

А тф цАмф

 Протеникиназа
Гликогенсинтаза «b»
Протеникиназа
Гликогенсинтаза «b»
 фосфорилирование
фосфорилирование

C а++ Киназа фосфорилазы Гликогенсинтаза «а»

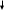
УДФ-глюкоза Гликоген
Ф
 осфорилаза
«b»
Фосфорилаза «а»
осфорилаза
«b»
Фосфорилаза «а»

Фосфоролиз гликогена гликогена
УДФ-глюкоза
 Глюкоза-1-фосфат
Глюкоза-1-фосфат
Глюкоза-1-фосфат
Глюкоза-1-фосфат
Глюкоза-6-фосфат Глюкоза-6-фосфат

СО2+ Н2О Глюкоза
Гликолиз.
Под гликолизом в общем смысле принято понимать разложение углеводов. В зависимости от конкретного организма и (или) условий его роста, гликолиз может идти по анаэробному пути ( в этом случае он так и называется –анаэробный гликолиз ).
И может идти по аэробному пути ( соответственно – аэробный гликолиз ). Для многих микроарганизмов, растущих в анаэробных условиях ( в отсутствие кислорода ) гликолиз является основным катаболическим путем для получения энергии из углеводных субстратов, приводя к образованию определенных конечных продуктов метаболизма, например, этанола, лактата, глицерина. Процесс такого типа часто называют брожением. У животных анаэробный гликолиз ( с образованием лактата ) обеспечивает энергией сокращение скелетных мышц в условиях ограниченного снабжения кислородом. В некоторых случиях гликолиз ( аэробный гликолиз ) идет по анаэробному пути в аэробных условиях т. е. конечным продуктом является лактат ), хотя кислород и присутствует . Такие процес-сы идут, например, в эритроциттах, сетчатке, слизистой кишечника, в тканях плода (вскоре после рождения ). Гликолиз –единственный анаэробный путь, дающий энергию. В аэробных условиях ( в присутствии кислорода ) реакции гликолиза составляют начальную форму разложения углеводов,связанную далее с циклом лимонной кислоты. В этом случае гликолиз останавливается на стАДНи образования ПВК. В аэробных условиях гликолиз и цикл Кребса совместно приводят к полному окислению гексозы до углекислого газа. Этот процесс сопровождается выделением больших количеств потенциально доступной метаболической энергии, в основном в виде высокоэнергетических продуктов НАДН и ФАДН2 ,которые, окисляясь в ходе дыхательной цепочки, дают энергию, необходимую для процесса ОФ , в результате которого образуется АТФ. Гликолиз –это первая до конца расшифрованная последовательность биохимических реакций, для этого потребовалось почти 50 лет ( 1890-1940). За это время были открыты фосфоэфиры , АТФ, НАД ,выделены и охарактеризованы многие факторы и т.д.
Гликолиз протекает в цитозоле .
Схема гликолиза :
Очень важно образование на конечном этапе НАД+ ,который используется для окислительного фосфорилирования 3-фосфоглицеринового альдегида.
Лактат является конечным продуктом гликолиза. Т. к. лактат (как, впрочем, и пирува) достаточно легко проходит через плазматическую мембрану, он попадает из мышечной клетки (или эритроцитов) в кровь, с током крови попадает в печень, где окисляется в пируват , который в печени затем превращается в глюкозу. Таким образом образуется своего рода цикл. Эти превращения и составляют цикл Кори :
Вообще же в ходе гликолиза образуется 4 молекулы АТФ ,а расходуется 2 АТФ. Таким образом, суммарный эффект окисления одной молекулы глюкозы в ходе гликолиза до лактата равен 2 АТФ. Утилизируется всего около 3% всей возможной энергии , которую можно было бы получать при окислении глюкозы, т.е. в сутки нужно было бы расщепить 27 кг глюкозы , чтобы обеспечить ортанизм энергией ( если бы энергия получалась бы только путем гликолиза ).
Однако ,гликолиз имеет место в клетках. Он нужен в мышцах для быстрого получения энергии. А, например, эритророциты только за счет гликолиза и могут получать энергию , т. к. у них нет митохондрий , и, следовательно , нет и окислительного фосфорилирования . Большинство простых сахаров можно расщепить гликолитическим путем. Все, что для этого необходимо – это превратить эти сахара в Г-1- Ф, Г-6-Ф, Ф-6-Ф. Это происходит в ходе взаимного превращения моносахаров.
Аэробное окисление глюкозы
В аэробных условиях (в присутствии кислорода ) реакции гликолиза составляет начальную фазу расщипления углеводов, связанную дальше с другим важным циклом- циклом лимонной кислоты (или его называют цикл трикарбоновых кислот, или цикл Кребса ). В этом случае гликолиз останавливается на стАДНи образования пирувата, который затем путем окислительного декарбоксилирования превращается в ацетил- КоА, а ацетил-КоА , вступая в цикл Кребса, окисляется до СО2 и Н2О. Образующиеся в ходе цикла Кребса и в ходе окислительного декарбоксилирования восстановленные формы НАДН и ФАДН2 поступают в дыхательную цепь, где энергия, образующаяся при переносе с них Н+ и е- на О2 может быть использована , в ходе окислительного фосфорилирования, для синтеза АТФ. Легко рассчитать энергетический эффект этого процесса. На стАДНи образования пирувата из глюкозы образуются 2 молекулы АТФ и 2 молекулы НАДН2. На стАДНи образования ацетил-КоА из пирувата ( в ходе окислительного декарбоксилирования пирувата ) образуется 2НАДН2 ( т.к. из глюкозы получается 2 молекулы пирувата ). В ходе цикла Кребса образуется 1 моль ГТФ (=АТФ ), 3 молекулы НАДН2 и 1 молекула ФАДН2 ( все это умножает на 2, т. к. в цикл Кребса вступает 2 молекулы ацетил-КоА ). Итого получается 2 молекулы АТФ + 2 молекулы ГТФ (=АТФ) + 10 молекул (2+2+6 ) НАДН и 2 моле-кулы ФАДН2. Одна НАДН при окислении в дыхательной цепи может дать энергию достаточную для синтеза 3-х АТФ, следовательно , 10 НАДН2 дадут 30 АТФ. Молекула ФАДН2, окисляясь в дыхательной цепи, может дать энергию, достаточную для синтеза 2 АТФ, а 2ФАДН2=4АТФ. Итого, получается 38 молекул АТФ. Всего при окислении одной молекулы глюкозы выделяется около 600 ккал. При аэробном окислении глюкозы , как мы подсчитали, образуется 38 молекул АТФ. АТФ, как известно , при гидролизе до АДФ дает = 10 ккал, т.е. всего на синтез АТФ при аэробном окислении затрачивается 380 ккал, что составляет около 60%,т.е. КПД аэробного окисления =60%, в то время, как КПД гликолиза составляет около 3 % ( всего 2 молекулы АТФ ) .
Гликолиз протекает в цитозоле. Мы уже говорили о том, что ПВК относительно легко проходит через мембраны ( когда говорим о судьбе лактата ). Так и происходит , ведь все последующие этапы окисления глюкозы проходят в митохондриях. На начальных этапах окисления глюкозы (а они проходят в цитозоле) для окислительного фосфорилирования 3-фосфоглицеринового альдегида необходима окисленная форма НАД+. В ходе гликолиза она получается при восстановлении ПВК в лактат .При аэробном окислении глюкозы первый этап заканчивается образованием ПВК, которая через мембрану поступает внутрь митохондрий, подвергается окислительному декарбоксилированию, далее- цикл Кребса и дыхательная цепь. Можно было бы НАДН, образующуюся при фосфорилировании 3-фосфоглицеринового альдегида (т.к. мы не можем ее окислить с помощью ЛДГ- реакции до НАД+), отправить в митохондрии, там в дыхаательной цепи окислить до НАД+ и использовать для фосфорилирования 3-фосфоглицеринового альдегида. Но ,оказалось, что НАДН и НАД+ не проходят через мембраны. Здесь действуют челночные механизмы: за счет НАДН образуется какая-то восстановленная форма вещества , которая проходит через мембраны, внутрь митохондрий, а окисленная форма возвращается в цитозоль, т.е. это вещество является , по сути, проводником протонов (переносчиком). При этом НАДН Окисляется до НАД+, что и требовалось. Так работает глицерофосфатный челночный механизм:
Так же работает и малатный (яблочная кислота (или малат)-ЩУК) челночный механизм. Мы уже говорили, что большие количества АТФ угнетают ключевые ферменты гликолиза. Т. к. аэробный распад глюкозы дает большой выход АТФ,. то включение этого процесса подавляет процесс распада углеводов в лактат (это эффект Пастера), т.е. гликолиз ингибируется при поступлении кислорода. Кроме того, что при накоплении АТФ ингибируются ферменты гликолиза (гексокиназа, фосфофруктокиназы и др.), есть и другие механизмы подавления гликолиза при поступлении кислорода. Например, в процессе гликолиза для образования АТФ путем фосфорилирования АДФ, который нужен и при дыхании (для окислительного фосфорилирования),т.е. возникает конкуренция, в которой выигрывают митохондрии. Для образования молочной кислоты из пирувата нужен НАДН, который также нужен и для работы челночных механизмов в аэробных условиях (для восстановления фосфодиоксиацетона до глицерофосфата), т.е.снова конкуренция и снова выигрывают митохондрии
Таким образом в аэробных условиях гликолиз не идет, хотя возможен (например, в опухолях, где угнетен фермент глицерофосфат-ДГ, который катализирует восстановление фосфодиоксиацетона до глицерофосфата- челноки не работают, идет гликолиз.
В скелетных мышцах, где снабжение кислородом невелико и метаболические процессы в большей степени протекают по анаэробному пути, АТФ поступает в результате гликолиза, так что основным субстратом служат запасы гликогена, а конечным продуктом – лактат. В других мышечных тканях, способных к аэробному метаболизму в жестких условиях ( например, в сердечной мышце ) в основном одновременно идет гликолиз с образованием пирувата и действует цикл лимонной кислоты . В этом случае основная часть необходимого АТФ доставляется благодаря процессу ОФ в митохондриях. В соответствии с такими функциональными различиями между аэробным и анаэробными мышечными тканями сердечная мышца обычно содержит гораздо больше митохондрий, чем клетки скелетных мышц. Около 40% Сухой массы сердца приходится на митохондрии.
В периоды длительной мышечной активности АТФ используется непосредственно после его образования. В периоды отдыха энергия АТФ сохраняется в виде макророэрга – креатинфосфата, который в короткие периоды активности и в начальной фазе длительной активности служит донором фосфата для синтеза АТФ из АДФ.
Пентозо-фосфатный путь распада углеводов.
Большинство аэробных организмов используют для полного окисления углеводов до СО2 и Н2О аэробный путь распада. Но многие из аэробов способны также осуществлять и иные метаболические реакции. Один из таких путей, обнаруженный в 50-е годы, включает превращение глюкозо-6-фосфата в СО2 и рибулозо-5-фосфат, т.е. производное пентозы. Поэтому, весь путь целиком называют пентозо-фосфатным.Поскольку глюкозо-6-фосфат может включаться и в гликолиз, и в пентозо-фосфатный путь, его иногда называют гексозомонофосфатным шунтом. Этот путь широко распространен ( животные, растения, бактерии ) и отвечает за многие важные метаболические функции. Пентозофосфатный путь протекает, как и гликолиз, в цитоплазме. Более того, оба пути (и гликолиз и пентозофосфатный ) могут реализовываться в клетке одновременно.
Для изучения пентозофосфатного пути целесообразно рассматривать окисление сразу 3-х молекул глюкозы.
Т.е. всего 8 реакций, из которых первые три- окислительные:
Окисление глюкозо-6-фосфаьа до лактона.
Гидролиз.
Окислительное декарбоксилирование.
Изомеризация кетозы в альдозу.
Обращение конфигурации при С3.
Перенос двууглеродного фрагмента от кетозы к альдозе.
Перенос трёхуглеродного фрагмента от кетозы к альдозе.
Перенос двухуглеродного фрагмента от кетозы к альдозе.
Значение кентозо-фосфатного пути: