

КАРАГАНДИНСКАЯ ГОСУДАРСТВЕННАЯ МЕДИЦИНСКАЯ АКАДЕМИЯ
Кафедра биологической химии
УЧЕБНОЕ ПОСОБИЕ
По теме «Строение и функции нуклеиновых кислот»
(для студентов всех факультетов)
СОСТАВИТЕЛИ: зав. каф. проф. Л.Е. Муравлева, доцент Т.С. Омаров, преподаватель Д.А. Клюев
Караганда 2004
1.Днк, строение функции.
Нуклеиновые кислоты - гетерополимеры, их мономерами являются мононуклеотиды. В состав мононуклеотида входит пуриновое или пиримидиновое азотистое основание, пентоза и остаток фосфорной кислоты. К пуриновым основаниям относят аденин и гуанин, к пиримидиновым - цитозин, тимин и урацил. (рис.1).
Пентоза: рибоза или дезоксирибоза входят в состав НК в фуранозной форме и связаны с азотистым основанием N-гликозидной связью. Такое соединение называется нуклеозид. В случае образования связи с пуринами к названию добавляется суффикс –озин: аденозин, гуанозин, а пиримидинами – идин: тимидин, цитидин (рис.1). Так, например, аденин и рибоза образуют нуклеозид аденозин (1, сокращенно А). Соответствующие производные других азотистых оснований носят названия гуанозин (G), уридин (U), тимидин (T) и цитидин (С). Если углеводный остаток представлен 2-дезоксирибозой, образуется дезоксинуклеозид, например 2'-дезоксиаденозин. В клетке 5'-ОН-группа углеводного остатка нуклеозида этерифицирована фосфорной кислотой. Соответствующее производное 2'-дезокситимидина (dT), звено ДНК, называется 2'-дезокситимидин-5'-монофосфат (dTMP) (2). Если 5'-фосфатный остаток соединяется с другими нуклеозидфосфатными остатками. получаются нуклеозидди- и нуклеозидтрифосфаты, например АДФ и АТФ — важнейшие коферменты энергообмена.
Остаток фосфорной кислоты (H3PO4) присоединяется к пентозе обычно в 5-ом положении. Название такого соединения образуется от названия соответствующего нуклеозида с указанием места присоединения фосфорной кислоты и добавлением слова «фосфат». В зависимости от количества остатков существуют моно-, ди- и трифосфаты (аденозин–5–трифосфат, АТФ, ГМФ, ЦМФ, ТДФ, УТФ.ТМФ встречается только в ДНК, а УМФ - только в РНК. Все нуклеозидфосфаты объединяют под общим названием нуклеотиды.
Остатки фосфорной кислоты могут связываться за счет образования фосфоангидридной связи. Следовательно, два нуклеотида могут быть связаны через фосфатные группировки с образованием соответствующего динуклеотида. К этой группе соединений относятся коферменты [HAДФ+(NADP+)] и KoA (CoA), а также флавин [ФАД(FAD)].
Если фосфатная группа одного нуклеотида взаимодействует с З'-ОН-группой другого нукпеотида, образуется динуклеотид с фосфодиэфирной связью. Такой динуклеотид несет на 5'-конце свободную фосфатную группу, а на 3'-конце свободную ОН-группу. Поэтому можно за счет образования еще одной фосфодиэфирной связи присоединить новый мононуклеотид. Таким путем образуются олигонуклеотиды и, наконец, полинуклеотиды.
Полинуклеотиды, составленные из рибонуклеотидных звеньев, называются рибонуклеиновыми кислотами (РНК), из дезоксирибонуклеотидных мономеров — дезоксирибонуклеиновыми кислотами (ДНК). При обозначении полинуклеотидов указывают сокращенные названия нуклеозидных звеньев в направлении 5'→3', т.е. слева направо. Иногда в название включают фосфатную группу ("p")..
Первичная структура цепей ДНК - это порядок чередования дезоксирибонуклеозидмонофосфатов полинуклеотидной цепи.
Вторичная структура ДНК представляет собой правозакрученную спираль, состоящую из двух полинуклеотидных цепей с антипараллельным ходом, закрученными относительно друг друга и вокруг общей оси. Антипараллельный ход означает, что 3’-концу одной цепи соответствует 5’-конец другой цепи и наоборот.
Остатки оснований направлены внутрь спирали. На один виток спирали приходится 10 пар оснований. Цепи ДНК не идентичны, так как нуклеотидный состав их различен, однако первичная структура одной цепи предопределяет нуклеотидную последовательность другой цепи, то есть они комплементарны друг другу.
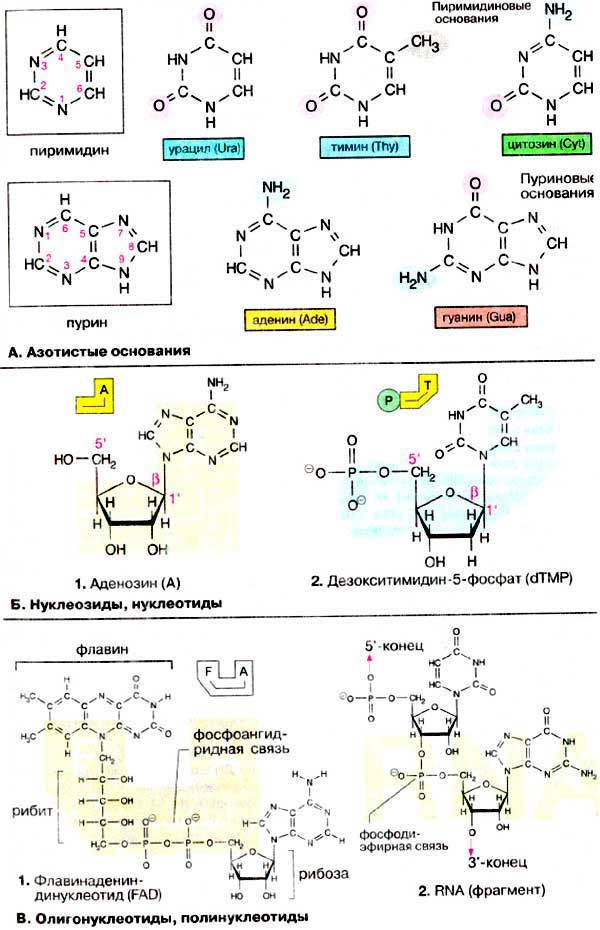
Рисунок 1 Структура азотистых оснований, олигонуклеотидов
Комплементарность А и T, соответственно G и С, становится понятной, если рассмотреть возможные водородные мостики между основаниями. В качестве доноров выступают аминогруппы (аденина, цитозина, гуанина) и NН-группы гетероциклов (тимина и гуанина). Возможными акцепторами являются карбонильные группы (тимина, цитозина, гуанина) и атомы азота гетероциклов. Пара A-T может образовывать два, а пара G-C даже три линейных и поэтому особенно устойчивых мостика. Урацил, содержащийся в РНК вместо тимина, ведет себя при спаривании основании подобно тимину.
Стабилизируют двойную спираль также гидрофобные взаимодействия, возникающие между основаниями двуцепочечной молекулы.
Нуклеиновые кислоты встречаются в организме не в свободном виде, а в составе нуклеопротеинов. Каждая молекула ДНК упакована в отдельную хромосому. Все связывающиеся с ДНК эукариотов белки можно разделить на 2 группы: гистоны и негистоновые белки. Комплекс белков с ядерной ДНК клеток называют хроматином.
Гистоны — это белки небольшого размера (мол. масса около 20 000) с очень высоким содержанием положительно заряженных аминокислот (лизина и аргинина). Хроматин содержит 5 типов гистонов: Н2А, Н2В, НЗ, Н4 (нуклеосомные гистоны) и H1. Суммарный положительный заряд позволяет им прочно связываться с ДНК.
Негистоновые белки — это разные типы регуляторных белков, связывающихся со специфическими последовательностями ДНК, а также ферменты, участвующие в матричных биосинтезах.