

Потенциал действия
При совершении работы, то есть, находясь в состоянии функциональной активности, электровозбудимые клетки быстро теряют потенциал покоя, или говоря иными словами, теряют гомеостатическую «поляризацию» клеточной мембраны. Трансмембранным методом после действия стимула регистрируется резкое изменение заряда клеточной мембраны. Это может быть полная инверсия заряда: в точке приложения стимула мембрана снаружи приобретает отрицательный заряд, а внутри положительный. Такая потеря нормальной поляризации получила название деполяризации (лат. приставка de- означает отсутствие, отмену, устранение чего либо). В этом случае результатом действия стимула будут процессы возбуждения, развивающиеся на мембране нейрона. Однако в результате действия стимула может возникнуть не деполяризация, а наоборот – усиление поляризации клеточной мембраны. Заряд наружной поверхности становится еще более положительным, а внутренней – еще более отрицательным. Эти процессы в электрофизиологии получили название гиперполяризации (от греч. hyper – над, сверх, превышающий норму), соответствуют повышению электрического сопротивления мембраны и, следовательно, развитию процессов торможения. Экспериментальные исследования убедительно показали, что состояния деполяризации и гиперполяризации обусловлены изменением проницаемости клеточных мембран для ионов натрия и калия.
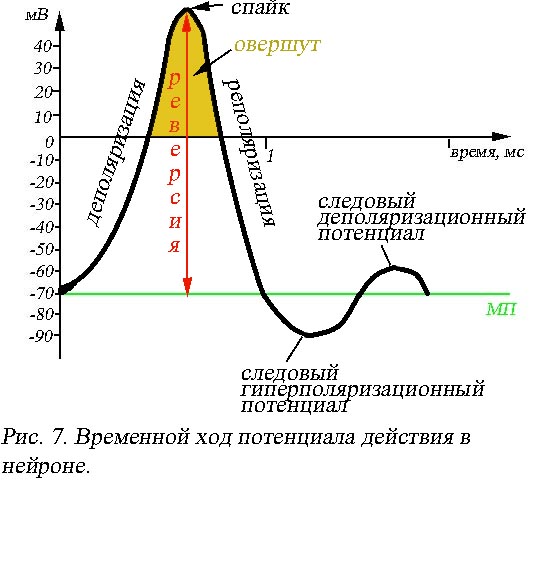 Деполяризация
клеточной мембраны (рис. 7) сопровождается
быстрым сдвигом мембранного потенциала
в положительном направлении, соответствует
на графике фазе нарастания и длится
около 0,2-0,5 мс. Верхняя точка фазы
нарастания лежит в области положительных
значений, соответствует амплитуде
потенциала действия, является фиксированной
для данного нейрона величиной и называется
спайк (пик). Разность потенциалов,
на которую произошла перезарядка
мембраны (от уровня МП до спайка) именуется
реверсией. Участки кривой, лежащие
в области положительных значений оси
ординат, соответствуют инверсии заряда,
- мембрана снаружи становится заряженной
отрицательно, изнутри положительно, -
называют овершут (перелет). Минуя
спайк, график переходит в фазу убывания,
которой соответствуют процессы
реполяризации (лат. re –
снова, заново) клеточной мембраны,
в течение этой фазы происходит
восстановление исходного мембранного
потенциала покоя. Последний участок
реполяризации часто бывает пологим,
например, в мышечных клетках длительность
этой фазы может достигать 0,5-1,5 мс. В
нейронах кроме потенциала действия
можно наблюдать и следовые потенциалы.
Следовый гиперполяризационный
потенциал – участок кривой, расположенный
ниже значений потенциала покоя. Следовый
деполяризационный потенциал –
является результатом незначительной
затухающей деполяризации клеточной
поверхности.
Деполяризация
клеточной мембраны (рис. 7) сопровождается
быстрым сдвигом мембранного потенциала
в положительном направлении, соответствует
на графике фазе нарастания и длится
около 0,2-0,5 мс. Верхняя точка фазы
нарастания лежит в области положительных
значений, соответствует амплитуде
потенциала действия, является фиксированной
для данного нейрона величиной и называется
спайк (пик). Разность потенциалов,
на которую произошла перезарядка
мембраны (от уровня МП до спайка) именуется
реверсией. Участки кривой, лежащие
в области положительных значений оси
ординат, соответствуют инверсии заряда,
- мембрана снаружи становится заряженной
отрицательно, изнутри положительно, -
называют овершут (перелет). Минуя
спайк, график переходит в фазу убывания,
которой соответствуют процессы
реполяризации (лат. re –
снова, заново) клеточной мембраны,
в течение этой фазы происходит
восстановление исходного мембранного
потенциала покоя. Последний участок
реполяризации часто бывает пологим,
например, в мышечных клетках длительность
этой фазы может достигать 0,5-1,5 мс. В
нейронах кроме потенциала действия
можно наблюдать и следовые потенциалы.
Следовый гиперполяризационный
потенциал – участок кривой, расположенный
ниже значений потенциала покоя. Следовый
деполяризационный потенциал –
является результатом незначительной
затухающей деполяризации клеточной
поверхности.
Что бы ответить на вопрос: «какова причина столь резких изменений возникающих на клеточной мембране под действием стимула?» необходимо проследить кинетику ионных токов через мембрану при развитии ПД.
Под действием стимула открываются потенциалзависимые Na+-каналы. Напомним, что в состоянии покоя вход Na+ в клетку конкурентно блокируется катионами кальция. Если сила раздражителя меньше пороговой величины, то потенциал действия не возникает, но развиваются локальные (подпороговые) потенциалы. Если сила раздражителя менее 50-75% пороговой величины, то проницаемость мембраны для ионов изменяется неспецифично и незначительно. Если сила раздражителя достигает и/или превышает 50-75% от порога раздражимости, то начинает преобладать Na+ проницаемость (рис. 8). Под действием стимула мембранные натриевые каналы освобождаются от Са2+, что приводит к изменению мембранного потенциала. Следует отметить, что высокие концентрации наружного Са2+ стабилизируют потенциал покоя, деполяризация в этом случае развивается с задержкой до 35 мс. Напротив, снижение концентрации наружного Са2+, способствует развитию быстрой деполяризации мембраны. Низкие концентрации наружного Na+ приводят к невозможности развития ПД даже при действии стимула.
Графически порог раздражимости клетки отражает критический уровень деполяризации – КУД – это разность потенциалов, которая должна быть достигнута, чтобы местные изменения перешли в распространяющиеся. Иными словами разность потенциалов между МП и КУД отражает возбудимость ткани. В нейронах эта величина практически постоянная и равна -40-(-50) мВ.
Локальные потенциалы (на рис. 8 обозначены
красным цветом) могут суммироваться в
том случае, если они превышают 50-75% от
КУД. Известны два вида суммации:
пространственная и временная. При
пространственной суммации потенциал
действия возникает в результате
одновременного пространственно
разобщенного действия некоторого числа
подпороговых стимулов. Временная
суммация – процесс развития потенциала
действия после серии подпороговых
стимулов, приложенных к одной точке
мембраны. Если локальные потенциалы
ниже 5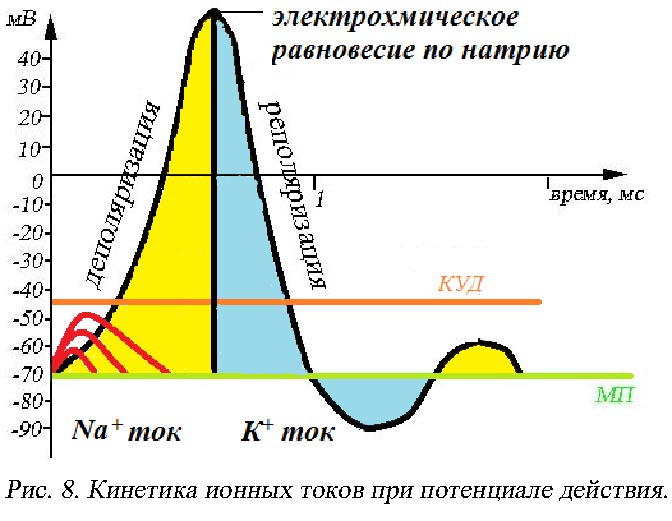 0-75%
от КУД, то они не суммируются ни при
каких условиях.
0-75%
от КУД, то они не суммируются ни при
каких условиях.
При преодолении КУД деполяризация развивается лавинообразно, т.к. все потенциалзависимые m-ворота натриевых каналов в точке действия стимула открываются для вхождения Na+ внутрь клетки. Открытие Na+-каналов является самоподкрепляющимся регенеративным процессом: развивающаяся деполяризация мембраны вызывает дополнительное увеличение ее проницаемости для Na+. Именно поэтому фаза деполяризации характеризуется как Na+-ток, направленный внутрь клетки.
В тот момент, когда амплитуда потенциала действия достигает максимального значения, происходит закрытие h-ворот (инактивационных) Na+-каналов, которые наиболее чувствительны к величине внутриклеточного положительного заряда и поступление Na+ в клетку прекращается. Точка спайка достигается при установлении электрохимического равновесия по Na+ с обеих сторон клеточной мембраны.
Одновременно с наступлением электрохимического равновесия по Na+ происходит повышение проводимости для К+. В мембране открываются K+-каналы, имеющие только одни ворота (активационные), закрытые в покое. Экспериментально установлено, что если при проведении потенциала действия искусственно снизить проводимость мембраны для К+ (этого можно добиться вводя препарат тетраэтиламмоний), то реполяризация развивается очень медленно и очень полого. Следовательно фаза реполяризации характеризуется К+-током, направленным за пределы клетки.
Физиологический смысл реполяризации состоит в восстановлении возбудимости клеточной мембраны. Это возможно только при возвращении состояния потенциала покоя. Для этого необходимо «откачать» катионы натрия и калия против градиента концентрации, что и происходит в результате активации натрий-каливого насоса. При реполяризации происходит накопление ионов натрия на наружной поверхности клеточной мембраны вследствие двух причин: инактивации натриевых каналов и из-за работы Na+/К+-насоса. При достижении значений потенциала близких к значениям потенциала покоя часто наблюдается замедление реполяризации, которое связано с закрытием значительной части ворот K+-каналов, а также - с возрастанием противоположно направленного электрического градиента.
Однако процессы перемещения ионов через мембрану характеризуются достаточной инерционностью. Следовый гиперполяризационный потенциал возникает в результате выхода избытка К+ за пределы клетки. Это происходит по тему, что, несмотря на возникающее при реполяризации электрическое равновесие при достижении значений мембранного потенциала, соответствующего состоянию покоя, концентрационный градиент по калию все еще остается высоким. Состояние гиперполяризации всегда сопровождается повышением сопротивления мембраны и, следовательно, снижением возбудимости клетки. Длительная следовая гиперполяризация хорошо выражена в тонких немиелинизированных нервных волокнах, например, в болевых афферентах. Натрий-калиевый насос непосредственно на фазы потенциала действия не влияет, он работает как в состоянии покоя, так и продолжает работать во время развития потенциала действия.
Компенсирует эти процессы следовый деполяризационный потенциал (развивается не всегда). Причины возникновения этого потенциала просты: избыточный внешний калий действует на m-ворота Na+-каналов как подпороговый внешний стимул, внутренние h-ворота открываются по причине нарастания электроотрицательности. В результате этих процессов внутрь клетки просачивается некоторое количество ионов Na+. Такие изменения демонстрируют все свойства местного возбуждения, а именно: градуальны, затухают со временем. Как только К+ проницаемость вернется к исходному уровню регистрируется мембранный потенциал (покоя).
Возбудимость клеточной мембраны неоднократно изменяется в зависимости от фазы потенциала действия:
Кратковременное повышение возбудимости – на графике соответствует начальному участку кривой, до достижения значений 50-75% от КУД. Это состояние вызывается как подпороговыми, так и пороговыми (или надпороговыми) стимулами. В зависимости от силы раздражителя на этом участке графика формируется либо локальный ответ, либо участок кривой потенциала действия, соответствующий латентному периоду (времени, затраченному на преодоление порога раздражимости клетки). Возбудимость повышается потому, что клетка частично деполяризована, разность между величиной деполяризации и критическим уровнем деполяризации ниже пороговых значений. То есть мы получаем наглядное подтверждение снижения порога раздражимости. Любой дополнительный подпороговый стимул, воздействующий на данный участок мембраны в это время, может оказаться достаточным для развития полноценного потенциала действия.
Рефрактерность – самая длительная и самая важная фаза потенциала действия. Выделяют рефрактерность абсолютную и относительную. Графически абсолютный рефрактерный период начинается от точки пересечения восходящей кривой деполяризации до окончания овершута. Какой бы по силе дополнительный стимул не подействовал в данный момент – он не вызовет никаких изменений в развитии потенциала действия. Клетка не в состоянии распознать стимуляцию в этот период, потому что m-ворота Na+-каналов уже открыты, а еще закрытые открываются в ответ на уменьшение мембранного потенциала. По преодолении спайка, в начальном участке реполяризации рефрактерность тоже еще абсолютна так как, закрываются h-ворота Na+-каналов, и это состояние не изменяется даже при сильной стимуляции. Относительный рефрактерный период соответствует оставшемуся нисходящему участку реполяризации и включает в себя следовый гиперполяризационный потенциал. В это время происходит восстановление возбудимости, но концентрация внешнего калия все еще достаточно высока. В это время клетка способна ответить на сверхсильную стимуляцию, но амплитуда развивающегося при этом потенциала действия будет ниже исходной. Рефрактерность предохраняет клетку от перевозбуждения и отграничивает отдельные потенциалы действия во времени и пространстве. Чем короче время рефрактерного периода, тем выше лабильность нервного волокна. При продолжительности абсолютного рефрактерного периода 2мс нервное волокно способно генерировать потенциалы с частотой 500имп/с. Этот показатель типичен для большинства нервных волокон. Однако в перехватах Ранвье миелиновых двигательных волокон и в аксонах нейронов ретикулярной формации плотность натриевых каналов настолько высока, что лабильность достигает 1000имп/с.
Экзальтация – фаза кратковременного повышения возбудимости при следовом деполяризационном потенциале. Этому участку кривой соответствует повышенная проницаемость для катионов натрия, следовательно, порог раздражимости будет снижаться по сравнению с потенциалом покоя. Действующий в это время подпороговый стимул может привести к преодолению критического уровня деполяризации и развитию полноценного потенциала действия.
Заметим, что скорость смены возбудимости клеточной мембраны не является строго определенной величиной, она в значительной степени зависит от обменных процессов (метаболических и энергетических) и от силы и длительности стимуляции. Если в клетке снизится запас энергии (влияет на активный транспорт) или израсходуется запас метаболитов, в том числе потенциалобразующих ионов – лабильность и возбудимость будут стремительно падать. То же самое мы будем наблюдать при действии пессимального (длительного по времени или слишком сильного раздражителя): каждый раз активируясь в фазу относительной рефрактерности, клетка все слабее с более низкой амплитудой будет отвечать на стимуляцию, пока ответ станет невозможен вовсе. Такое состояние расценивается как парабиоз. Если стимул продолжает действовать и дальше – клетка погибает. Если стимуляция прекращается – через некоторое время (пессимальное торможение) клетка восстанавливает свои потенции и вновь готова отвечать на стимуляцию.