

Молекулярная биология клетки Глава 2
.pdf11
ралкортикоидов. Холестерин участвует в формировании рафтов (плотиков) — дискретных мембранных доменов, богатых сфинголипидами и холестерином. Рафты представляют собой жидкостно-упоря- доченную фазу (участок плотноупакованных липидов) и обладают отличной от плазмолеммы плотностью и точкой плавления, так что они могут «плавать» — перемещаться в плоскости жидкостно-неупо-
Рис. 2-2. Молекулы плазматической мембраны. Наружный слой содержит преимущественно фосфатидилхолин, сфингомиелин и гликолипиды. Внутренний слой содержит фосфатидилэтаноламин, фосфатидилсерин и фосфатидилинозитол. Холестерин распределён по обоим слоям. Показаны отрицательные заряды головок фосфатидилсерина и фосфатидилинозитола. A — холестерин; B — олигосахарид в составе гликопротеина на наружной поверхности; C — погружённый интегральный белок; D — погружённый полуинтегральный белок; E — молекулы фосфолипидов; F — хвосты жирных кислот в составе фосфолипидов; G — полярные головки фосфолипидов; H — поверхностный (периферический) белок.
12
рядоченной плазмолеммы для выполнения определённых функций.
•Помимо липидов, в состав мембраны входят белки (в среднем до 60%). Они определяют большинство специфических функций мембраны;
◊периферические белки расположены на наружной или внутренней поверхности билипидного слоя;
◊полуинтегральные белки частично погружены в липидный билипидный слой на различную глубину;
◊трансмембранные, или интегральные белки пронизывают мембранунасквозь.
•Углеводный компонент мембран (до 10%) представлен олигосахаридными или полисахаридными цепями, ковалентно связанными с молекулами белков (гликопротеины) или липидов (гликолипиды). Цепи олигосахаридов выступают на наружной поверхности билипидного слоя и формируют поверхностную оболочку толщиной 50 нм — гликокаликс.
Функции плазматической мембраны
Основные функции плазмолеммы: трансмембранный транспорт веществ, эндоцитоз, экзоцитоз, межклеточные информационные взаимодействия.
Трансмембранный транспорт веществ. Транспорт веществ через плазматическую мембрану — это двустороннее движение веществ из цитоплазмы во внеклеточное пространство и обратно. Трансмембранный транспорт обеспечивает доставку в клетку питательных веществ, газообмен, выведение продуктов метаболизма. Перенос веществ через билипидный слой происходит путём диффузии (пассивная и облегчённая) и активного транспорта.
Эндоцитоз — поглощение (интернализация) клеткой воды, веществ, частиц и микроорганизмов. Эндоцитоз также происходит при перестройке или разрушении участков клеточной мембраны. К морфологически различаемым вариантам эндоцитоза относят пиноцитоз, фагоцитоз, опосредованный рецепторами эндоцитоз с образованием окаймлённых клатрином пузырьков и клатрин-независимый эндоцитоз с участием кавеол.
Экзоцитоз (секреция) — процесс, когда внутриклеточные секреторные везикулы (одномембранные пузырьки) сливаются с плазмолеммой, а их содержимое освобождается из клетки. При конститутивной (спонтанной) секреции слияние секреторных пузырьков происходит по мере их образования и накопления под плазмолеммой. Регулируемый экзо-
13 цитоз запускается с помощью определённого сигнала, чаще всего вслед-
ствие увеличения концентрации ионов кальция в цитозоле.
Межклеточныеинформационныевзаимодействия. Клетка,воспринимая различные сигналы, реагирует на изменения окружающей её среды изменением режимафункционирования. Плазматическая мембрана — место приложения физических (например, кванты света в фоторецепторах), химических (например, вкусовые и обонятельные молекулы, рН), механических (например, давлениеили растяжениев механорецепторах) раздражителей внешней среды и сигнальных молекул (лигандов) информационного характера из внутренней среды организма. Сигнальные молекулы (гормоны, цитокины, хемокины) специфически связываются с рецептором — высокомолекулярным веществом, встроенным в плазмолемму. Клетка-мишень при помощи рецептора способна узнавать лиганд и отвечать изменением режима функционирования при связывании этого лиганда с его рецептором. Рецепторы гормонов стероидной природы (например, глюкокортикоидов, тестостерона, эстрогенов), производных тирозина и ретиноевой кислоты локализуются в цитозоле.
ТРАНСПОРТВЕЩЕСТВ ЧЕРЕЗ ПЛАЗМАТИЧЕСКУЮ МЕМБРАНУ
Избирательная проницаемость мембраны поддерживает внутриклеточный гомеостаз, оптимальное содержание в клетке ионов, воды, ферментов и субстратов, обеспечивающих жизнедеятельность клетки. Гидрофобный характер сердцевины бислоя определяет возможность (или невозможность) непосредственного проникновения через мембрану различных с физико-химической точки зрения веществ (в первую очередь, полярных и неполярных). Неполярные вещества (например, холестерин и его производные) свободно проникают через биологические мембраны. Трансмембранный перенос полярных соединений осуществляют специальные транспортёры, встроенные в билипидный слой.
Существует три механизматрансмембранного переноса молекул: пассивный транспорт, облегчённая диффузия и активный транспорт. В зависимости от направления перемещения веществ и количества переносимых веществ данным транспортёром различают три типа транспорта: унипорт, симпорт и антипорт (рис 2-3).
Пассивный транспорт
Этот вид транспорта характеризуется низкой специфичностью. Молекулы в обоих направлениях перемещаются по градиенту концентрации без затрат энергии. Так, при дыхании диффузия газов происходит

14
Рис. 2-3. Типы транспорта. Унипорт — однонаправленный перенос ионов по концентрационному градиенту. Симпорт — сочетанный (совместный) транспорт двух и более веществ в одном направлении при помощи одного переносчика за счет разницы концентраций. Антипорт (противотранспорт) — согласованный перенос двух и более веществ через мембрану в противоположных направлениях. Перенос вещества в одном направлении сопряжён с потоком другого вещества в противоположном направлении.
по градиенту их концентрации, а определяющим диффузию фактором является парциальное давление O2 и CO2 (рис. 2-4).
Облегчённая диффузия
Облегчённая диффузия осуществляется с участием компонентов мембраны (ионные каналы, белки-переносчики, анионообменники) по градиенту концентрации и без непосредственных затрат энергии; проявляет специфичность по отношению ктранспортируемым молекулам (рис. 2-5).
Ионные каналы (унипорты) представлены большой группой гетеромультимерных интегральных мембранных белков, обеспечивающих избирательный транспорт ионов через фосфолипидный бислой мембраны из клетки в межклеточное пространство и обратно. Пору канала, как правило, образуют несколько белковых субъединиц. Просвет поры сформирован остатками гидрофильныхаминокислот и заполнен водой.

15
Рис. 2-4. Газообмен. Парциальное давление O2 (pО2) в тканях равняется 40 мм рт. ст., что способствует диссоциации оксигемоглобина. O2 свободно диффундирует из плазмы в ткани, а дезоксигемоглобин соединяется с Н+. В лёгких (pО2 = 100 мм рт. ст.) комплекс «дезоксигемог- лобин–Н+» диссоциирует, Н+ участвует в реакции Н+ + НСО3– → СО2 + Н2О, а дезоксигемоглобин связывается с O2, образуя оксигемоглобин.
Узкий просвет поры и поверхностный заряд образуют селективный фильтр, который определяет специфичность канала, т.е. его проницаемость для конкретного иона. В геноме человека закодирован синтез не менее 50 различных типов ионных каналов. Общее количество пептидов, связанных с работой каналов, достигает 400, 100 из которых клонированы и функционально охарактеризованы. Различают каналы утечки и воротные каналы.
•Каналы утечки позволяют ионам перемещаться по градиенту концентрации в клетку или из неё. Например, утечка из клетки положительно заряженных ионов калия приводит к тому, что внутренняя поверхность мембраны заряжается отрицательно относительно наружной.
•Воротные каналы постоянно закрыты и открываются в ответ на действие различных стимулов. Воротные каналы подразделяют на потенциалозависимые, внеклеточный лиганд-активируемые, внут-
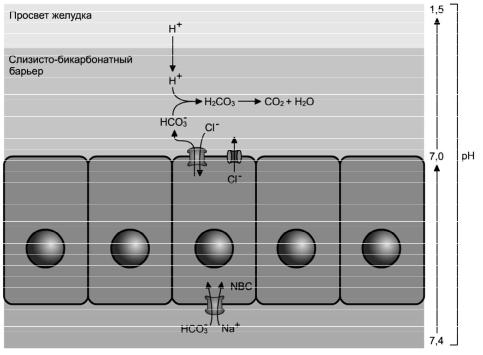
16
Рис. 2-5. Механизм нейтрализации соляной кислоты на слизистой оболочке желудка. Через базолатеральную мембрану с помощью симпорта Na+/HCO3- (NBC) HCO3- входит в клетку, транспортируется к апикальной поверхности и через анионообменник Cl-/HCO3- выводится из клетки в просвет желудка. Ионы хлора во внешнюю среду выходят через хлорный канал (унипорт). Ионы водорода, диффундирующие из просвета желудка в слизисто-бикарбонатный барьер, нейтрализуются ионами HCO3- с образованием Н2О и СО2.
риклеточный лиганд-управляемые и механочувствительные, или регулируемые объёмом клетки.
◊Натриевые каналы в возбудимых структурах (например, скелетные мышечные волокна, кардиомиоциты, нейроны) участвуют в генерации потенциала действия, присутствуют практически в любой клетке, не обязательно генерирующей потенциалы действия.
◊Калиевые каналы обнаружены в плазмолемме всех клеток. Их функции: поддержание мембранного потенциала, регуляция объёма клетки, модуляция электрической возбудимости нервных и
17
мышечных структур.
◊Кальциевые каналы (каналы депо кальция) участвуют в сокращении, секреции (в т.ч. гормонов и нейромедиаторов) и множестве иных клеточных процессов.
◊Хлорные каналы регулирует электрическую возбудимость плазмолеммы скелетных мышечных волокон, контролирует объём
клетки, участвует в образовании соляной кислоты в желудке. Водные каналы — мембранные поры для воды, образованные бел-
ком аквапорином. Семейство аквапорина включает не менее 12 белков, образующих мембранные каналы для воды в клетках различных тканей и органов. Вода — основное вещество, поступающее в клетки и выходящее из них. Организм человека в основном состоит из воды. Её содержание в общей массе тела постепенно уменьшается от 75% у новорождённых до 55% у пожилых людей. Внутриклеточная жидкость составляет 55%, внеклеточная жидкость — 45% всей воды организма. Вода служит растворителем, обеспечивает транспорт веществ и является средой для химических реакций. Движение воды в организме подчиняется законам осмоса.
•Аквапорин 1 (генAQP1, локализация 7p14).AQP1 экспрессируется в эритроцитах, в почке (проксимальные извитые канальцы и тонкий отдел петли Хенле), в тканях глаза обеспечивает гомеостаз внутриглазной жидкости.
•Аквапорин 2 (ген AQP2, 12q13) экспрессируется только в собирательных трубочках почки. Активность этого канала регулирует антидиуретический гормон (АДГ), увеличивая реабсорбцию воды из просвета трубочек в межклеточное пространство. Нефрогенный несахарный диабет (тип II, 107777, 12q13) — следствие мутации гена AQP2.
•Аквапорин 3 (ген AQP3, локализация 7q36.2-q36.3) — водный канал базолатеральных мембран собирательных трубочек почки. Экспрессия AQP3 найдена также в печени, поджелудочной железе, кишечнике, селезёнке, простате.
•Аквапорин 4 (ген AQP4, локализация 18q11.2-q12.1) экспрессируется в клетках эпендимной выстилки сосудистого сплетения желудочков и водопровода мозга, в синтезирующих вазопрессин нейросекреторных нейронах гипоталамуса. Этот канал расценивают как осморецептор. AQP4 присутствует в астроцитах. При тяжёлой форме множественного склероза, известной как оптиконевромиелит (болезнь Девика), в организме больных вырабатываются аутоантитела, которые атакуют этот водный канал.
•Аквапорин 5 (ген AQP5, локализация 12q13) принимает участие в
18
формировании слёзной жидкости, слюны, секретов желёз воздухоносных путей.AQP5 выявлен в эпителиальных клетках околоушной, подчелюстной, подъязычной, слёзной, потовой железы, трахеи, легких.
Осмос — поток воды через полупроницаемую мембрану из компартмента с меньшей концентрацией растворённых в воде веществ в компартмент с большей их концентрацией. Поток воды через биологические мембраны определяет разность осмотического и гидростатического давлений по обе стороны мембраны. Численно осмотическое давление при равновесном состоянии (вода перестала проникать через полупроницаемую мембрану) равно гидростатическому давлению. По отношению к клетке растворы, в которых она находится, могут быть изо-, гипер- и гипоосмотическими (иногда применяют термин «–тонический», справедливый для электролитов).
•Изоосмотические (изотонические) растворы — концентрация растворённых веществ и осмотическое давление такие же, как в клетке. В этих растворах объём клетки остаётся неизменным. Изотонические растворы называются физиологическими (0,85% раствор NaCl). Вливание изотоничного солевого раствора увеличивает объём межклеточной жидкости, но не влияет на объём внутриклеточной жидкости.
•Гиперосмотические (гипертонические) растворы — концентрация растворённых веществ и осмотическое давление выше, чем в клетке. Клетки в таких растворах уменьшаются в объёме («сжимаются»), поскольку теряют воду.
•Гипоосмотические (гипотонические) растворы — концентрация веществ и осмотическое давление меньше, чем в клетке. Клетки в таких растворахв результате поглощения воды увеличиваются в объёме («набухают»). Такое состояние клетки называют тургором, а гидростатическое давление,оказываемое на мембрану—тургорным. При избыточном поступлении воды в клетку нарушается целостность плазматической мембраны, а содержимое клетки выходит во внеклеточное пространство.
Трансмембранные белки-переносчики (симпорты) участвуют в сочетанном транспорте двух веществ, при котором движущей силой является перемещение по градиенту концентрации одного из веществ, например, ионов натрия. При помощи Na+ в клеткутранспортируются глюкоза (симпорт Na+/глюкоза), аминокислоты (симпорт Na+/АК), фосфаты (симпорт Na+/Рi), бикарбонат (симпорт Na+/HCO3-), хлор (симпорт для
Анионообменники(антипорты) — регуляторы внутриклеточного pH.

19
Среди переносчиков этой группы хорошо изучены анионообменники Cl– на бикарбонат и Na+ на H+ .
Активный транспорт
Активный транспорт происходит при участии АТФаз (насосов), обеспечивающих энергозависимый трансмембранный перенос ионов против электрохимического градиента. Наиболее известны следующие насосы: натрий, калиевый (Na+,K+-АТФаза) антипорт, протонный (H+,K+- АТФаза) антипорт и кальциевый (Ca2+-АТФаза) унипорт.
•Натрий, калиевая АТФаза выкачивает Na+ из клетки в обмен на K+, поддерживает мембранный потенциал покоя и участвует в генерации потенциала действия в мембране нервных и мышечных клеток (рис. 2-6).
•Протонная, калиевая АТФаза участвуют в образовании соляной кислоты (электронейтральный обмен внеклеточного K+ на внутриклеточный H+) париетальными клетками желёз слизистой оболочки желудка (рис. 2-7).
•Кальциевая АТФаза откачивает ионы кальция из цитозоля против концентрационного градиента во внутриклеточные депо кальция (цистерны гладкой эндоплазматической сети).
Рис. 2-6. Генерация мембранного потенциала покоя. Натрий, калиевая АТФаза поддерживает трансмембранный градиент Na+ и K+. При гидролизе одной молекулы АТФ три иона натрия выкачивается из клетки и два иона калия закачивается в неё. Калиевые каналы пассивной утечки контролируют мембранный потенциал покоя и возбудимость мембраны нервных и мышечных клеток [по Dudek R.W., 2004].

20
Рис. 2-7. Образование HCl в желудке. Карбоангидраза катализирует реакцию CO2 + H2O → H+ + HCO3–. Н+,К+-АТФаза обеспечивает выход H+ через апикальную мембрану по всей поверхности внутриклеточных канальцев в обмен на К+. Cl– транспортируется в клетку в обмен на HCO3– через мембрану боковой поверхности, а выходит через апикальную мембрану через Cl-–канал. Na+,К+–АТФаза, К+–каналы и анионообменник NHE поддерживают внутриклеточный гомеостаз ионов при активации клетки.