

8.3. Общие свойства ферментов
К ферментам применимы три основных критерия, характерных для неорганических катализаторов:
они остаются относительно неизменными после реакции, т. е. освобождаются вновь и могут реагировать с новыми молекулами субстрата;
ферменты оказывают свое действие в ничтожно малых концентрациях, например: 1 молекула реннина (содержится в слизистой оболочке желудка теленка) створаживает около 106 молекул казеиногена молока за 10 мин при 37 °С;
наличие фермента (катализатора) не оказывает влияния ни на величину константы равновесия, ни на изменения свободной энергии. Катализаторы лишь повышают скорость реакций (точку равновесия они не сдвигают).
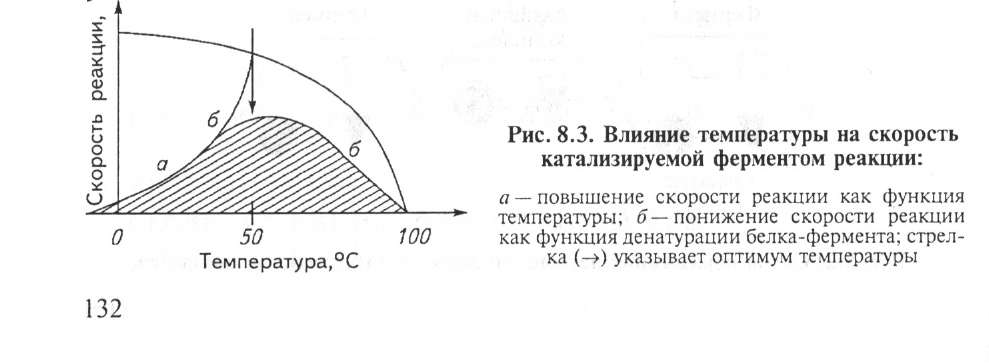
Термолабильность ферментов. Скорость химической реакции зависит от температуры. Реакции, катализируемые ферментами, также чувствительны к изменениям температуры. Скорость химической реакции повышается в 2 раза при повышении температуры на 10 "С. Ускорение реакции идет до 45 "С, затем, в связи с денатурацией белка-фермента, снижается. При 100 "С почти все ферменты полностью утрачивают свою активность.
Оптимальная температура для действия ферментов теплокровных 37...40 "С. При низких температурах (0 °С и ниже) ферменты не разрушаются, не денатурируют, но активность их снижается до нуля (рис. 8.3).
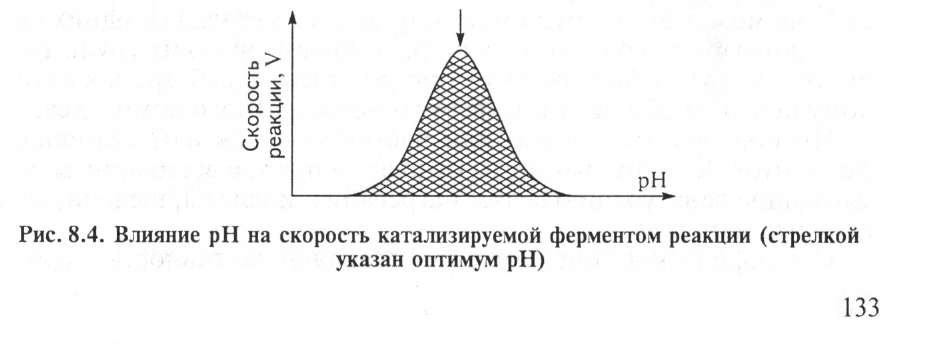
Зависимость активности ферментов от рН среды. Ферменты активны в узких пределах значений рН, как правило, 6,0...8,0. Каждый фермент имеет свой оптимум рН, который лежит в пределах физиологических значений. Исключением является пепсин, рН- оптимум которого 2,0. Объясняется это функцией данного фермента, так как в желудочном соке содержится свободная хлороводородная (соляная) кислота, обусловливающая кислое значение рН.
Влияние колебаний рН среды на активность фермента заключается в том, что изменяется сте-
пень ионизации кислотных и основных групп (—СООН, —SH, имидазольной, —NH2 и т. д.).
При разных значениях рН активный центр может находиться в частично ионизированной или неионизированной форме, что сказывается на третичной структуре белка и соответственно на формировании активного комплекса фермент—субстрат (рис. 8.4).
Специфичность ферментов. Ферменты обладают высокой специфичностью действия и этим существенно отличаются от неорганических катализаторов. Например, измельченная платина или палладий могут катализировать восстановление десятков соединений различной структуры, а фермент обычно проявляет свою активность только при полном соответствии активного центра и субстрата, только при «узнавании» обычно протекает одна высокоспецифичная реакция среди тысячи других химических процессов, осуществляемых в живой клетке. В зависимости от механизма действия различают ферменты с относительной, или групповой, и абсолютной специфичностью.
Для действия некоторых гидролитических ферментов характерна относительная специфичность. Например, пепсин расщепляет белки животного и растительного происхождения, хотя они могут различаться по химическому строению, аминокислотному составу и физико-химическим свойствам; он разрывает пептидную связь — СО—NH— между аминокислотами. Однако пепсин не расщепляет жиры и углеводы.
Липаза, катализирующая гидролиз жиров на глицерин и жирные кислоты, разрывает сложноэфирные связи. Аналогично групповой специфичностью обладают трипсин; химотрипсин; пептидазы; ферменты, гидролизирующие а-гликозидные связи в полисахаридах. Все перечисленные ферменты — пищеварительные, их групповая специфичность имеет биологический смысл.
Абсолютная специфичность — это способность фермента катализировать превращение единственного субстрата. Например, аргиназа, расщепляющая в организме аргинин; уреаза, катализирующая распад мочевины, и др.
Стереохимическая специфичность — это способность фермента различать L- и D-изомеры. Например, оксидазы (аминокислот) действуют только на свой специфический стереоизомер L- или D-аминокислот. Специфичностью обладают также ферменты, катализирующие синтетические реакции.