

Современное представление о механизме инстинктивного поведения. Ключевые раздражители и пусковые ситуации.
Современное понимание проблемы инстинкта и научения
В проблеме инстинкта и научения большое место занимает вопрос о пластичности инстинктивного поведения. Этот вопрос является весьма важным для уяснения не только эволюции инстинктивного поведения, но и вообще всех вопросов, касающихся психической деятельности животных.
Дарвин считал, что, по существу, одна пластичность инстинктов, проистекающая из изменчивости их врожденных морфологических основ и дающая «материал» для действия естественного отбора, является достаточной для эволюции инстинктивного поведения, а тем самым и поведения вообще. Впоследствии многие ученые посвятили свои усилия изучению того, насколько врожденное, ви-дотипичное поведение является устойчивым или вариабельным, насколько инстинкты постоянны, ригидны или же изменчивы и могут модифицироваться. В результате сегодня мы знаем, что пластичность поведения животных является значительно более сложным явлением, чем это представлялось во времена Дарвина, ибо генетически фиксированы и передаются по наследству не отдельные готовые движения или их сочетания, а нормы реагирования, в пределах которых формируются двигательные реакции в онтогенезе.
Глубокую разработку проблемы инстинкта и научения, как отмечалось, дал В.А.Вагнер, особенно в своем фундаментальном труде «Биологические основания сравнительной психологии» (1910—1913). Опираясь на большой фактический материал, полученный им в полевых наблюдениях и экспериментах и охватывающий как беспозвоночных, так и позвоночных, Вагнер пришел к выводу, что инстинктивные компоненты поведения животных воз-никли и развились под диктовку среды и под контролем естественного отбора и что их никак нельзя считать неизменными, стереотипными. Инстинктивное поведение, по Вагнеру, — это развивающаяся пластическая деятельность, изменяемая внешними воздействиями.
53
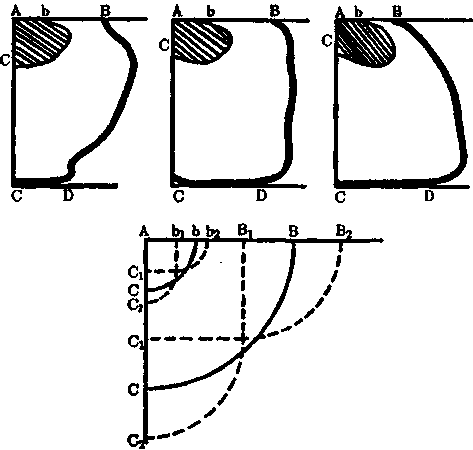
Рис. 4. Гнездостроение у городской ласточки (по Вагнеру). Верхний ряд — контуры разных форм ласточкиных гнезд. Постройки различаются, в частности, относительными размерами крыши (АВ) и дна (CD) гнезда (вид сбоку). Abe — леток. Внизу—схема Вагнера, изображающая вариабельность гнездострои-тельного поведения. Внутренняя дуга означает среднюю форму и величину летка, внешняя — гнезда. Пунктирные линии — отклонения по длине крыши и боковой стенке. Крайние точки (Б,, Б2, С,, С2; b,, b2, с,, с2) указывают на видотипичные пределы амплитуд изменчивости этих параметров при нормальном гнездостроении
Вариабельность инстинктивного поведения особенно убедительно была показана Вагнером на примерах конструктивной деятельности пауков и ласточек (рис. 4). Обстоятельный анализ этих фактов привел его к выводу, что лабильность инстинктивного поведения ограничена четкими видотипичными рамками, что стабильными в пре-
54
делах вида являются не сами инстинктивные действия, а пределы амплитуд их изменчивости. Тем самым Вагнер предвосхитил одно из основных положений современной этологии.
Впоследствии и другие советские ученые разрабатывали вопросы изменчивости инстинктивного поведения и его связи с процессами научения. Академик Л.А.Орбели проанализировал зависимость пластичности поведения животных от степени их зрелорождения (см. ч. II, гл. 3). Советский орнитолог А.Н.Промптов указывал на то, что инстинктивные действия животных (птиц и млекопитающих) всегда включают в себя неотъемлемые, очень трудно отчленяемые, но чрезвычайно существенные ус-ловнорефлекторные компоненты, формирующиеся в процессе онтогенеза. Именно эти компоненты, по Промптову, обусловливают пластичность инстинктивного поведения. С другой стороны, взаимодействие врожденных реакций, детерминированных видотипичными признаками строения, с приобретенными на их основе в течение индивидуальной жизни условными рефлексами дает в итоге видоти-пичные особенности, названные Промптовым «видовым стереотипом поведения».
Е.В.Лукина иллюстрировала эти положения Промптова примерами по пластичности гнездостроительной деятельности воробьиных птиц. Так, молодые самки, гнездящиеся впервые в жизни, вьют гнезда, характерные для своего вида. Однако в необычных условиях этот стереотип заметно нарушается. Так, горихвостка и синица-пухляк, являющиеся дуплогнездниками, устраивают при отсутствии дуплистых деревьев свои гнезда под корнями, а серая мухоловка, вьющая гнезда в укрытиях (расщелинах пней, углубленных стволов, за отставшей корой и т.п.), может в случае необходимости устраивать их и на горизонтальных ветках или даже прямо на земле и т.д.
Как мы видим, все это случаи модификации гнездост-роительного инстинкта, конкретно — в отношении местоположения гнезда. Описано также немало примеров замены гнездостроительного материала: вместо травинок,
55
мха, лишайника подчас используются такие искусственные материалы, как вата, упаковочные стружки, марля, веревки и т.п. Известны даже случаи, когда мухоловки-пеструшки сооружали свои гнезда в парках Москвы почти целиком из трамвайных билетов. Аналогичные данные были получены и в специальных опытах, в которых исследовалась пластичность инстинктивного поведения при замене яиц или птенцов (опыты Промптова, Лукинбй, Скребиц-кого, Вилке).
У насекомых пластичность строительной деятельности (сооружение чехликов у гусениц Psyche viciella) была в 60-х годах обстоятельно изучена в лаборатории польского зоопсихолога Р.И.Войтусяка (исследования К.Громыша). У другого насекомого, гусеницы Autispila stachjanella, там же изучалась пластичность инстинктивного поведения при устройстве ходов в листьях и коконах (исследования М.Бе-рестынской-Вильчек). Ученые обнаружили большую адаптивную вариабельность инстинктивных действий, особенно при починке сооружений этих насекомых, причем в искусственных, экспериментальных условиях эти действия могут заметно отличаться от выполняемых в обычных, естественных условиях.
Промптов был, безусловно, прав, когда подчеркивал значение слияния врожденных и приобретаемых компонентов во всех формах поведения. Вместе с тем его понимание пластичности инстинктов представляет собой шаг назад по сравнению с концепцией Вагнера, доказавшего, что врожденными являются не инстинктивные действия, а те рамки, в пределах которых эти действия могут выполняться в измененном виде в соответствии с данными условиями среды. Факты, приводимые Промптовым, лишь подтверждают это.
Отказав инстинктивным компонентам поведения во всякой собственной вариабельности, Промптов считал, что пластичность обеспечивается лишь условнорефлек-торными компонентами поведенческого акта. В действительности же мы имеем здесь дело с разными по своей величине и своему значению категориями изменчивости
56
поведения. Во-первых, это изменчивость самих врожденных, т.е. инстинктивных, компонентов, которая проявляется в индивидуальной вариабельности видотипичного поведения (в пределах наследственно закрепленной нормы видотипичного реагирования). Примерами такой вариабельности могут служить приведенные Вагнером факты по гнездостроительной деятельности ласточек и упомянутые исследования инстинктивного поведения гусениц. Во-вторых, правда реже, в экстремальных условиях видо-типичное, инстинктивное поведение может довольно сильно видоизменяться. Здесь проявляется роль индивидуального опыта, так как именно он обеспечивает более или менее выраженную приспособляемость к необычным, выходящим за рамки нормы внешним условиям. И наконец, существуют различные индивидуально приобретаемые и поэтому максимально вариабельные формы поведения, в которых главенствующую роль играют уже разные формы научения, имеющие под собой, конечно, тоже инстинктивную основу и переплетающиеся с врожденными компонентами поведения.
Принципиальное значение различий в изменчивости инстинктивного и благоприобретаемого поведения глубоко проанализировал академик А.Н.Северцов, основоположник эволюционной морфологии. В работах «Эволюция и психика» (1922) и «Главные направления эволюционного процесса» (1925) он показал, что у высших животных (млекопитающих) существуют два типа приспособления к изменениям окружающей среды: (I) изменение организации (строения и функций животных), совершающееся весьма медленно и позволяющее приспособиться лишь к очень медленно протекающим постепенным изменениям среды, и (2) изменение поведения животных без изменения их организации на основе высокой пластичности ненаследственных, индивидуально приобретаемых форм поведения. В последнем случае возможно эффективное приспособление к быстрым изменениям среды именно благодаря изменению поведения. В этом случае наибольший успех будут иметь особи с более развитыми психическими способнос-
57
тями, «изобретатели» новых способов поведения, как метафорически выразился Северцов, — словом, животные, способные выработать наиболее гибкие, пластичные навыки и другие высшие формы индивидуального изменчивого поведения. Именно в этом плане Северцов рассматривает значение прогрессивного развития головного мозга в эволюции позвоночных.
Что же касается инстинктивного поведения, то оно из-за своей малой изменчивости (ригидности) не может выполнять такую функцию. Но подобно изменениям строения тела животного изменения врожденного поведения могут служить приспособлением к медленным, постепенным изменениям среды, поскольку они требуют много времени для своего осуществления.
Напомним, что это изменение врожденного поведения совершается в процессе эволюции на основе индивидуальной изменчивости инстинктивного поведения. Именно в этой индивидуальной лабильности видотипичных действий животных Дарвин искал источник зарождения и развития инстинктов.
Северцов подчеркивал, что значение такого постепенного приспособления не менее важно, чем приспособление с помощью изменения индивидуально приобретаемого поведения. «Инстинкты, — писал он, — суть приспособления видовые, полезные для вида в такой же степени, как и те или другие морфологические признаки, и столь же постоянные»23. При этом Северцов обращал внимание на то, что способность к научению, к установлению новых ассоциаций зависит от определенной наследственной высоты психической организации. Сами действия не являются при этом наследственными. В инстинктивном же поведении наследственно закреплено и то и другое. Сегодня мы можем в подтверждение этого тезиса Северцова сказать, что инстинктивное поведение животных определяется врожденной программой действий, реализуемой в ходе накопления индивидуального опыта.
23 Северцов А.Н. Собр. соч. Т. III. M.; Л., 1945. С. 300.
58
Подводя итог изложенному и учитывая современные знания о поведении животных, можно следующим образом охарактеризовать взаимосвязь и взаимозависимость между врожденными и приобретаемыми компонентами поведения и биологическое значение их специфической изменчивости.
Постоянство, ригидность инстинктивных компонентов поведения необходимы для обеспечения сохранности и неуклонного выполнения наиболее жизненно важных функций независимо от случайных, преходящих условий среды, в которых может оказаться тот или иной представитель вида. Во врожденных компонентах поведения хранится итог всего эволюционного пути, пройденного видом. Это квинтэссенция видового опыта, самое ценное, что приобретено в ходе филогенеза для выживания особи и продолжения рода. И эти передаваемые от поколения к поколению обобщенные и генетически фиксированные программы действий не должны и не могут легко изменяться под влиянием случайных, несущественных и непостоянных внешних воздействий. В экстремальных же условиях еще имеются шансы выживания благодаря резервной пластичности инстинктивного поведения в виде модификации.
В остальном реализация врожденной программы поведения в конкретных условиях индивидуального развития животного обеспечивается процессами научения, т.е. индивидуального приспособления врожденного, видотипич-ного поведения к частным условиям среды. Для этого необходима предельная гибкость поведения, но опять же возможность индивидуального приспособления без утраты существенного, накопленного в ходе эволюции вида, требует незыблемой основы в виде устойчивой инстинктивной диспозиции. Только она дает животному способность с зыгодой для себя реагировать в любой ситуации.
Внешние факторы инстинктивного поведения
Когда говорят об автономности внутренних факторов поведения, об их независимости от внешней среды, то необходимо помнить, что эта независимость является лишь относительной. Уже из приведенных опытов Хольста видно, что эндогенная активность не существует ни «сама по себе», ни «сама для себя»: значение этих спонтанных процессов в центральной нервной системе состоит в предис-позиции к возникновению жизненно важных ситуаций («в случае чего — все готово»). В результате животное способно по первому же сигналу незамедлительно и с максимальной пользой для себя реагировать на изменение в окружающей среде.
Эта готовность обеспечивается тем, что соответствующие эндогенные системы периодически активируются как собственной ритмикой, так и внешними воздействиями (например, изменением продолжительности светового дня, повышением или уменьшением температуры и т.д.). Однако инстинктивные движения, по этологической концепции, заблокированы специальной системой «врожденных пусковых механизмов». Последние представляют собой совокупность нейросенсорных систем, обеспечивающих приуроченность поведенческих актов к биологически адекватным условиям среды (к «пусковой ситуации»). Как только животное оказывается в такой ситуации, соответствующий врожденный пусковой механизм обеспечивает распознавание, оценку и интеграцию специфических для данной инстинктивной реакции раздражителей, после чего наступает растормаживание, снятие «блокировки». Очевидно, одновременно происходит активация соответствующих нервных центров и снижение порогов их раздражимости.
Характерной особенностью для врожденных пусковых механизмов является избирательность реагирования на внешние стимулы: они отзываются только на совершенно определенные комбинации раздражителей, которые только и могут вызвать биологически целесообразную реакцию. Иными словами, в сенсорной сфере существует некая «фильтрующая» функция, выражающаяся в специфической преадаптационной «готовности» воспринимать такие раздражители.
Итак, благодаря врожденным пусковым механизмам внутренняя мотивация поведения получает «выход наружу», т.е. создается возможность без индивидуального опыта в биологически значимых ситуациях реагировать так, чтобы это способствовало сохранению особи и вида.
Подводя итог сказанному, можно сказать, что под врожденным пусковым механизмом следует понимать совокупность нейросенсорных систем, обеспечивающих адекватность поведенческих актов по отношению к «пусковой ситуации»: настройку анализаторов на восприятие специфических раздражителей и распознавание последних, интеграцию соответствующих раздражений и растормаживание (или активацию) нервных центров, связанных с данным поведенческим актом.
Внешние раздражители, составляющие в своей совокупности пусковую ситуацию, получили название «ключевых раздражителей», поскольку они подходят к своим врожденным пусковым механизмам, как ключ к замку. Ключевые раздражители являются такими признаками компонентов среды, на которые животные реагируют независимо от индивидуального опыта врожденными, видр-типичными формами поведения, точнее, определенными инстинктивными движениями. В описанном поведении
68
курицы это будут определенные общие признаки, свойственные всем ее наземным врагам.
Кроме собственно растормаживающих ключевых раздражителей (их называют также «пусковыми раздражителями») различают также настраивающие ключевые раздражители, предварительно понижающие порог раздражимости нервных центров, причастных к данным действиям животного, а также направляющие ключевые раздражители, о которых пойдет речь при обсуждении таксисов. Общим свойством всех ключевых раздражителей является то, что это специфические элементарные признаки жизненно важных компонентов среды. Ключевыми раздражителями являются простые физические или химические признаки («просто» форма, размер, подвижность, цвет, запах и т.д.), или их пространственные отношения (взаиморасположение деталей, относительная величина и т.д.), или же векторы. Носителями этих признаков могут быть как другие животные, так и растения и объекты неживой природы. В последнем случае ключевые раздражители выполняют преимущественно направляющую функцию. Так, например, немецкий этолог Ф.Вальтер показал, что у детенышей антилоп ключевым стимулом, определяющим выбор места отдыха (лежа, неподвижно), является «что-то вертикальное» вне зависимости от того, что конкретно это за объект.
Наилучшим образом изучены ключевые раздражители, носителями которых являются животные. Эти раздражители представляют особый интерес и потому, что являются первичными, генетически фиксированными элементами общения у животных. Так, в ставших уже классическими опытах голландского зоолога Н.Тинбергена, одного из основоположников современной этологии, с помощью макетов изучалась пищевая реакция («попрошайничество») птенцов серебристых чаек (клевание клюва родительской ороби) и дроздов (вытягивание шеи и раскрытие клюва) при появлении родительской особи.
В естественных условиях голодный птенец серебристой чайки клюет красное пятно на желтом клюве родителя, и
69
тот в ответ отрыгивает пищу в рот птенцу. В опытах предъявлялась серия все более упрощаемых моделей — макетов. Первая модель точно воспроизводила внешний облик естественного носителя ключевых раздражителей, т.е. головы взрослой серебристой чайки с желтым клювом и красным пятном на нем. В последующих моделях путем проб постепенно исключались отдельные детали, и в результате макет становился все менее похожим на голову птицы (рис. 7). В конце концов остался лишь плоский красный предмет с продолговатым выступом. Но этот предмет оказался способным вызвать даже более сильную реакцию птенцов, чем исходная модель. Еще более эта реакция может быть усилена, если этот макет заменить тонкой белой палочкой, исчерченной поперечными темно-красными полосами. Ключевыми раздражителями в данном примере будут просто «красное» и «продолговатое».
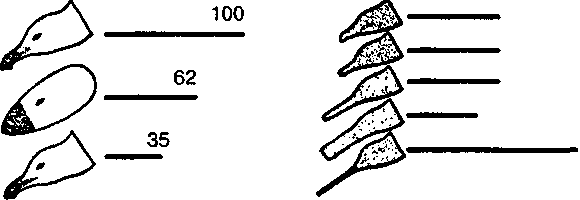
Рис. 7. Опыты Тинбергена с применением макетов головы серебристой чайки. Линиями обозначена интенсивность реакций птенцов на макет (число клеваний)
В опытах, проведенных Тинбергеном с десятидневными птенцами дроздов, выявилась другая категория ключевых раздражителей — взаиморасположение и относительная величина деталей объекта-носителя. В качестве макетов использовались плоские диски. Если показать птенцам такой круг, они будут тянуться к его верхней части (местонахождению головы птиц). Если же к большому кругу прибавить маленький, птенцы потянутся к нему. Если, наконец, прибавить к большому кругу два меньших круга, но разных размеров, то решающее значение приобретет
70

Рис. 8. Опыты Тинбергена с птенцами дроздов
относительная величина этих фигур. Размеры «головы» не лолжны приближаться к размерам «туловища»: при большом «туловище» птенцы потянутся к большему дополнительному кругу («голове») (рис. 8), при маленьком же — к меньшему.
Интересные опыты проводил еще в середине 30-х годов Г.Л.Скребицкий совместно с Т.И.Бибиковой на подмосковном озере Киёво, изучая отношение чайки к ее яйцам. Яйца перекладывались из гнезда в гнездо, заменялись яйцами других видов, ис->£ кусственными яйцами, а затем и раз-
Лш личными посторонними предметами
J/F различной величины, формы и окрас-
^ ки. Оказалось, что чайки садились как
на чужие, так и на деревянные, стек-J лянные, каменные, глиняные яйца
Ч^^Ыф различной величины и самой разнооб-^^^■^ разной окраски и начинали их «выси-^Щ0 живать». То же самое наблюдалось,
когда вместо яиц в гнездо подклады-вались разноцветные шары, камешки или картофелины. По свидетельству экспериментаторов, «чайки, сидящие на таких предметах, представляли очень оригинальную картину, но в особенности необычайным становилось зрелище, когда согнанная с гнезда птица возвращалась к нему обратно и, прежде чем сесть, заботливо поправляла клювом разноцветные шары, камешки или картошку»1.
Если, однако, чайкам подкладывались предметы иной формы, например кубики или камни с неровными края-
1 Скребицкий Г. Отношение чайки к гнезду, яйцам и птенцам // Рефлексы, инстинкты и навыки. М., 1936. С. 144—145.
71
ми, поведение птиц заметно изменялось. Когда за край гнезда клалось по одному яйцу и инородному предмету, птицы вкатывали обратно в гнездо наряду с яйцами лишь округлые гладкие предметы, размеры которых соответствовали размерам яиц. В известных пределах не препятствовали этому существенные отклонения от нормы в весе (каменное яйцо весило в два с лишним раза больше, чем чаечье), материал, из которого был изготовлен предмет, и окраска.
Исследователи пришли к выводу, что положительная реакция чайки на яйцо определяется лишь несколькими его элементарными признаками: округлостью, отсутствием выступов, углублений или насечек (рис. 9). Именно эти признаки и выступали здесь как ключевые раздражители.
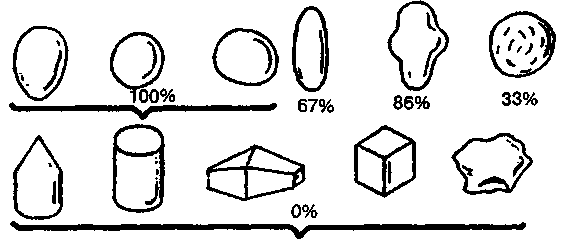
Рис. 9. Вкатывание в гнездо различных предметов в зависимости от их свойств у обыкновенной чайки (в процентах). Верхний ряд, слева направо: яйца (ча-ечьи, куриные, деревянные и каменные), деревянные шары, другие округлые предметы (картошка, камни) удлиненное глиняное яйцо с углублениями, глиняный шар с зарубками. Нижний ряд, слева направо: деревянный конус, деревянный цилиндр, деревянные и глиняные многоугольники, деревянные и глиняные кубики, угловатые камни (по Скребицкому и Бибиковой)
Важная особенность действия ключевых раздражите^ лей заключается в том, что они подчиняются закону сум-мации: с увеличением их параметров пропорционально
72
усиливается инстинктивная реакция животного. В экспериментальных условиях это может привести к так называемым «супероптимальным» реакциям, когда животное «преувеличенно», сильнее, чем в норме, реагирует на искусственный раздражитель, в котором «сгущены краски». Мы уже встречались с этим явлением при описании опыта с птенцами чайки, когда они сильнее реагировали на поперечно исчерченную красными полосками палочку, чем на настоящий клюв живой птицы. Суммация проистекает здесь из многократности красных меток и их большей контрастности.
Эффект супероптимальной реакции может в условиях эксперимента привести даже к биологически абсурдному поведению животного. Если, например, предложить чайке два яйца разной величины, она закатит в гнездо более крупное. В итоге может возникнуть такое положение, что птица бросит свое яйцо, чтобы попытаться высиживать деревянный макет яйца гигантских размеров, обладающих супероптимальными признаками ключевого раздражителя.
Как видно из приведенных примеров, ключевые стимулы действуют на поведение животного принудительно, заставляя его выполнять определенные инстинктивные движения, невзирая на возможно воспринимаемую животным общую ситуацию. Это объясняет многие, ранее казавшиеся загадочными моменты в поведении животных.
Так, например, еще в начале нашего века один из авторов антропоморфических сочинений по психологии животных Т.Целл дал следующий ответ на вопрос о том, почему крупные хищники в нормальных условиях при встрече с человеком не нападают на него: например, лев не нападает не потому, что почитает человека, а потому, что не уверен в исходе схватки. «Есть ли у человека оружие, да какое оно, это оружие? — думает лев. — Да ну его, пойду-ка я лучше своей дорогой».
Конечно, лев, как и другие крупные хищники, как правило, уклоняется от нападения на человека не потому, что руководствуется такими глубокомысленными рассуждениями. Разгадка «почтительного» отношения диких зве-
73
рей к человеку, очевидно, кроется в следующем. Будучи сытым, хищник не реагирует и на присутствие животных, которыми обычно питается. У голодного же хищника преследование животных-жертв и нападение на них обусловливаются сочетанием рассмотренных выше внутренних факторов (первопричина — изменение уравновешенности внутренней среды организма в результате недостатка питательных веществ) с соответствующими внешними стимулами — ключевыми раздражителями, носителями которых являются естественные объекты питания, т.е. животные-жертвы, но не человек.
Даже самый кровожадный хищник не волен нападгть на кого угодно и когда угодно. Эти действия также ориентируются во времени и пространстве ключевыми раздражителями, как и все прочие поведенческие акты. Другими словами, дело не в том, «хочет» или «не хочет» животное поступить так или иначе. Если внутреннее состояние животного соответствует определенной внешней пусковой ситуации, то оно волей-неволей вынуждено вести себя так, как это диктует для данных условий генетически зафиксированный код видотипичного поведения.
44. Инстинктивное поведение животных. Развитие представлений о взаимосвязи инстинкта и научения в истории зоопсихологии.
Древние уже задумывались над тем, есть ли у животных душа. Платон считал, что всё поведение животных определяется влечениями. Аристотель считал, что души животных смертны, в отличие от человеческих душ, но всем живородящим животным с красной кровью присущи те же 5 чувств, что и человеку. На основании наблюдений Аристотель обосновал представление об индивидуальном приобретении определённых компонентов поведения. У стоиков впервые появляется понятие инстинкта – прирождённое, целеустремлённое влечение, направляющее животного на приятное, полезное и уводящее его от опасного, вредного. По Хризиппу, инстинктивное поведение осуществляется животными без участия разума (которого у животных нет), без понимания смысла своей деятельности, на основе врождённых знаний. Сенека указывал на однообразие форм и результатов прирождённого поведения животных. Т.о. уже во времена античности и средневековья уже поднимались проблемы врождённого и приобретённого, инстинкта и научения, обсуждалась роль внешних и внутренних факторов психической активности животных.
К концу 18 века – рост интереса к психике животных. Ламеттри считал инстинкты совокупностью движений, выполняемых вынужденно, вне зависимости от размышлений и опыта; применял сравнительный подход к изучению психической деятельности животных. Показал прогрессивное усложнение психических способностей по направлению к человеку. Кондильяк считал, что инстинкты произошли из разумных действий путём постепенного выключения сознания. Леруа возражал и высказывал мысль о происхождении человеческого разума от инстинкта животных. Бюффон высказал мысль о приспособительном значении психики и о том, что психические качества животных не менее важны, чем их физические качества. Т.о. в результате пришли к выводу, что сложные действия животных являются результатом сочетания врождённых природных функций, доставляющих животному удовольствие, и привычек.
По мере развития эволюционной теории вопросом о взаимодействии врождённого и приобретённого у животных интересуются всё больше. Ламарк считал инстинкт наклонностью, возникшей на основе потребностей, принуждающей к выполнению действий без всякого участия мысли и воли. Инстинкты возникли в процессе эволюции в результате длительных воздействий на организм определённых агентов среды. Потребности порождают привычку, привычка переходит в инстинкт. Генетическая передаваемость привычек и инстинктов. То есть Ламарк видел в инстинктах не какую-то сверхъестественную силу, а естественные реакции организма на воздействия среды, сформировавшиеся в процессе эволюции. Рулье видел первопричину происхождения психических качеств и особенностей животных во взаимодействии организма с условиями жизни. Инстинкт – это сформировавшаяся на протяжении истории вида, выработанная условиями наследственная реакция на определённые воздействия среды. В индивидуальной жизни инстинкты могут изменяться в результате накопления опыта. Надо понимать, что под “инстинктом” понимали самые разнообразные проявления животных. Дарвин много внимания уделил проблеме инстинкта. Инстинкт – акт животного, который выполняется им без предварительного опыта или одинаково многими особями, без знания с их стороны цели, с которой он производится. Происхождение инстинктов Дарвин объяснял естественным отбором. Прогресс умственных способностей Дарвин объясняет тем, что отдельные части мозга мало-помалу утратили способность отвечать на ощущения инстинктивным способом.
45. Сравнительное изучение раннего онтогенеза детей и детенышей человекообразных обезьян.
Локомоторные движения у шимпанзе развиваются быстрее, чем у ребёнка, но в той же последовательности (поднимание головы 3мес/3мес; самостоятельное хождение 64мес/25 мес). Но при этом ребёнок выхаживается родителями, а шимпанзе рождается в стае. Важно воспитывать: наследника одного из немецких князей Каспара Хаузера с детства до 17 лет держали в темнице и с ним никто не разговаривал; в результате на свободе он с трудом ходил и сумел сказать только 1 фразу.
Были проведены эксперименты по воспитанию шимпанзе в домашних условиях. Супруги Келлог воспитывали шимпанзе Гуа совместно с сыном. Дети вместе играли. Шимпанзе хуже научалась подражать ранее неизвестным движениям. Дети дружили. Гуа любила играть с детьми, но не любила, когда на неё показывали пальцем и смеялись (проявляла агрессию). Смех – с 8 месяцев. Эмоциональные выражения радости, облегчения. Кашель, смех, храп ночью . Гуа неспособна произнести “папа”. Вначале Гуа лучше понимала слова, потом Дональд (сын) перегнал её. Научили носить одежду, чистить зубы, включать свет, сидеть на стуле.
Ладыгина-Котс взяла на 2 года полуторагодовалого шимпанзе Иони. Игры, в основном деструктивного характера. Экспериментальные игры (с водой). Собирание предметов и объединение их по разным признакам. Разглядывание мира через цветную клеёнку. Мучил маленьких животных, не любил детей, тараканов давил рукой через тряпку. Когда рядом никого не было, практически не играл – нужна компания. Тоже эмоции радости. Ладыгина-Котс позже наблюдала за своим сыном и отметила сходство изменений мимики. Иони понимал ряд предложений типа “иди в клетку”, “иди ко мне”, “горячо”. Воспитание шимпанзе сводится к выработке условных рефлексов и навыков. Подражание. Рисование на бумаге не продвигается дальше линий.
46. Групповое поведение животных и проблема зарождения общественных отношений.
Групповое поведение зарождается уже у насекомых. Но в отличие от строгой иерархии насекомых, у человекообразных обезьян есть возможность прижизненной смены ролей (здесь разумно привести пример с канистрами, которыми стучал субдоминантный самец, чтобы завоевать авторитет, и как свергнутый таким образом доминантный самец выкрал эти канистры и путём такого же грохотания вернул себе былую славу). Дружеские связи играют важную роль в определении доминантности. Ритуализированная агрессия: причина появления мимики у обезьян. Пристальный взгляд – первое мимическое выражение.
Формы сотрудничества в стаде обезьян:
Взаимное обслуживание. Очищение кожных покровов и придатков, реже – глаз и зубов. Врождённость рефлекса обыскивания. Обыскивание – признак контакта между двумя обезьянами. Демонстрация дружбы.
Взаимопомощь и сотрудничество на базе инстинкта самосохранения. Заступничество, совместное выступление против отдельного животного (в т.ч. вожака), помощь матери в уходе за детёнышем, сотрудничество в угрозе внешнему врагу.
Сотрудничество обезьян в экспериментальных условиях.
Стадное стимулирование и подражание. Стадное стимулирование – активизация обычных для данной ситуации оборонительных или пищевых рефлексов; низшая стадия развития подражания, свойственная и более низшим животным: если одна лягушка прыгает в озеро, остальные тоже прыгают. Три формы подражания, по Войтонису: подражание простому действию, непосредственно приводящего к получению желаемого объекта (при прыгании, ударах, за которыми следует подкрепление); внешнее воспроизведение приёмов без внимания к связям и отношениям; повторение приёма с вниманием к результатам действий и пониманием их. Последняя форма – для передачи умений и навыков. Ладыгина-Котс считает, что шимпанзе может изменить своё поведение в соответствии с поведением близких к нему людей (Иони научился плеваться, вытирать нос тряпочкой, умываться и т.п.)
Особенности средств стадной коммуникации в стаде обезьян:
Активное побуждение одного животного другим к какому-либо действию. Побуждение к следованию, обыскиванию, нападению, сидению и проч.
Направленность, адресованность средств общения обезьян.
Способность к двойственным поступкам (см. в обманывании)
Различия между групповым поведением насекомых и обезьян:
Пластичная иерархия и прижизненная смена ролей у обезьян
Ролевые различия на физиологическом уровне у муравьёв
Важность дружеских связей у обезьян, факультативное научение, социальные традиции
У насекомых научение облигатное
Забота о потомстве у обезьян: помощь в выращивании чужого ребёнка, табу на ребёнка. Самцы при матери до 18 лет, самки пожизненно.
Забота о потомстве у насекомых: кормление личинок, проветривание, очистка помещений. Нет пожизненного общения с родителем и передачи опыта.
Ритуализированная агрессия у обезьян как признак высокого уровня группового поведения.