

Ядро клетки
Ядро является важнейшим компонентом клетки, содержащим ее генетический аппарат.
Функции ядра:
1. хранение генетической информации (в молекулах ДНК, находящихся в хромосомах);
2. реализация генетической информации, контролирующей осуществление разнообразных процессов в клетке – от синтетических до запрограммированной гибели (апоптоза);
3. воспроизведение и передачу генетической информации (при делении клетки).
Обычно в клетке имеется только одно ядро, однако встречаются многоядерные клетки, которые образуются вследствие деления клеток, не сопровождающегося цитотомией, или слияния нескольких одноядерных клеток (последние правильнее называть симпластами).
Форма ядра различных клеток неодинакова: встречаются клетки с округлым, овальным, бобовидным, палочковидным, многолопастным, сегментированным ядром; нередко на поверхности ядра имеются вдавления. Чаще всего форма ядра в целом соответствует форме клетки: оно обычно сферическое в клетках округлой или кубической формы, вытянутое или эллипсоидное в призматических клетках, уплощенное в плоских.
Расположение ядра варьирует в разных клетках; оно может лежать в центре клетки (в клетках округлой, плоской, кубической или вытянутой формы), у ее базального полюса (в клетках призматической формы) или на периферии (например, в жировых клетках).
Величина ядра относительно постоянна для каждого типа клеток, однако она может меняться в определенных пределах, увеличиваясь прй усилении функциональной активности клетки и уменьшаясь при ее угнетении.
Компоненты ядра. В ядре неделящейся (интерфазной) клетки выявляются кариолемма (ядерная оболочка), хроматин, ядрышко и кариоплазма (ядерный сок). Как будет видно из дальнейшего изложения, хроматин и ядрышко представляют собой не самостоятельные компоненты ядра, а являются морфологическим отражением хромосом, присутствующих в интерфазном ядре, но не выявляемых в качестве отдельных образований.
Ядерная оболочка
Ядерная оболочка (кариолемма) на светооптическом уровне практически не определяется; под электронным микроскопом обнаруживается, что она состоит из двух мембран - наружной и внутренней, -разделенных полостью шириной 15-40 нм (перинуклеарным пространством) и смыкающихся в области ядерных пор (рис. 3-18 и 3-19).
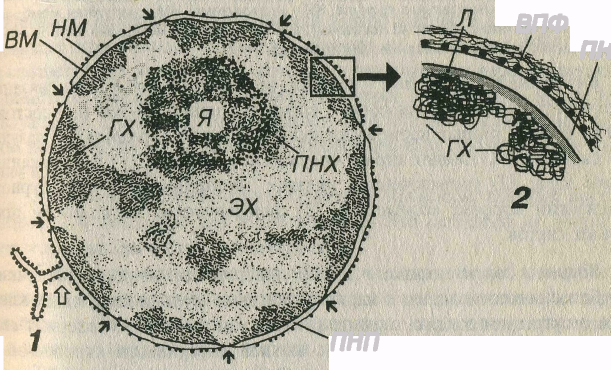
Рис. 3-18. Ядро клетки – общий вид (1) и участок ядерной оболочки (2). ГХ – гетерохроматин, ЭХ – эухроматин, Я – ядрышко, ПНХ – перинуклеолярный хроматин, НМ – наружная мембрана ядерной оболочки, ВМ - внутренняя мембрана, ПНП – перинуклеарное пространство, ВПФ – виментиновые промежуточные филаменты, Л - ламина. Черными стрелками показаны ядерные поры, белой – участок соединения ядерной оболочки с грЭПС.
Наружшя мембрана составляет единое целое с мембранами – на ее поверхности имеются рибосомы, а перинуклеарное пространство соответствует полости цистерн грЭПС и может содержать сиатезарованный материал. Со стороны цитоплазмы наружная мембрана окружена рыхлой сетью промежуточных (виментиновых) филаментов (см- рас. 3-18).
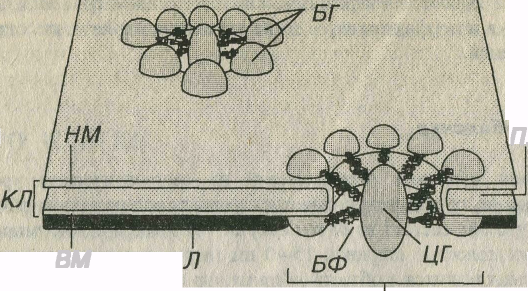
Рис. 3-19. Комплекс ядерной поры (КЯП). БГ – белковые гранулы, БФ – белковые фибриллы, ЦГ центральная гранула, КЛ – кариолемма: НМ – наружная мембрана (рибосомы на ее поверхности не показаны), ВМ – внутренняя мембрана кариолеммы, Л – ламина, ПНП – перинуклеарное пространство.
Внутренняя мембрана – гладкая, ее интегральные белки связаны с ядерной пластинкой – ламиной – слоем толщиной 80–300 нм, состоящим из переплетенных промежуточных филаментов (ламинов), образующих кариоскелет. Ламина играет очень важную роль в: (1) поддержании формы ядра; (2) упорядоченной укладке хроматина; (3) структурной организации поровых комплексов; (4) формировании кариолеммы при делении клеток.
Ядерные поры занимают 3-35% поверхности ядерной оболочки. Они более многочисленны в ядрах интенсивно функционирующих клеток и отсутствуют в ядрах спермиев. Поры (см. рис. 3-19) содержат два параллельных кольца (по одному с каждой поверхности кариолеммы) диаметром 80 нм, которые образованы 8 белковыми гранулами. От этих гранул к центру сходятся фибриллы, формирующие перегородку (диафрагму) толщиной около 5 нм, в середине которой лежит центральная гранула (по некоторым представлешям, это - транспортируемая через пору субъединица рибосомы). Совокупность структур, связанных с ядерной порой, называется комплексом ядерной поры. Последний образует водный канал диаметром 9 нм, по которому движутся мелкие водорастворимые молекулы и ионы. Гранулы поровых комплексов структурно связаны с белками ядерной ламины, которая участвует в их организации.
Ядерная оболочка в клетках животных и человека содержит до 2000-4000 поровых комплексов. В ядро из цитоплазмы через них поступают синтезированные белки, в обратном направлешш переносятся молекулы РНК и субъединицы рибосом.
Функции комплекса ядерной поры:
1. Обеспечение регуляции избирательного транспорта веществ между цитоплазмой и ядром.
2. Активный перенос в ядро белков, имеющих особую маркировку виде так называемой последовательности ядерной локализации – Nuclear Locflizftion Stquence (NLS), распознаваемой рецепторами NLS (в комплексе поры).
3. Перенос в цитоплазму субъединиц рибосом, которые, однако, слишком велики для свободного прохождения пор; их транспорт, вероятно, сопровождается изменением конформации порового комплекса.
Хроматин
Хроматин (от греч. chroma – краска) мелкие зернышки и глыбки материала, который обнаруживается в ядре клеток и окрашивается основными красителями. Хроматин состоит из комплекса ДНК и белка и соответствует хромосомам, которые в интерфазном ядре представлены длинными, тонкими перекрученными нитями и неразличимы как индивидуальные структуры. Выраженность спирализации каждой из хромосом неодинакова по их длине. Различают два вида хроматина – эухроматин и гетерохроматин.
Эухроматин соответствует сегментам хромосом, которые деспирализованы и открыты для транскрипции. Эти сегменты не окрашиваются и не видны в световой микроскоп.
Гетерохроматин соответствует конденсироватым, плотно скрученным сегментам хромосом (что делает их недоступными для транскрипцш). Он интенсивно окрашивается основньши красителями, и в световом микроскопе имеет вид гранул.
Таким образом, по морфологическим признакам ядра (соотношению содержания эу- и гетерохроматина) можно оценить активностъ Процессов транскрипции, а, следовательно, синтетической функции клетки. При ее повышении это соотношение изменяется в пользу эухроматина, при снижении – нарастает содержаше гетерохроматина. При полном подавлении функции ядра (например, в поврежденных и гибнущих клетках, при ороговении эпителиальных клеток эпидермиса – кератиноцитах при образовании ретикулоцитов крови) оно уменыпается в размерах, содержит только гетерохроматин и окрашивается основными красителями интенсивно и равномерно. Такое явление называется кариопикнозом (от греч. karion – ядро и pyknosis – уплотнение).
Распределение гетерохроматина (топография его частиц в ядре и соотношение содержания эу- и гетерохроматина характерны для клеток каждого типа, что позволяет осуществлять их идентификацию как визуально, так и с помощью автоматических анализаторов изображения. Вместе с тем, имеются определенные общие закономерности распределения гетерохроматина в ядре: его скопления располагаются под кариолеммой, прерываясь в области пор (что обусловлено его связью с ламиной) и вокруг ядрышка (перинуклеолярный гетерохроматин), более мелкие глыбки разбросаны по всему ядру (см. рис. 3-18).
Телъце Барра – скопление гетерохроматина, соответствующее одной Х-хромосоме у особей женского пола, которая в интерфазе плотно скручена и неактивна. В болылшстве клеток оно лежит у кариолеммы, а в гранулоцитах крови имеет вид маленькой добавочной дольки ядра ("барабанной палочки"). Выявление тельца Барра (обычно в эпителиальных клетках слизистой оболочки полости рта) используется как диагаостический тест для определения генетического пола (обязателен, в частности, для женщин, участвующих в Олимпийских Играх).
Упаковка хроматина в ядре. В деконденсированнюм состоянии длина одной молекулы (двойной спирали) ДНК, образующей каждую хромосому, равна в среднем, около 5 см, а общая длина молекул ДНК всех хромосом в ядре (диаметром около 10 мкм) составляет более 2 м (что сравнимо с укладкой нити длиной 20 км в теннисный мячик диаметром около 10 см), а в S-период интерфазы - более 4 м. Конкретные механизмы, препятствующие спутыванию этих нитей во время транскришщи и репликации, остаются нераскрытыми, однако очевидна необходимость компактной упаковки молекул ДНК. В клеточном ядре это осуществляется благодаря их связи со специальными основными (гистоновыми) белками. Компактная упаковка ДНК в ядре обеспечивает:
(1) упорядоченное расположение очень длинных молекул ДНК в небольшом объеме ядра;
(2) функциональный контроль активности генов (вследствие влияния характера упаковки на активность отдельных участков генома.
Уровни упаковки хроматина (рис. 3-20). Начальный уровень упаковки хроматина, обеспечивающий образование нуклеосомной нити диаметром 11 нм, обусловлен намоткой двойной нити ДНК (диаметром 2 нм) на блоки дисковидной формы из 8 гистоновых молекул (нуклеосомы). Нуклеосомы разделены короткими участками свободной ДНК. Второй уровень упаковки также обусловлен гистонами и приводит к скручиванию нуклеосомной нити с формированием хроматиновой фибриллы диаметром 30 нм. В интерфазе хромосомы образованы хроматиновыми фибриллами, причем каждая хроматида состоит из одной фибриллы. При дальнейшей упаковке хроматиновые фибриллы образуют петли (петельные домены) диаметром 300 нм, каждый из которых соответствует одному или нескольким генам, а те, в свою очередь, в результате еще более компактной укладки, формируют участки конденсированных хромосом, которые выделяются лишь при делении клеток.
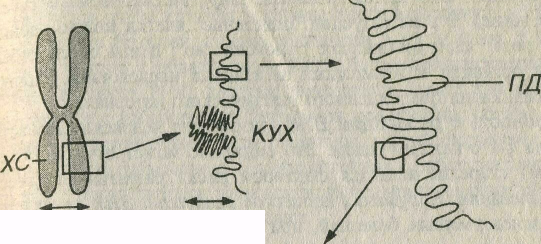
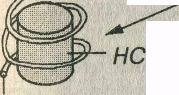
Рис. 3-20. Уровни упаковки хроматина в ядре клетки. Двойная спираль ДНК обраэует нить диаметром 2 нм, которая намотана на блоки дисковидной формы – нуклеосомы (НС), входящие в состав нуклеосомной нити (НН) диаметром 11 нм. Скрученная НН образует хроматиновую фибриллу (ХФ) диаметром 30 нм, которая формирует петельные домены (ПД) диаметром 300 нм. Более плотно упакованные ПД образуют конденсированные участки хромосомы (КУХ) диаметром 700 нм, являющиеся частью метафазной хромосомы (ХС) размером около 1400 нм.
В хроматине ДНК связана помимо гистонов также и с негистоиновыми белками, которые регулируют активность генов. Вместе с тем, и гистоны, ограничивая доступность ДНК для других ДНК-связывающих белков, могут участвовать в регуляции активности генов.
Функция хранения генетической информации в ядре в неизмененном виде имеет исключительно важное значение для нормальной жизнедеятельности клетки и всего организма. Подсчитано, что при репликации ДНК и в результате ее повреждений внешними факторами в каждой клетке человека ежегодно происходят изменения 6 нуклеотидов. Возникшие повреждения молекул ДНК могут исправляться в результате процесса репарации или путем замещения после распознавания и маркировки соответствующего участка.
В случае невозможности репарации ДНК при слишком значительных повреждениях включается механизм запрограммированной гибели клетки (см. ниже). В этой ситуации "поведение" клетки можно оценить как своего рода "альтруистическое самоубийство": ценой своей гибели она спасает организм от возможных негативных последствий реплификации и амплификации поврежденного генетического материала.
Способность к репарации ДНК у взрослого человека снижается примерно на 1% с каждым годом. Это снижение может отчасти объяснить, почему старение является фактором риска развития злокачественных заболеваний. Нарушения процессов репарации ДНК характерно для ряда наследственных болезней, при которых резко повышены как чувствительность к повреждающим факторам, так и частота развития злокачественных новообразований.
Функция реализации генетической информации в интерфазном ядре осуществляется непрерывно благодаря процессам транскрипции. Геном млекопитающих содержит около 3х109 нуклеотидов, однако не более 1% его объема кодирует важные белки и принимает участие в регуляции их синтеза. Функции основной некодирующей части генома неизвестны.
При транскрипции ДНК образуется очень крупная молекула РНК (первичный транскрипт), которая связывается с ядерными белками с образованием рибонуклеопротеинов (РНП). В первичном РНК-транскрипте (как и в матричной ДНК) имеются дискретные значашие последовательности нуклеотидов (экзоны), разделенные длинными некодирующими вставками (интронами). Процессинг РНК-транскрипта включает отщепление интронов и стыковку экзонов – сплайсинг (от анга. splicing – сращивание). При этом очень крупная молекула РНК превращается в достаточно мелкие молекулы иРНК, отделяющиеся от связашых с ними белков при переносе в цитоплазму.