

Г л а в а 3
ЦИТОЛОГИЯ:
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ КЛЕТКИ
ОБЩИЕ ПРИНЦИПЫ СТРУКТУРНО-ФУНКЦИОНАЛЬНОЙ
ОРГАНИЗАЦИИ КЛЕТКИ И ЕЕ КОМПОНЕНТЫ
Клетка – элементарная структурная, функциональная и генетическая единица в составе всех растительных и животных организмов. Организм взрослого человека состоит примерно из 1013 клеток, которые подразделяют более чем на 200 типов, существенно различающихся своими структурными и функциональными особенностями. Вместе с тем, клетки всех типов характеризуются сходством общей организации и строения важнейших компонентов.
Компоненты клетки. Каждая клетка состоит из двух основных компонентов – ядра и цитоплазмы. В ядре находятся хромосомы, содержащие генетическую информацию, которая в результате процесса транскрипции постоянно избирательно считывается и направляется в цитоплазму, где она контролирует ход многообразных процессов жизнедеятельности клетки, в частности, сбалансированные процессы синтеза, анаболизма (от греч. anabole – повышение), и разрушения, катаболизма (от греч. kataballo - разрушаю). Указанные процессы осуществляются в цитоплазме благодаря взаимодействию ее компонентов.
Компоненты цитоплазмы. Цитоплазма отделена от внешней (для данной клетки) среды внешней клеточной мембраной (плазмолеммой) и содержит органеллы и включения (рис. 3-1), погруженные в гиалоплазму (клеточный матрикс).
Органеллы - постоянно присутствующие в цитоплазме структуры, специализированные на выполнении определенных функций в клетке подразделяются на органеллы общего значения и специальные органеллы.
(1) органеллы общего значения имеются во всех клетках и необходимы для обеспечения их жизнедеятельности. К ним относятся митохондрии, рибосомы, эндоплазматическая сеть (ЭПС), комплекс Гольджи, лизосомы, пероксисомы, клеточный центр, компоненты цитоскелета;
(2) специальные органеллы имеются лишь в некоторых клетках и обеспечивают выполнение их специализированных функций. К ним относят реснички, жгутики, микроворсинки, миофибриллы, акросому (спермиев). Специальные органеллы образуются в ходе развития как производные органелл общего значения.
В состав многих органелл входит элементарная биологическая мембрана, поэтому органеллы подразделяют также на мембранные и немембранные. К мембранным органеллам относятся митохондрии, ЭПС, комплекс Гольджи, лизосомы, пероксисомы, к немембранным – рибосомы, клеточный центр, реснички, микроворсинки, жгутики, компоненты цитоскелета.
Функциональные системы (аппараты) клетки – комплексы органелл, которые под контролем ядра обеспечивают выполнение важнейших функций клетки. Выделяют: (1) синтетический аппарат; (2) энергетический аппарат; (3) аппарат внутриклеточного переваривания (эндосомально-лизосомальный); (4) цитоскелет.
Включения – временные компоненты цитоплазмы, образованные в результате накопления продуктов метаболизма клеток. Подразделяются на несколько типов (см. ниже).
Помимо структур цитоплазмы, которые можно четко отнести к органеллам или включениям, в ней имеется огромное количество разно-образных транспортных пузырьков, обеспечивающих не только перенос веществ между различными компонентами клетки, но и их частичное преобразование (процессинг) благодаря наличию ферментов в мембране, которая образует их стенку.
Мембранные структуры (компоненты) клетки – совокупное название различных структур цитоплазмы и ядра: плазмолеммы, ряда органелл, включений, транспортных пузырьков, а также ядерной оболочки (кариолеммы), в состав которых входят клеточные мембраны. Последние в различных мембранных структурах клетки организованы сходным образом, однако существенно различаются, в первую очередь, составом мембранных белков, определяющим специфику их функций.
Гиалоплазма (клеточный сок, цитозоль, клеточный матрикс) – внутренняя среда клетки, на которую приходится до 55% ее общего объема. Она представляет собой сложную прозрачную коллоидную систему, в которой взвешены органеллы и включения, и содержит различные биополимеры: белки, полисахариды, нуклеиновые кислоты, а также ионы. Претерпевает превращения по типу гель-золь. В гиалоплазме происходит большая часть реакций межуточного обмена.
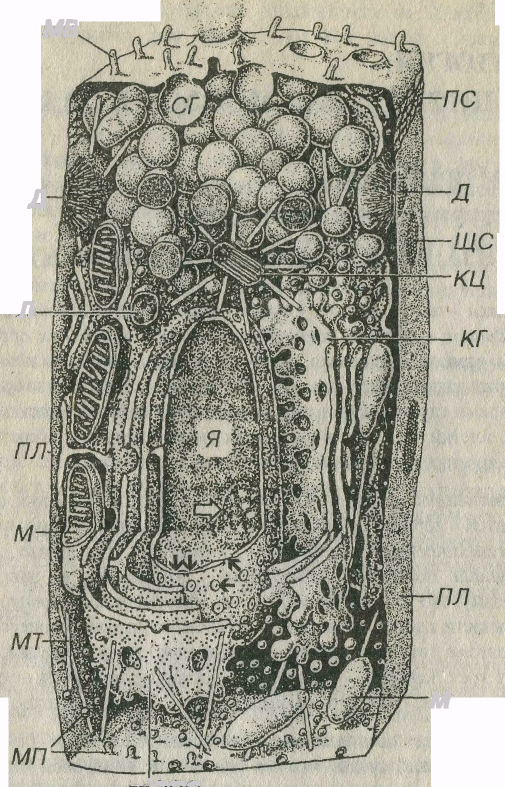
Рис. 3–1. Схема строения клетки (по R.V. Krstic, 1976). Я – ядро; ядрышко показано светлой стрелкой, кариолемма - двойными черными стрелками, ядерные поры – отдельными черными стрелками, М – митохондрии, КГ – комплекс Гольджи СГ – секреторные гранулы, Л – лизосома, КЦ - клеточный центр, МТ – микротрубочки ПЛ – плазмолемма, МП – микропиноцитозные пузырьки, МВ – микроворсинки, ПС – плотное соединение, ЩС – щелевое соединение, Д – десмосома.
ПЛАЗМОЛЕММА
Плазмолемма (внешняя клеточная мембрана, цитолемма, плазматическая мембрана) занимает в клетке пограничное положение и играет роль полупроницаемого селективного барьера, который, с одной стороны, отделяет цитоплазму от окружающей клетку среды, а с другой – обеспечивает ее связь с этой средой.
Функции плазмолеммы определяются ее положением и включают:
1. Распознавание данной клеткой других клеток и прикрепление к ним;
2. Распознавание клеткой межклеточного вещества и прикрепление к его элементам (волокнам, базальной мембране);
3. Транспорт веществ и частиц в цитоплазму и из нее (посредством ряда механизмов);
4. Взаимодействие с сигнальными молекулами (гормонами, медиаторами, цитокинами и др.) благодаря наличию на ее поверхности специфических рецепторов к ним;
5. Движение клетки (образование псевдо-, фило- и ламеллоподий) – благодаря связи плазмолеммы с сократимыми элементами цитоскелета. Структура плазмолеммы. Плазмолемма – самая толстая из клеточных мембран (7,5-11 нм); под электронным микроскопом она, как и другие клеточные мембраны, имеет вид трехслойной структуры, представленной двумя электронно-плотными слоями, которые разделены светлым слоем. Ее молекулярное строение описывается жидкостно-мозаичной моделью, согласно которой она состоит из липидного (фосфолипидного) бислоя, в который погружены и с которым связаны молекулы белков.
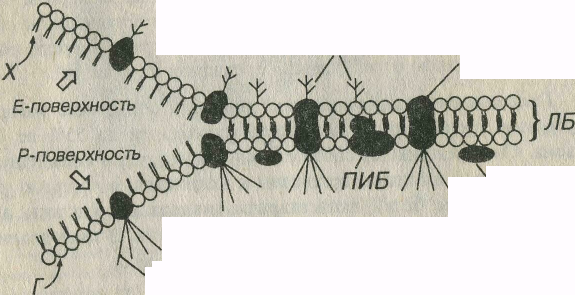
Рис. 3-2. Плазмолемма. ЛБ - липидный бислой: Г - головки (липидных моле- | кул), X - хвосты, ИБ - интегральные белки, ПИБ - полуинтегральные белки, ПБ - пе-'| риферические белки, МО - молекулы олигосахаридов, связанные с белками и липи- I дами, АМФ - актиновые микрофиламенты, связанные с белками плазмолеммы. Слева показаны поверхности мембраны, выявляемые в результате ее расщепления при за-;У мораживании-скалывании. Ш
Липидный бислой представлен преимущественно молекулами фосфатидилхолина (лецитина) и фосфатидилэтаноламина (цефалина), состоящими из гидрофильной (полярной) головки и гидрофобного (неполярного) хвоста. В состав большинства мембран входит также холестерин (холестерол). В мембране гидрофобные цепи обращены внутрь бислоя, а гидрофильные головки – кнаружи (рис.3-2). Состав липидов каждой из половин бислоя неидентичен. Липиды обеспечивают основные физико-химические свойства мембран, в частности, их текучесть при температуре тела. Некоторые липиды (гликолипиды) связаны с олигосахаридными цепями, которые выступают за пределы наружной поверхности плазмолеммы, придавая ей асимметричность. Электронно-плотные слои соответствуют расположению гидрофильных участков липидных молекул.
Мембранные белки составляют более 50% массы мембраны и удерживаются в липидном бислое за счет гидрофобных взаимодействий с молекулами липидов. Они обеспечивают специфические свойства мембраны (типы белков и их содержание в мембране отражают ее функцию) и играют различную биологическую роль (переносчиков, ферментов, рецепторов и структурных молекул). По своему расположению относительно липидного бислоя мембранные белки разделяются на две основные группы – интегральные и периферические (см. рис. 3-2).
Периферические белки непрочно связаны с поверхностью мембраны и обычно находятся вне липидного бислоя.
Интегральные белки либо полностью (собственно интегральные белки), либо частично (полуинтегральные белки) погружены в липидный бислой; часть белков целиком пронизывает всю мембрану (трансмембранные белки). Интегральные белки плазмолеммы хорошо выявляются при использовании метода замораживания-скалывания. При этом плоскость скола обычно проходит через гидрофобную середину бислоя, разделяя его на два листка – наружный и внутренний (см. рис. 3-2). Интегральные белки имеют вид округлых внутримембранных частиц, большая часть которых связана с Р-поверхностью (от англ. protoplsmic) – протоплазматической, т.е. ближайшей к цитоплазме поверхности скола (наружной поверхности внутреннего листка), меньшая – на Е-поверхности (от англ. external) - наружной, более близкой к внешней среде поверхности скола (внутренней поверхности наружного листка).
Часть белковых частиц связана с молекулами олигосахаридов (гликопротеины), которые выступают за пределы наружной поверхности плазмолеммы, другая имеет липидные боковые цепи (липопротеины). молекулы олигосахаридов связаны также с липидами с составе гликолипидов. Углеводные участки гликолипидов и гликопротеинов придают поверхности клетки отрицательный заряд и образуют основу так называемого гликокаликса (от греч. glycos – сладкий и calyx – оболочка), который выявляется под электронным микроскопом в виде рыхлого слоя умеренной электронной плотности, покрывающего наружную поверхность плазмолеммы. Эти углеводные участки играют роль рецепторов, обеспечивающих распознавание клеткой соседних клеток и межклеточного вещества, а также адгезивные взаимодействия с ними. В состав гликокаликса некоторые авторы включают, помимо углеводных компонентов, периферические мембранные белки и полуинтегральные белки, функциональные участки которых находятся в над мембранной зоне (например, иммуноглобулины). В гликокаликсе находятся рецепторы гистосовместимости, некоторые ферменты (часть которых может производиться не самой клеткой, а адсорбироваться на ее поверхности), рецепторы гормонов.
Белковые молекулы мозаично распределены в липидном бислое однако они не жестко фиксированы в нем, а напротив, могут перемещаться в его плоскости. В некоторых условиях определенные белки способны накапливаться в отдельных участках мембраны, образуя агрегаты. Перемещение белковых частиц, по-видимому, не является произвольным, а контролируется внутриклеточными механизмами, в которых участвуют микрофиламенты (см. цитоскелет), прикрепленные к некоторым интегральным белкам, связанным с Р-поверхностью (см. рис. 3-2).
Мембранный транспорт веществ может включать однонаправленный перенос молекулы какого-то вещества или совместный транспорт двух различных молекул в одном или противоположных направлениях.
Пассивный транспорт включает простую и облегченную диффузию – процессы, которые не требуют затраты энергии. Механизмом простой диффузии осуществляется перенос мелких молекул (например, О2, Н2О, СО2); этот процесс малоспецефичен и протекает со скоростью, пропорциональной градиенту концентрации транспортируемых молекул по обеим сторонам мембраны. Облегченная диффузия осуществляется через каналы и (или) белки-переносчики, которые обладают специфичностью в отношении транспортируемых молекул. В качестве ионных каналов выступают трансмембранные белки, образующие мелкие водные поры, через которые по электрохимическому градиенту транспортируются мелкие водорастворимые молекулы и ионы. Белки-переносчики также являются трансмембранными белками, которые претерпевают обратимые изменения конформации, обеспечивающие транспорт специфических молекул через плазмолемму. Они функционируют в механизмах как пассивного, так и активного транспорта.
Активный транспорт является энергоемким процессом, благодаря которому перенос молекул осуществляется с помощью белков-переносчиков против электрохимического градиента. Примером механизма, обеспечивающего противоположно направленный активный транспорт ионов, служит натриево-калиевый насос (представленный белком-переносчиком Na+-К+-АТФазой), благодаря которому ионы К+ выводятся из цитоплазмы, а ионы К+ одновременно переносятся в нее. Этот механизм обеспечивает поддержание постоянства объема клетки (путем регуляции осмотического давления), а также мембранного потенциала. Активный транспорт глюкозы в клетку осуществляется белком-переносчиком и сочетается с однонаправленным переносом иона Na+.
Облегченный транспорт ионов опосредуется особыми трансмембранными белками - ионными каналами, обеспечивающими избирательный перенос определенных Ионов. Эти каналы состоят из собственно транспортной системы и воротного механизма, который открывает канал на некоторое время в ответ на (а) изменение мембранного потенциала, (б) механическое воздействие (например, в волосковых клетках внутреннего уха), (в) связывание лиганда (сигнальной молекулы или иона).
Эндоцитоз. Транспорт макромолекул в клетку осуществляется с помощью механизма эндоцитоза (от греч. endo – внутрь и cytos – клетка). Материал, находящийся во внеклеточном пространстве, захватывается в области впячивания (инвагинации) плазмолеммы, края которого смыкаются с формированием эндоцитозного пузырька или эндосомы - мелкого сферического образования, герметически окруженного мембраной (рис. 3-3 и 3-5). Далее содержимое эндосомы подвергается внутриклеточной переработке (процессингу). В частности, в эндосоме в условиях закисления среды происходит отделение лиганда от рецептора (последний в дальнейшем используется повторно) - см. ниже. Разновидностями эндоцитоза служат пиноцитоз и фагоцитоз.
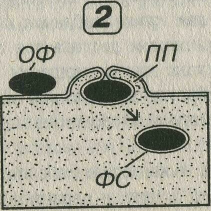
Рис. 3-3. Пиноцитоз (1) и фагоцитоз (2). ПС - пиносомы, ОФ - объект фагоцитоза, ПП - псевдоподии, ФС - фагосома.
Пиноцитоз (от греч. pinein – пить и cytos – клетка) - захват и поглощение клеткой жидкости и (или) растворимых веществ; подразделяется на макроптоцитоз (диаметр эндосом 0.2-0.3 мкм) и микроптоцитоз (диаметр эвдосом - 70-100 нм).
Фагоцитоз (от греч. phagein - поедать и cytos – клетка) - захват и поглощение клеткой плотных, обычно крупных (размером более 1 мкм) частиц; обычно сопровождается образованием выпячиваний цитоплазмы - псевдоподий, охватывающих объект фагоцитоза и смыкающихся над ним (см. рис. 3-3).
Рецепторно-опосредованный эндоцитоз. Эффективность эндоцитоза существенно увеличивается, если он опосредован мембранными рецепторами, которые связываются с молекулами поглощаемого вещества или молекулами, находящимися на поверхности фагоцитируемого объекта – лигандами (от лат. ligare - связывать). В дальнейшем (после поглощения вещества) комплекс рецептор–лиганд расщепляется, и рецепторы могут вновь возвратиться в плазмолемму.
Примером рецепторно-опосредованного взаимодействия может служить фагоцитоз лейкоцитом бактерии (см. рис. 7-8). Поскольку на плазмолемме лейкоцита имеются рецепторы к иммуноглобулинам (антителам), скорость фагоцитоза резко возрастает, если поверхность бактерии покрыта антителами (опсонинами - от греч. opson – приправа).
Окаймленные пузырьки и ямки. Рецепторы макромолекул в плазмолемме, перемещаясь латерально по клеточной поверхности, могут, связывая свои лиганды, накапливаться в области формирующихся эндоцитозных ямок. Очень часто вокруг таких ямок и образующихся из них пузьфьков со стороны цитоплазмы собирается сетевидная оболочка из белка клатрина, которая на срезах имеет вид щетинистой каемки (рис. 3-4). В покрытых клатриновой оболочкой (окаймленных) ямках; рецепторные белки мембраны вытесняют все остальные; таким образом; ямки действуют как приспособления для накопления и сортировки молекул. Этим механизмом достигается и значительная экономия в ходепроцесса эндоцитоза: для поглощения определенного количества молекул лиганда требуется значительно меньше пузырьков, чем было в случае диффузного распределения комплексов рецептор–лиганд.
Окаймленная ямка достигает своего максимального размера (около 0.3 мкм) в течение 1 мин. и превращается в окаймленный пузырек. Его содержимое может подвергаться процессингу лишь после того, как через несколько секунд он утратит клатриновую оболочку. Если она сохраняется, пузырек не способен сливаться с другими структурами (аналогичными пузырьками, лизосомами), и его содержимое остается неизмененным. Окаймленные эндоцитозные пузырьки транспортируют иммуноглобулины, белки желточных включений (в цитоплазму овоцитов), факторы роста, липопротеины низкой плотности (ЛНП). Некоторые транспортные мембранные пузырьки в цитоплазме окружены неклатриновой белковой оболочкой.
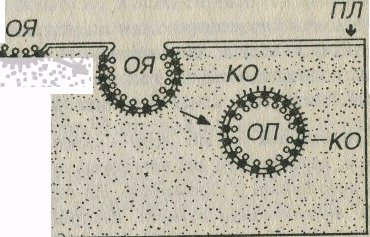
Рис 3-4 Рецепторно-опосредованный эндоцитоз. ПЛ - плазмолемма, Л - лиганд, Р - рецепторы, ОЯ - окаймленная ямка, ОП - окаймленный пузырек, КО - клатриновая оболочка.
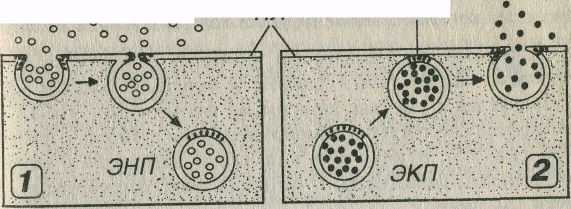
Рис 3-5 Эндоцитоз (1) н экзоцитоз (2). ВКП - внеклеточное пространство, ПЛ - плазмолемма, ЭНП - эндоцитозный пузырек, ЭКП - экзоцитозный пузырек, ФГБ - фузогенные белки.
Нарушение транспорта ЛНП описанным механизмом при врожденном наследственном заболевании - семейной гиперхолестеринемии - обусловлено отсутствием или наличием дефектных рецепторов ЛНП, неспособных связывать лиганд или накапливаться в окаймленных ямках. При этом поглощение клетками холестерина, поступающего с ЛНП, ослаблено, а его уровни в крови резко повышены, вызывая быстрое развитие атеросклероза и смерть больных в молодом возрасте от ищемической болезни сердца.
Экзоцитоз (от греч. exo – наружу и cytos – клетка) - процесс, обратный эндоцитозу, при котором мембранные экзоцитозные пузырьки приближаются к плазмолемме и сливаются с ней своей мембраной, которая встраивается в плазмолемму. При этом содержимое пузырька (продукты собственного синтеза клетки или транспортируемые ею молекулы, непереваренные и вредные вещества и др.) выделяется во внеклеточное пространство (см. рис. 3-5).
Судьба выделяемых экзоцитозом синтезированных клеткой молекул неодинакова: (1) прикрепляясь к клеточной поверхности, они могут становиться периферическими белками (например, антигенами); (2) они могут войти в состав межклеточного вещества (например, коллаген и гликозаминогликаны; (3) попадая во внеклеточную жидкость, могут выполнять роль сигнальных молекул (гормоны, цитокины).
Трансцитоз (от лат. trans – сквозь, через и греч. cytos – клетка) процесс, характерный для некоторых типов клеток, объединяющий признаки эндоцитоза и экзоцитоза. На одной поверхности клетки формируется эндоцитозный пузырек, который переносится к противоположной поверхности клетки и, становясь экзоцитозным пузырьком, выделяет свое содержимое во внеклеточное пространство. Процессы трансцитоза протекают очень активно в цитоплазме плоских клеток, выстилающих сосуды (эндотелиоцитах), особенно в капиллярах. В этих клетках пузырьки, сливаясь, могут образовывать временные трансцеллюлярные каналы, через которые транспортируются водорастворимые молекулы.
Ход образования эндоцитозных пузырьков опосредуется особыми (фузогенными - от лат. йшо - слияние) мембранными белками, которые концентрируются в участках инвагинации плазмолеммы. Эти же белки при экзоцитозе способствуют слиянию мембраны пузырька с плазмолеммой (см. рис. 3-5). Важную роль в процессах эндоцитоза и экзоцитоза играют элементы цитоскелета, в частности, микрофиламенты и трубочки (см. ниже).
Баланс процессов эндоцитоза и экзоцитоза. Эндоцитоз вследствие постоянной отшнуровки пузырьков с поверхности плазмолеммы должен приводить к уменьшению ее площади при одновременном увеличении объема клетки. Так, например, в макрофагах за 1 ч за счет эндоцитоза вносится до 25% объема цитоплазмы, а за 0.5 ч общая площадь поверхности эндоцитозных пузырьков составляет 100% площади плазмолеммы. При экзоцитозе, напротив, постоянно происходит увеличение площади плазмолеммы вследствие встраивания в нее мембраны экзоцитозных пузырьков. Так, в секреторной клетке ацинуса поджелудочной железы совокупная площадь мембраны секреторных гранул в 30 больше, чем поверхность плазмолеммы.
Вместе с тем, в действительности, активные процессы эндоцитоза и экзоцитоза не приводят к существенным изменениям площади поверхности плазмолеммы, так как они уравновешиваются формированием экзоцитозных и эндоцитозных пузырьков, соответственно, компенсирующим происходящую потерю мембраны или ее увеличение за счет противоположно направленного процесса. Эти явления отражают постоянно происходящий в клетке круговорот мембран, который получил название "мембранного конвейера".
Мембранные рецепторы являются преимущественно гликопротеинами, которые расположены на поверхности плазмолеммы клеток и обладают способностью высокоспецефически связываться со своими лигандами. Они выполняют ряд функций:
(1) регулируют проницаемость плазмолеммы, изменяя конформацию белков и ионных каналов;
(2) регулируют поступление некоторых молекул в клетку;
(3) действуют как датчики, превращая внеклеточные сигналы во внутриклеточные;
(4) связывают молекулы внеклеточного матрикса с цитоскелетом; эти рецепторы, называемые интегринами, играют важную роль в формировании контактов между клетками и клеткой и компонентами межклеточного вещества.
Рецепторы, связанные с каналами, взаимодействуют с сигнальной молекулой (нейромедиатора), которая временно открывает или закрывает воротный механизм, в результате чего инициируется или блокируется транспорт ионов через канал.
Каталитические рецепторы включают внеклеточную часть (собственно рецептор) и цитоплазматическую часть, которая функционирует как протеинкиназа (посредством таких рецепторов на клетки воздействуют инсулин и некоторые факторы роста).
Рецепторы, связанные с G-белками – трансмембранные белки, ассоциированные с ионным каналом или ферментом, – состоят из рецептора, взаимодействующего с сигнальной молекулой (первый посредник), и G-белка (гуанозин трифосфат-связывающего регляторного белка), включающего несколько компонентов, который передает сигнал на связанный с мембраной фермент (аденилат циклазу) или ионный канал, вследствие чего активируется второй внутриклеточный посредник – чаще всего циклический АМФ (цАМФ) или Са2+. Около 80% всех гормонов и нейромедиаторов действуют через рецепторы, связанные с эффекторными механизмами посредством G -белков.
В составе плазмолеммы находятся интегрины, называемые клеточными адгезионными молекулами (КАМ) – трансмембранные белки, служащие рецепторами для внеклеточных фибриллярных макромолекул фибронектина и ламинина (см. рис. 10-9). Фибронектин связывается с клетками и молекулами внеклеточного матрикса (коллагеном, гепарином, фибрином). Таким образом, фибронектин играет роль адгезионного мостика между клеткой и компонентами межклеточного вещества. Между тем, внутриклеточная часть молекулы интегрина через ряд других белков (талин, винкулин и α-актинин) связана с цитоскелетом.
Поверхностный аппарат клетки выделяется некоторыми авторами, которые рассматривают его как структурно и функционально единое образование, состоящее из трех компонентов: (1) надмембранного комплекса (гликокаликса), (2) плазмолеммы и (3) подмембранного комплекса (см. рис. 3-17). Первые два компонента описаны выше. Подмембранный комплекс образован специализированной периферической частью цитоплазмы, прилежащей к плазмолемме (кортикалъный слой) и содержащей элементы цитоскелета (см. ниже), преимущественно актиновые микрофиламенты. Более глубоко располагаются промежуточные филаменты и микротрубочки. Благодаря сокращению сети микрофиламентов, связанных с белками плазмолеммы, происходят изменения формы клетки и ее отдельных участков, формирование псевдоподий, выростов, перемещение клетки в пространстве.
СИНТЕТИЧЕСКИЙ АППАРАТ КЛЕТКИ
Синтетический аппарат клетки включает органеллы, участвующие в синтезе различных веществ, которые могут в дальнейшем использоваться самой клеткой или выделяться ею во внеклеточное пространство. Деятельность синтетического аппарата клетки, располагающегося в ее цитоплазме, контролируется ядром благодаря активности находящихся в нем генов. В синтетический аппарат входят рибосомы, эндоплазматическая сеть (ЭПС) и комплекс Гольджи.
Рибосомы
Рибосомы - мелкие (диаметр - 15-30 нм) плотные немембранные органеллы, обеспечивающие синтез белка путем соединения аминокислот в полипептидные цепочки. Информация о синтезе приносится к рибосомам информационнной РНК (иРНК), которая образуется в ядре в ходе считывания (транскрипции) фрагментов генетической информации с ДНК. Синтетически активная клетка содержит несколько миллионов рибосом (например, в клетке печени их число составляет 107, на которые приходится около 5% ее сухой массы.
Каждая рибосома состоит из двух асимметричных субъединиц: малой, связывающей РНК, и большой, катализирующей образование пептидных цепей (рис. 3-6). По форме малая субъединица напоминает телефонную трубку, большая - ковш. Субъединицы образованы рибосомальными РНК (рРНК), на которые приходится около 50% их массы, и особыми белками (до 80 различных видов). Первые образуются в ядрышке, белки же синтезируются в цитоплазме, после чего транспортируются в ядро, где связываются с рРНК. В дальнейшем субъединицы поотдельности через ядерные поры направляются из ядра в цитоплазму, где они участвуют в синтезе белка.
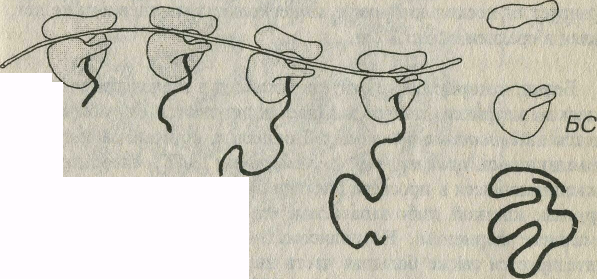
Рис. 3-6. Синтез белка на полирибосоме. Молекула синтезируемого полипептида (ПП) удлиняется по мере движения рибосом (Р), образующих полирибосому, по иРНК (направление показано стрелкой). По завершении синтеза ПП отделяется от Р, которые диссоциирует на две субъединицы - малую (МС) и большую (БС).
Рибосомы могут встречаться в цитоплазме поодиночке (в этом случае они функционально неактивны) или формировать скопления, которые называются полирибосомоми (полисомоми). В последних отдельные рибосомы (в количестве 3-30) удерживаются общей нитью иРНК толщиной 1.5 нм (см. рис. 3-6). Информация, переносимая иРНК, кодирует последовательность аминокислот в белке соответствующей последовательностью нуклеотидов. Рибосомы переводят (транслируют) эту генетическую информацию в реальную последовательность аминокислот в ходе белкового синтеза.
Функционально неактивные (нетранслирующие) рибосомы постоянно обмениваются своими субъединицами; их сборка происходит в начале синтеза белка, а по завершении синтеза одного полипептида они вновь обратимо диссоциируют.
Синтез белка рибосомой (см. рис. 3-6) начинается со связывания малой субъединицы с участком иРНК; далее рибосома передвигается вдоль цепи иРНК, причем на каждом этапе происходит специфическое присоединение к рибосоме молекулы транспортной РНК (тРНК), антикодон которой комплементарен соответствующему кодону иРНК В полипептид включается около 20 аминокислот в 1 секунду; белковая молекула среднего размера синтезируется за 20-60 с. Когда образование белковой цепочки завершается, субъединицы диссоциируют, освобождаясь от иРНК. Пока продолжается синтез белка данной рибосомой, новая рибосома занимает освобождающееся на иРНК место. По этой причине активно транслируемая иРНК находится в полисомах. Средняя продолжительность существования синтезированной белковой молекулы варьирует от нескольких минут до нескольких месяцев и даже лет, составляя в среднем около 2 сут.
Белки, которые после синтеза остаются в гиалоплазме (цитоплазматическом матриксе) клетки и далее используются ею, обычно синтезируются на свободных полисомах. Полисомы, которые своими большими субъединицами прикреплены к мембранам ЭПС, синтезируют белки, накапливающиеся в просвете цистерн ЭПС и в дальнейшем либо секретируемые клеткой, либо запасаемые ею внутри гранул (например, лизосомальные ферменты). На полисомах, связанных с мембранами ЭПС синтезируется также большая часть интегральных мембранных белков. Будет ли белок синтезироваться на ЭПС или на свободных полисомах зависит от характера начально образуемого отдела полипептидной цепи (сигнальной последовательности или пептида).
Присутствие значительного числа рибосом в цитоплазме клеток активно синтезирующих белок, придает ей при исследовании на свет оптическом уровне базофилию.