

1. 6.8 Действие физических и химических факторов окружающей среды на микроорганизмы
Каждый вид микробов имеет свои температурные зоны жизни: оптимальную, максимальную и минимальную. В оптимальной зоне микробная популяция размножается с заданной геномом скоростью. В зонах подавления рост клеток происходит со сниженной скоростью. В зависимости от температурных зон выделяют психрофильные, мезофильные и термофильные микроорганизмы.
Психрофильные микроорганизмы – группа микробов, нижняя граница роста которых находится около 0°С. Существуют факультативные и облигатные психрофильные микробы. Факультативные психрофильные микробы могут размножаться при +30°С. К ним относятся иерсинии, псевдомонады, клебсиеллы пневмонии. У облигатных психрофильных микробов верхняя граница роста соответствует 20°С, оптимальная зона 10-15°С. Они обитают в арктических и антарктических водах, ледяных пещерах, почвах вечной мерзлоты. Для психрофильных микробов характерны длительный период генерации, малая плотность популяции и медленно протекающие процессы биосинтеза.
Мезофильные микроорганизмы – группа микробов, температурные границы роста которых находятся в пределах 20-45°С (оптимальная температура 35-37°С). Обитают в организме теплокровных животных, в почве, воде, могут переживать в воздухе.
Термофильные микроорганизмы обитают при оптимальной температуре роста в диапазоне 45-93°С. Предельные температуры роста простейших находятся в границах 56°С, водорослей - 60°С, грибов - 60-62°С, фотобактерий - 70-72°С, хемолитотрофов – выше 90°С. Облигатные термофильные микробы обитают в фумаролах, кипящих и горячих источниках, промышленных и бытовых водах, самовозгорающихся материалах, конденсатах паровых труб.
Высушивание приводит к обезвоживанию микробной клетки и нарушению окислительно-восстановительных процессов. Наиболее чувствительны к обезвоживанию возбудители сифилиса, менингита, дизентерии, холеры и другие. Однако высушивание почти не действует на спорообразующие бактерии. Так, возбудители ботулизма и возбудители столбняка могут сохранять жизнеспособность в почве несколько лет.
Лиофилизация и высушивание под вакуумом при низкой температуре и последующее хранение в безвоздушной среде обеспечивает жизнеспособность на длительное время и используется для сохранения иммунобиологических препаратов и культур микроорганизмов.
Лучистая энергия. Ультрафиолетовые лучи, инфракрасные, гамма-лучи и солнечный свет воздействуют на микроорганизмы фотонами. Чем выше энергия фотонов, тем сильнее биологический эффект. Так, инфракрасные лучи, соприкасаясь с микроорганизмами, выделяют тепловую энергию, при этом лучистая энергия переходит в тепловую. Рентгеновские и гамма-лучи в больших дозах 280-440 Дж/кг вызывают ионизацию органических веществ, появление свободных радикалов, которые приводят к разрушению ядерного вещества и клеточной ДНК. Малые дозы стимулируют рост микробов. Микроорганизмы более устойчивы к радиоактивному излучению, чем высшие организмы. Их обнаруживают в воде атомных реакторов, залежах урановых руд (тионовые бактерии).
Ультрафиолетовые лучи распространяются в пространстве в виде электромагнитных волн длиной от 10 до 400 нм. Выделяют дальние (110-200 нм) УФ-лучи и ближние – 200-400 нм. В ближних УФ-лучах различают длинноволновую часть спектра (300-400 нм) и коротковолновую – 200-300 нм. Микробицидное действие характерно для коротковолновых УФ-лучей, особенно с длиной 250-270 нм. Механизм их действия связан с поглощением излучения молекулами ДНК, в результате чего образуются тиминовые, тимин-цитозиновые и цитозиновые димеры. Такие повреждения ДНК востанавливаются механизмами темновой репарации и фотореактивации. При действии высоких доз УФ-лучей происходит необратимая цепная реакция окисления липидов и белков. Высокочувствительны к УФ-излучению большинство вегетативных форм бактерий, наименьшей чувствительностью обладают возбудители туберкулеза, стафилококки.
2.Иммуноглобулины по структуре, антигенным и иммунобиологическим свойствам разделяются на пять классов: IgM, IgG, IgA, IgE, IgD (табл. 9.1). Иммуноглобулины M, G, А имеют подклассы. Например, IgG имеет четыре подкласса (IgG,, IgG2, IgG3, IgG4). Все классы и подклассы различаются по аминокислотной последовательности.
По их данным, молекулы иммуноглобулинов всех пяти классов состоят из полипептидных цепей: двух одинаковых тяжелых цепей Н (от англ. heavy — тяжелый) и двух одинако-
149
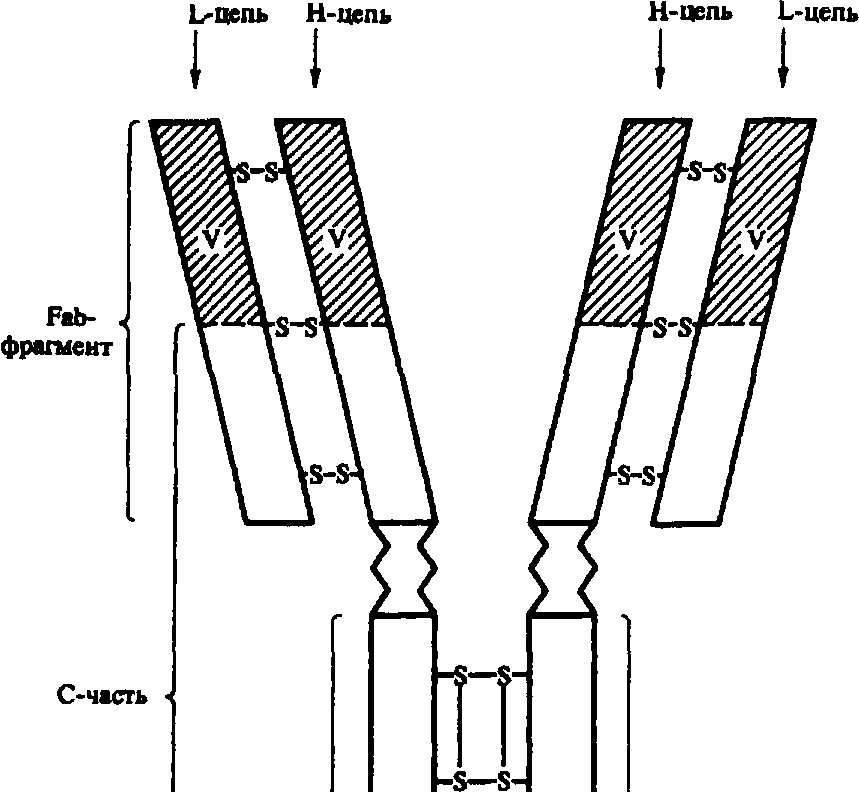
Fc-фрагмент
Рис.9.6. Строение иммуноглобулина класса G (схема). Объяснение в тексте.
вых легких цепей — L (от англ. light — легкий), соединенных между собой дисульфидными мостиками (рис. 9.6). Соответственно каждому классу иммуноглобулинов, т.е. М, G, А, Е, D, различают пять типов тяжелых цепей: ц (мю), Y (гамма), а (альфа), е (эпсилон) и 8 (дельта), имеющих молекулярную массу в пределах 50—70 кД (содержат 420—700 аминокислотных остатков) и различающихся по антигенности. Легкие цепи всех пяти классов являются общими и бывают двух типов: к (каппа) и X (ламбда); имеют молекулярную массу 23 кД (214—219 аминокислотных остатков). L-цепи иммуноглобулинов различных классов могут вступать в соединение (рекомбинироваться) как с гомологичными, так и с гетерологичными Н-цепями. Однако в одной и той же молекуле могут быть только идентичные L-цепи (к или х).
Как в Н-, так и в L-цепях имеется вариабельная — V (от англ. various — разный) область, в которой последовательность аминокислот непостоянна, и константная — С (от англ. constant — постоянный) область с постоянным набором аминокислот. В легких и тяжелых цепях различают NH2- и СООН-концевые группы.
Основной функцией иммуноглобулинов является их антительная активность. Эта функция осуществляется благодаря наличию у них Fab-фрагмента (антигенсвязывающей части молекулы).
Величина молекул иммуноглобулинов является одним из факторов, определяющих их способность проникновения в ткани. Большую часть иммуноглобулинов в сыворотке крови составляет IgG, на долю которого приходится до 80% от общего количества (пула) иммуноглобулинов. Присутствуют иммуноглобулины класса IgG и в тканях. Эти иммуноглобулины легко проникают через плаценту (табл. 4). IgG обеспечивают нейтрализацию вирусов и бактериальных токсинов, облегчают процесс фагоцитоза и лизис (разрушение) бактерий.
Иммуноглобулины класса IgM являются самыми крупными иммуноглобулинами и находятся преимущественно в кровяном русле, так как в меньшей степени способны проникать через стенки капилляров. IgM не могут проходить через плаценту. Благодаря наличию 10 антигенсвязывающих точек, IgM обладают высоким аффинитетом, т.е. способностью прочно связываться с антигенами. Особенно эффективно IgM участвуют в лизисе микроорганизмов, опосредованном комплементом.
Иммуноглобулины класса IgA по содержанию в сыворотке крови занимают второе место. В то же время иммуноглобулины этого класса являются доминирующими в секретах пищеварительного тракта и респираторной системы, а также в человеческом молозиве и грудном молоке. С IgA связан местный иммунитет слизистых оболочек к вирусам; кроме того, они препятствуют чрезмерному размножению микроорганизмов на поверхности слизистых оболочек. Иммуноглобулины класса А проявляют свою активность и в пищеварительном тракте, будучи более резистентными к действию протеолитических ферментов, чем антитела других классов.
Билет 13