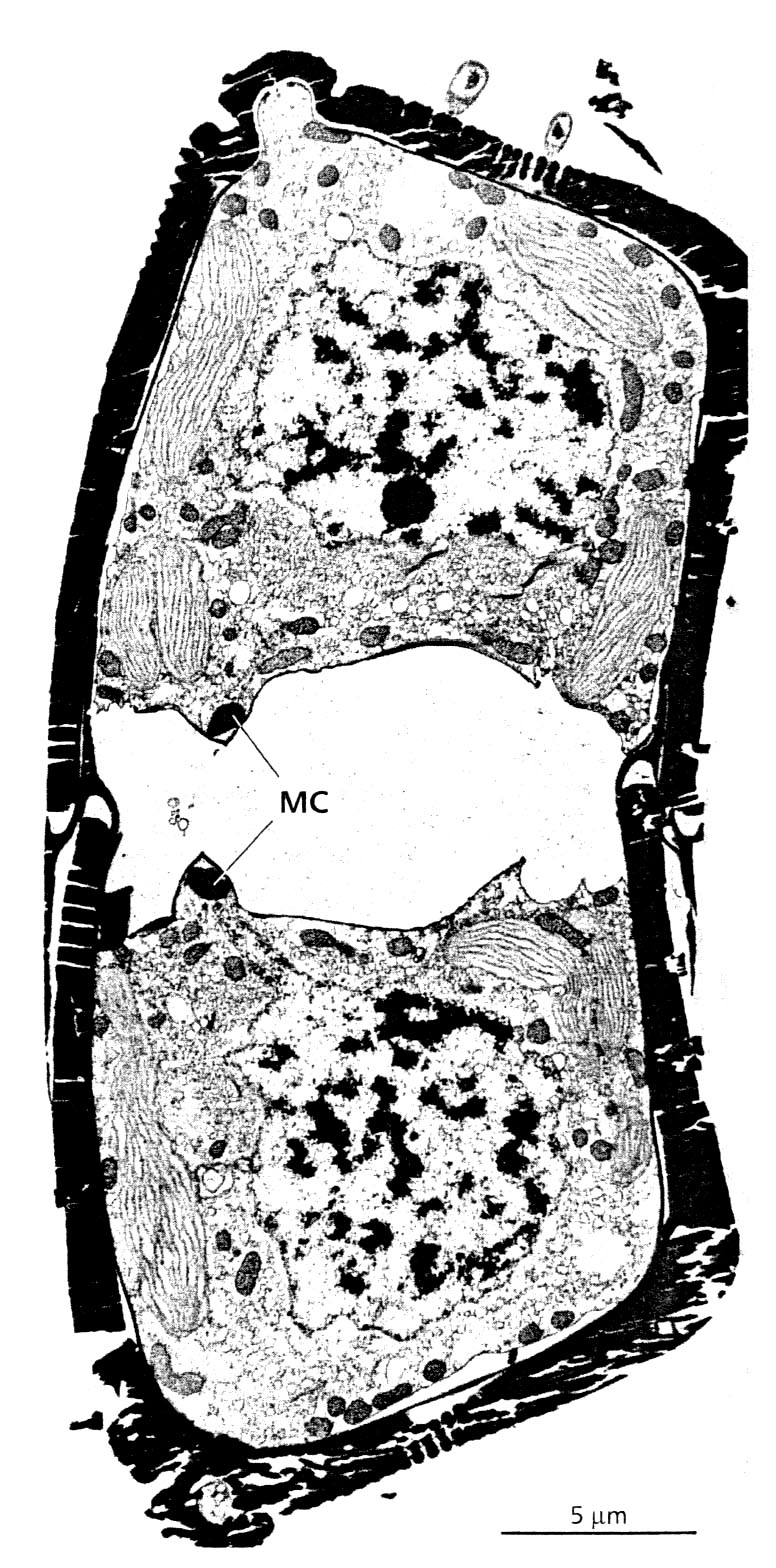
Рис. 90. Будова клітини Hantzschia amphioxys під електронним мікроскопом. Видно дві дочірні клітини, що недавно утворилися, йде утворення кремнієвого панцира, а також видно клітинний центр, що утворений мікротрубочками (МС) в кожній дочірній клітині (за: Pickett-Heaps & Kowalski, 1981).
Розмноження – вегетативне, поділом клітини. Є статевий процес. При вегетативному поділі протопласт збільшується, розсуває епітету і гіпотеку, мітозу передує і поділ хлоропластів. Загальна схема мітозу у Centrophyceae i Pennatophyceae, на ультраструктурному рівні, однакова (рис. 90). Кожний новий протопласт успадковує половину панцира, а друга половина панцира утворюється заново, причому у обох дочірніх клітин добудовується гіпотека (рис. 91). При кожному поділі одна буде материнською, а друга буде меншою і, внаслідок подальших поділів клітин в популяціях, клітини прогресивно зменшуються (рис. 92). Відновлення первинних розмірів клітин відбувається під час проростання відпочиваючих спор і внаслідок статевого процесу, який супроводжується утворенням ауксоспор.
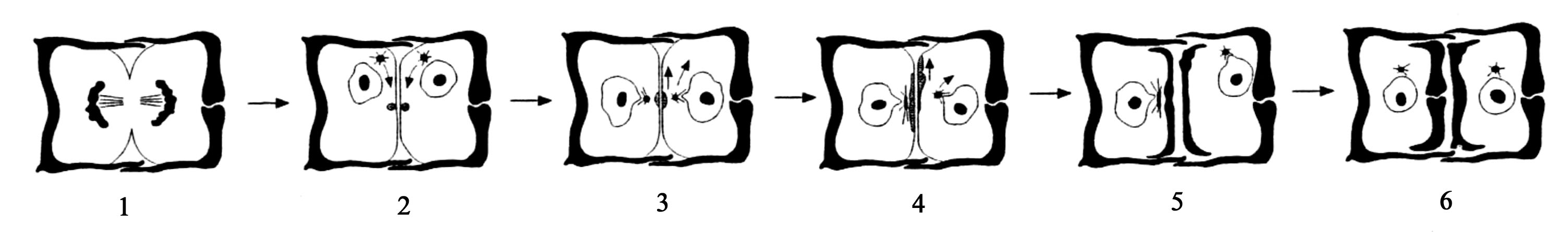
Рис. 91. Схема утворення панцира у дочірніх клітин при поділі Achnanthes coarctata (за Boyle, et al., 1984):
1-6 етапи утворення гіпотеки в дочірніх клітинах.
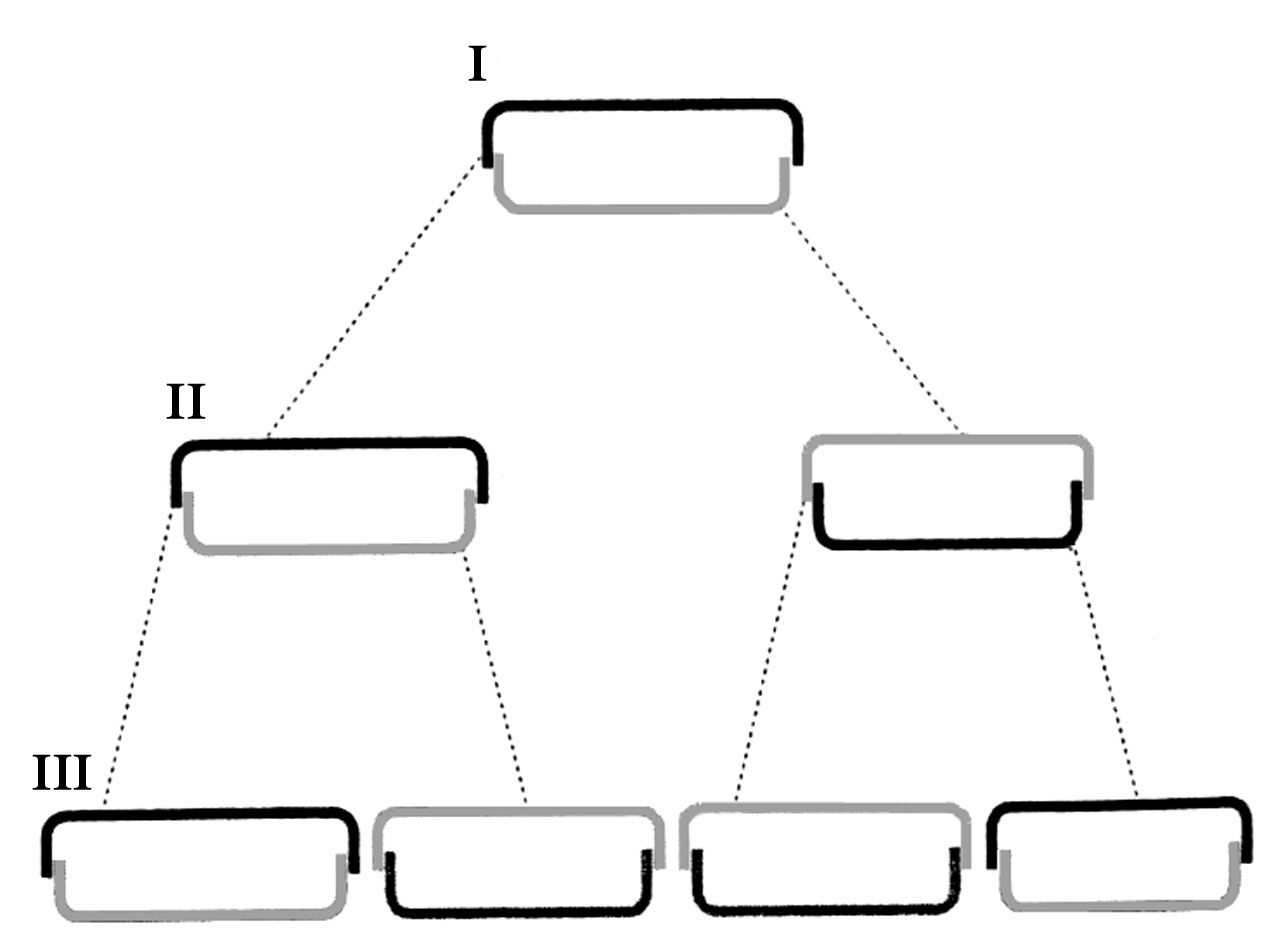
Рис. 92. Три послідовні поділи клітин (I-III) у діатомей:
У більшості видів статевий процес починається тоді, коли розміри клітини зменшаться приблизно до 1/3 їх максимального розміру. Вважається, що відновлення розміру клітини є основною функцією статевого процесу. Якщо розміри клітини зменшаться на величину меншу, ніж 1/3 максимального розміру, то такі клітини нездатні до статевого процесу і поступово відмирають.
Клітини діатомей мають диплоїдний набір хромосом і утворенню гамет передує мейоз. Диплоїдність і, як наслідок цього – гетерозиготність, збільшують генетичну та еволюційну різноманітність. Відносно мало відомо про стимуляційну дію зовнішніх факторів на статевий процес у діатомей. Встановлені стимулюючий вплив підвищення температури, збільшення інтенсивності освітлення та трофічності середовища на ініціацію статевого розмноження у деяких видів. Існує припущення про наявність феромонів у Bacillariophyta, але їх хімічна природа не встановлена. Вважається, що статевий процес у діатомових водоростей відбувається частіше, ніж це спостерігали дослідники.
Центричні діатомеї переважно оогамні (рис. 93А). В жіночих клітинах утворюється одна або дві яйцеклітини (дві або три клітини з чотирьох, що утворюються внаслідок мейозу – відмирають). В чоловічих клітинах утворюється 4-128 сперматозоїдів або нерухомих спермаціїв. Сперматозоїди – рухомі, мають редукований джгутиковий апарат, представлений єдиним перистим джгутиком. Джгутик несе два ряди ретронем, але при цьому часто не має центральної пари мікротрубочок (формула 9+0 або 9+1). Базальні тільця джгутиків складаються з дублетів мікротрубочок, а не триплетів, як у більшості джгутиконосців. Антеридії мають отвір, через який звільняються сперматозоїди. Кремнієві стулки оогоніїв також розсуваються, оголюючи протопласт яйцеклітин. Гамети голі, і мають пластиди. Внаслідок злиття гамет утворюється велика ауксоспора
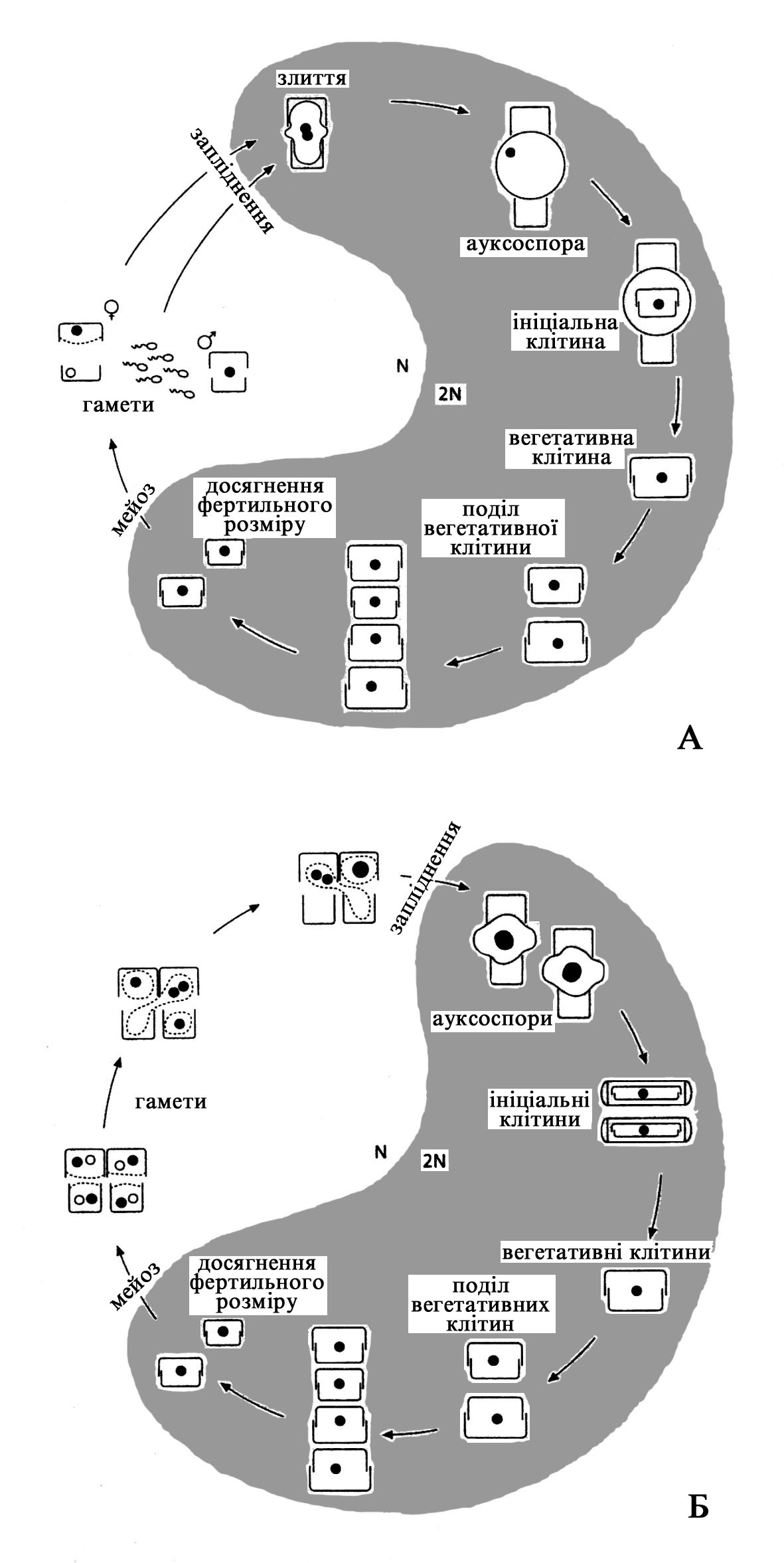
Рис. 93. Схема життєвого циклу Bacillariophyta (за: Hasle & Syvertsen, 1997):
А – життєвий цикл типового представника Centrophyceae, Б – життєвий цикл типового представника Pennatophyceae.
Ауксоспори можуть бути: вільними від батьківського панцира, приєднаними до однієї стулки або мати дві стулки. Декілька родів, які розмножувалися оогамно, але мали білатеральну симетрію панцира (і за цією ознакою розглядалися як пенатні), тепер розглядаються як центричні на основі статевого розмноження.
Представники Pennatophyceae майже завжди ізогамні, дві гамети одинакові за розмірами і обидві безджгутикові (рис. 93Б). Виключення становить Rhabdonema – безшовна пенатна діатомея, у якої статевий процес описаний як видозмінена оогамія. У цієї водорості утворюється велика гамета – яйцеклітина, і маленька безджгутикова, що здатна до амебоїдного руху. Статевий процес у Rhabdonema розглядається як проміжний етап переходу від оогамії у центричних до ізогамії у пенатних, але це припущення потребує додаткового дослідження. Статеве розмноження більшості Pennatophyceae починається з’єднанням батьківських клітин спільним слизом. Нерівний цитокінез слідує за першим мейотичним поділом і менші з утворених клітин – гинуть. Стулки панцирів відкриваються до завершення другого поділу мейозу і протопласти звільняються. В межах кожного з двох протопластів гаплоїдне ядро дегенерує, а друге залишається живим. Постмейотичні протопласти містять по одному гаплоїдному ядру. Згодом протопласти зливаються і утворюють двоядерну клітину, що збільшується в розмірах, та формують ініціальну клітину, вкриту суцільною оболонкою. Згодом відбувається злиття ядер і відновлюється диплоїдність клітини. У деяких Pennatophyceae залишаються живими по два протопласти та утворюється дві ауксоспори.
У деяких діатомових ауксоспори утворюються шляхом автогамії, в процесі чого два гаплоїдних ядра, що виникають внаслідок мейозу, зливаються. Відомо також утворення ауксоспори шляхом апогамії, при якій ніякого злиття ядер не відбувається.
В сприятливих умовах, після серії поділів, з ауксоспор розвиваються вегетативні клітини звичайної будови максимально можливого розміру.
Утворення спор. У багатьох морських планктонних Bacillariophyta в клітинах розвиваються дрібні тільця – мікроспори. Вони бувають з джгутиками або без них, з хлоропластами або безбарвні. Утворення спор особливо характерно для видів роду Chaetoceros, у яких спостерігалося їх проростання. Способи утворення та природа мікроспор не досліджені.
При настанні несприятливих умов Bacillariophyta переходять в стан спокою. Під час утворення спочиваючих клітин протопласт пересувається до одного з кінців клітини і, внаслідок втрати клітинного соку, сильно стискається. Життєдіяльність таких клітин відновлюється при настанні сприятливих умов. Деякі прісноводні планктонні види в зимовий період опускаются на дно водойм, де перебувають в стані спокою до початку вегетаційного періоду.
У більшості Bacillariophyta утворенню спочиваючих спор передує вегетативний поділ клітини на дві, кожна з яких, в подальшому, стає материнською клітиною спори (рис. 94). Протопласт материнської клітини стискається, заокруглюється, на його поверхні виникає первинна стулка спори, згодом – вторинна, що насувається своїми краями на первинну. Вміст спори гомогенний. Структура стулок спори постійна для кожного виду і відрізняється від будови вегетативної клітини. Як правило, материнська клітина утворює одну екзогенну, напівендогенну або ендогенну спору. Зріла екзогенна спора знаходиться поза оболонкою материнської клітини. Одна стулка зрілої напівендогенної спори включена в материнську клітину, інша – залишається вільною. Зріла ендогенна спора знаходиться всередині материнської клітини. Спора проростає у вегетативну клітину, розмір якої значно перевищує розмір самої спори. Спори в бентосних відкладеннях можуть зберігати життєвість протягом десятиріч.

Рис. 94. Спори видів роду Chaetoceros:
а – ланцюжок з молодими спорами Chaetoceros abnormis, б – Ch. seiracanthus, в – Ch. subsecundus, г – Ch. lorenzianus, д, е – Ch. paulsenii, є – Ch. compressus, ж – Ch. rigidus.
Bacillariophyta мають широке розповсюдження і заселяють прісні і солоні, стоячі і проточні водойми, вологі скелі і, навіть, орні землі. Група дуже пластична до різних екологічних факторів зовнішнього середовища. У водних екосистемах вони є домінантами цілий рік. Беруть участь в утворенні різних фітоценозів усіх типів. У планктоні морів і океанів переважають Centrophyceae. Бентосні фітоценози дуже різноманітні і тут домінують Pennatophyceae. Bacillariophyta беруть участь в обростанні інших макроскопічних водоростей (епіфіти). Ендосимбіотично існують у протопластах форамініфер, є господарями паразитів. Серед них виявлені гетеротрофи (Synedra, Nitzschia). Diatomeae вегетують в діапазоні від 0 до 700С. Багата діатомеями флора евтрофних солоних озер.
Масовий розвиток деяких Bacillariophyta сприяє відмиранню личинок риб, викликає “цвітіння” води. Велике значення діатомей як ланки у трофічних зв’язках водних екосистем та в екологічному моніторингу. Вони є індикаторами забруднення водного середовища (Nitzschia angustata, Cymbella cesatii). Важлива роль водоростей полягає у нагромадженні осаду. При відмиранні діатомей на дні водойм утворюється діатомовий і сапропелевий намул (осад). Діатоміти бувають товщиною декілька сотень метрів. Чорна мука на 80% складається із панцирів викопних діатомей, які відомі з юрського періоду. При гороутворенні на поверхню були винесені відклади панцирів. Це біла або світло-сірого кольору маса, легка, пориста і тверда, яка використовується як шліфувальний та ізоляційний матеріал, а також як фільтруюча речовина в медичній, хімічній і харчовій промисловості, а також при виготовленні динаміту. В геології викопні діатомеї використовують для визначення віку осадових порід (діатомовий аналіз).
Існує декілька поглядів на таксономію відділу Bacillariophyta. Зокрема, деякі дослідники виділяють у відділі три класи: Coscinophyceae (до якого відносять центричні діатомові), Fragilariophyceae (що об’єднує безшовні пенатні) та Bacillariophyceae (що об’єднує шовні пенатні). Молекулярні данні не підтверджують поділ відділу на три класи. В цій книзі ми будемо дотримуватися класичного поділу відділу Bacillariophyta на класи Centrophyceae та Pennatophyceae.