

ЗАТ ЖӘНЕ ЭНЕРГИЯ АЛМАСУЫ.
Заттар алмасуы – бұл ағзаның тіршілігі үшін қажетті заттар мен энергия алмасуын қамтамассыз ететін ағзадағы химиялық реакциялардың жиыны. Заттар алмасуын келесі 3 этапқа бөледі: 1-ағзаға заттардың түсуі; 2-аралық алмасу немесе метаболизм; 3-соңғы өнімдердің шығарылуы.
Ағзаға заттардың түсуі тыныс алу (оттегі), қоректену және асқорыту нәтижелерінде өтеді. Тағам құрамында түскен белоктар, майлар, көмірсулар асқорыту трактісі ферменттерінің әсерінен қанға оңай сіңірілетін қарапайым құрам бөліктеріне (аминоқышқылдар, май қышқылдары және моносахаридтер) ыдырайды.
Тағам құрамында түсетін қосылыстар метаболизмнің субстраттары деп аталады.
Метаболизм (зат алмасуы) – бұл химиялық реакциялардың жиыны болып табылады. Метаболизм қарама қарсы бағыттағы екі фазадан катаболизм және анаболизмнен тұрады.
Катаболизм – бұл күрделі заттардың қарапайым құрам бөліктеріне ыдарауы процессі. Катаболизм процессінде күрделі органикалық молекулалар соңғы өнімдер СО2, Н2О және мочевинаға дейін ыдырайды. Катаболизм реакциялары энергия бөлініп жүретін болғандықтан экзергоникалық болады.
Анаболизм қарапайым заттардан күрделі қосылыстардың синтезделуі процессі. Аанболизм реакциялары энергия қабылдау арқылы өтетіндіктен эндергоникалық болады.
Амфиболизм – бұл катаболизм мен анаболизмнің аралығындағы фаза. Ол бір заттың ыдырауында басқа заттың синтезделуі қосарланып жүретін процес.
Метаболиттік цикл – бұл осы процеске қосылған қосылыстың соңғы өнімнің реакцияның бастапқы өнімі болып түзілуі.
Метаболиттік жол - бұл ағзадағы белгілі бір заттың химиялық реакцияларының кезектескен реті мен сипаттамасы. Метаболизм процессінде түзілген аралық өнімдер метаболиттер деп аталады, ал метаболиттік жолдағы соңғы қосылыс – соңғы өнім деп аталады.
Метаболизмнің интенсивтілігі торшаның қандай да бір зат немесе энергияға қажеттілігімен айқындалады және келесі төрт жолмен реттеледі:
1) Белгілі бір метаболиттік жолдың реакцияларының қосынды жылдамдығы бұл бұл жолдағы әрбір ферменттің концентрациясына, ортаның рН-мәніне, бұл жолдағы аралық өнімдердің торшаішілік концентрациясына, кофактор және кофермент концентрацияларына тәуелді;
2) Метаболиттік жолдың бастапқы және соңғы этаптарын катализдейтін реттеуші (аллостерлік) ферменттердің активтілігін байланысты;
3) Қандай да бір реакцияға қатысушы ферменттің синтезделу жылдамдығын айқындаушы генетикалық бақылау бойынша;
4) Гормондық реттелу. Кейбір гормондар метаболиттік жолдың ферменттерін активтендіруге немесе ингибирлеуге қабілетті.
Энергия алмасуында орталық рольді АТФ АДФ + Фк циклі атқарады. АТФ молекуласында үш макроэргиялық байланыс болады. АТФ-тің АДФ пен Н3РО4-ке гидролиттік ыдырауында макроэргиялық байланыстан 7,3 ккал /моль энергия бөлінеді. Ағзада энергия АТФ күйінде аккумуляцияланады және оның АДФ пен фосфор қышқылынан түзілуінде соншалықты энергия (7,3 ккал /моль) қабылданады. АТФ молекуласының макроэргиялық байланыстарына қажетті энергия катаболизм процессінде бөлінеді. Ағзада АТФ энергиясы келесі 5 жұмысқа жұмсалады: 1.механикалық жұмыс; 2.тасымалдану жұмысы; 3.химиялық жұмыс (синтез); 4.электрлік жұмыс; 5.жылу түзілуі.
Қалыпты жағдайда, адам тәулігіне орта есеппен 27 моль оттегіні қабылдайды. Оның негізгі бөлігі (25 моль) тыныс алу тізбегінде АТФ синтезі үшін жұмсалады. Сондықтан тәулік сайын 125 моль немесе 62 кг АТФ синтезделеді. Бірақ, бүкіл ағзадағы АТФ массасы небары 20-30 г. Бұл нәтиже АТФ молекуласының бір тәулікте 2500 рет синтезделіп және сонша рет энергия беру үшін гидролизденетінін көрсетеді. Бұл АТФ АДФ + Фк циклінің интенсивтілігін көрсетеді.
АТФ энергиясы жұмсалатын негізгі процестер:
1.Механикалық жұмыс, бұл бұлшықеттердің жиырылуы мен босаңсуының реакциялары.
2.Тасымалдану жұмысы-негізінен заттардың торша мембранасы арқылы активті тасымалдануы АТФ 30%-ті Na/K –АТФ-аза қызметі үшін жұмсалады.
3.Химиялық жұмыс – бұл ағзадағы синтез және молекулаішілік изомерлену реакциялары.
4.Электрлік жұмыс – бұл жүйке импульстерін беру жұмыстары.
5.Жылу түзілу жұмысы – бұл ағза ферменттерінің оптимальды қызметі үшін қажетті температура мәнін тұрақты ұстау.
Катаболизмнің жалпы және арнайы жолдары.
Катаболизм жалпы үш этапқа бөлінеді:
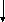
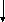
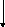 Белоктар
Көмірсулар
Майлар
Белоктар
Көмірсулар
Майлар
І-этап
 аминоқышқылдар
глюкоза майқышқылдар
аминоқышқылдар
глюкоза майқышқылдар
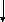

 глицерин
глицерин

пируват
 NН3
NН3
ІІ этап
ацетил-КоА
І ІІ
этап
ІІ
этап

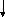
Со2 Н2О АТФ
Сурет 1. Таѓамныњ негізгі заттарыныњ катаболизмі.
Катаболизмніњ бастапќы этапы немесе спецификалыќ жолы єрбір кластаѓы ќосылыстардыњ спецификалыќ ферменттерініњ ќатысуымен µтеді. Нєтижесінде кµп мµлшердегі бастапќы заттарды аралыќ µнімдер арќылы пируват жєне ацетил-КоА т‰зіледі. Ацетил-КоА пируваттыњ тотыѓып декарбоксилденуі жєне дегидрленуінен т‰зіледі. Екінші жєне ‰шінші этаптыњ реакциялары катаболизмніњ жалпы жолдары деп аталады. Ол таѓамдыќ заттардыњ ыдырап пируваттан соњѓы µнімдер СО2, Н2О жєне АТФ т‰зілуінен т±рады.
Екінші этап реакциясы - б±л пируваттыњ тотыѓып декарбоксилденуі. ‡шінші этап реакциялары-б±л цитрат циклі.
Катаболизмніњ жалпы жолдарында ѓана дегидрлену реакцияларына арналѓан субстраттардыњ негізгі массасы т‰зіледі. Ары ќарай тыныс алу тізбегі мен фосфорлану реакциялары катаболизм жолына ќосарланып энергия АТФ формасына жиналады. Пируваттыњ тотыѓу реакциялары (ПТР) митохондрия матриксінде µтеді.
Пирож‰зім ќышќылыныњ (пируват) тотыѓып декарбоксилденуі.
Кµп стадиялы к‰рделі реакциялардыњ тењдеуі келесі т‰рде беріледі.
СООН КоА
С = О + NAD+ + HSKoA С = О + NADН + СО2
СН3 СН3
Пируват ацетил-КоА
Сурет 2.
Реакция кезектескен ретпен ж±мыс істейтін бір комплекске жинаќталѓан пируват дегидрогеназдыќ комплекс арќылы атќарылады. Ол ‰ш ферменттен т±рады.
Сурет 3.
Пируватдегидрогеназдыќ комплекс жєне оныњ ж±мысы. Б±л комплекс митохондрияныњ ішкі мембрансында орналасќан жєне оѓан матрикс жаѓынан ќосылады.
Ферменттердіњ б±л комплексі конвейер тєрізді ж±мыс істеп µнімдерді эстафета бойынша береді.
1-ші фермент - пируватдекарбоксилаза. Оныњ коферменті ретінде В1 витаминініњ туындысы - тиаминдифосфат болады. Фермент карбоксилдік топты СО2 т‰рінде бµледі де ацетилдік ќалдыќты екінші ферменттіњ коферменті липой ќышќылына ќосады.
Нєтижесінде ацетил-липоат ќосылысы т‰зіледі.
2-ші фермент - дегидролипоатацетилтрансфераза липой ќышќылындаѓы ацетилдік ќалдыќтыњ екінші коферменті НS-КоА-ѓа ќосылуын катализдейді. Нєтижесінде ацетил-КоА т‰зіледі. Б±л реакциядаѓы липоат ферментке мыќты байланысќан кофермент, ал НS-КоА реакция кезінде ќосылады. ФАДН2-ден сутегілер липой ќышќылына байланысып оны дегидролипоатќа айналдырады.
3-ші фермент-дегидролипоатдегидрогеназа липой ќышќылынан сутегілерді бµліп, оны ќайтадан ќалпына келтіреді (липоатта к‰кіртті кµпірше байланысы -S-S-ќайта т‰зіледі). Ал, сутекті ќабылдаѓанда т‰зілген НАДН-ы ары ќарай тыныс алу тізбегіне ќосылады.
Сурет 4. НS-КоА-ныњ ќ±рылысы.
Ары ќарай, негізгі т‰зілген µнім ацетил-КоА цитрат циклінде тотыѓады.
Ацетил-КоА май ќышќылдарыныњ, холестеринніњ, кетондыќ денелердіњ синтезініњ реакцияларына ќатысады. Оныњ ењ ‰лкен бµлігі лимон ќышќылын синтездеп ‰шкарбон ќышќылыныњ цикліне ќосылады.
Цитрат циклі.
Цитрат циклі (Кребс циклі ‰шкарбон ќышќылы циклі, лимон ќышќылыныњ циклі) - б±л бос кµміртекті ацетилдік фрагменттіњ толыќ тотыѓуына єкелетін реакциялар ж‰йесі (Сур. 5 жєне 6). Циклдіњ Ханс Кребс кµптеген реакцияларын ашып зерттегендіктен оныњ атын берген жєне 1953 жылы Новель сыйлыѓы берілді.
Цитрат циклі белоктар, кµмірсулар, майлар тотыѓуыныњ жалпы жєне соњѓы жолы болып табылады. Цитрат циклініњ барлыќ реакциялары митохондрияда шоѓырланѓан.
Толыќ цикл жолында келесі процесстер µтеді:
ацетилдік ќалдыќтыњ толыѓымен СО2-ніњ екі молекуласына дейін тотыѓуы;
тотыќсызданѓан НАД+ -тіњ 3 жєне ФАД+ -тіњ 1 молекуласы т‰зіледі;
субстраттыќ фосфорлануда ГТФ-тіњ 1 молекуласы т‰зіледі.
Сурет 5.
Сурет 6.
Сурет 6. Цитрат циклініњ ферменттері.
1-пируватдегидрогеназдыќ комплекс, 2-цитратсинтаза, 3-аконитаза, 4-изоцитратдегидрогеназа, 5--кетоглутаратдегидрогеназалыќ комплекс, 6-сукцинил-КоА-синтетаза, 7-сукцинатдегидрогеназа, 8-фумараза, 9-малатдегидрогеназа.
Цитратсинтаза. Циклдіњ бірінші реакциясы-б±л оксалоацетат пен ацетил-КоА-ныњ конденсациялануы. Реакция АТФ-тіњ гидролизденуінен бµлінген энергияны пайдаланып, цитратсинтазаныњ катализдеуімен цитрат т‰зіп µтеді.
Аконитаза. Циклдіњ екінші реакциясы - б±л цитраттыњ аконитаза ферменті єсерінен молекулаішілік ќайта ќ±рылып (изомерленіп) изоцитраттыњ т‰зілуі. Ќ±рамында гемдік емес темірі болатын, стереоспецификалыќ фермент аконитаза -ОН тобын кµрші кµміртегіне ауыстырады.
(ИДГ)
Изоцитратдегидрогеназа. Циклдіњ ‰шінші реакциясы б±л изоцитраттыњ тотыѓып декарбоксильденіп жєне дегидрленіп, изоцитратдегидрогеназа катализдеуімен -кетоглутараттыњ т‰зілуі. Реакцияда ИДГ кофакторы НАД+ екі протонды ќосып алып НАДН2-ге айналады. ИДГ митохондриялыќ -кофакторы НАД+ жєне цитоплазмалыќ пен митохондриялыќ -кофакторы НАДФ+ т‰рлеріне бµлінеді. Б±л реакциядаѓы т‰зілген СО2 цитрат синтезіне ж±мсалады.
-Кетоглутаратдегидрогеназа. Циклдіњ тµртінші реакциясы-б±л -кетоглутараттыњ декарбоксилденіп жєне дегидрленіп, -кетоглутаратдегидрогеназа катализдеуімен, сукцинил-КоА т‰зіледі. Реакция барысында СО2 жєне тотыќсызданѓан НАДН2 бµлінеді. Б±л ќайтымсыз реакция Сукцинил-КоА-да макроэргиялыќ байланысы бар ќосылыс.
Сукцинил-КоА-синтетаза (сукцинилтиокиназа). Циклдік бесінші реакциясы-б±л. Сукцинил-КоА-ныњ сукцинатќа айналады. Макроэргиялыќ байланыстыњ ‰зілуінен т‰зілген энергиядан ГДФ фосфорланып ГТФ т‰зіледі.
Бұл субстраттық фосфорлану реакциясы да болып табылады.
Сукцинатдегидрогеназа. Циклдің алтыншы реакциясы бұл сукцинаттың фумаратқа дегидрленіп айналуы.
Реакцияда ФАД екі протонды қабылдап ФАДН2-ге айналады.
Фумараза. Циклдің жетінші реакциясы – бұл фумаразаның катализдерімен малаттың түзілуі.
Малатдегидрогеназа. Циклдің сегізінші реакциясы бұл малаттың дегидрленіп оксалоацетатқа айналуы. Реакцияда үшінші тотықсызданған. НАДН2 түзіледі.
Осы реакциямен циклдің бір айналымы аяқталады. Түзілген оксалоацетат жаңа келген ацетил-КоА молекуласымен циклдің жаңа айналымын бастауға қатыса алады.
Циклдің қосынды теңдеуі.
СН3
│
 С
═ О + 2 О2 2 СО2 + Н2О
+ НS-КоА
С
═ О + 2 О2 2 СО2 + Н2О
+ НS-КоА
S КоА
12АДФ+Ф 12АТФ+Н2О
Цитрат циклінің биологиялық маңызы.
Цитрат циклінің (ЦЦ) негізгі маңызы бұл: 1.ЦЦ –АТФ-тің негізгі шығу көзі; 2.ЦЦ-бұл барлық кластағы қосылыстардың катаболизмінің терминалдық этапы, 3.ЦЦ-анаболизм процессінде маңызды роль атқарады. Мысалы, цитраттан май қышқылдары, -кетоглутараттан аминоқышқылдар, сукцинил-КоА-дан гемм синтезі және т.б.
Катаболизмнің жалпы жолдарының тыны салу тізбегімен қосарлануы.
|
|
Катаболизмнің жалпы жолдарында 5 дегидрлену реакциясы өтеді: 1-пируват дегидрленуі және 4 ЦЦ-да. Бұл реакцияларда түзілген 10 протон де-гидрогеназалар коферменттерін тотықсызданды-рады (НАДН2-лер) да олар тыныс алу тізбегінде тотығады. Тотыққан НАД-тар қайтадан кофер-менттік қызмет атқаруға пайдаланылады (катабо-лизмнің жалпы жолдарына. Коферменттердің де-генерациясы – бұл дегидрлену реакциясының өтуі үшін міндетті жағдай болып табылады. Сондықтан, катаболизмнің жалпы жолдары және тыны салу тізбегі өзара үздіксіз байланысқан болады және жеке қызмет етуі мүмкін емес. |
АТФ түзілуі көзқарасынан қарастырғанда, Кребс циклінің тотығуынан бөлінетін реакциялар НАДН-Н мен ФАДН2 тасымалдайтын электрондарда шоғырланған. Бұл қосылыстардағы «уақытша» ұсталған электрондар, жылдам арада тыныс алу тізбегіне, митохондрияның ішкі мембранасында орналасқан, ауысады.
Биологиялық тотығу.
Ағзадағы органикалық заттардың оттегімен су және СО2-ге дейін тотығуы процессі ұлпалық тыныс алу деп аталады. Ұлпалық тыныс алуға кіреді: а)субстраттан сутекті бөліп алу (дегидрлену); б)электрондардың оттегіге көп этапты тасымалдану процессі. Ұлпалық тыныс алу және тотығып фосфорлану (АТФ синтезі) энергиялық қосарланған процесс.
Тотығу процессін келесі теңдеумен беруге болады:
S Н2 + ½ О2 S + Н2О
Тотығушы әртүрлі органикалық заттар (S-субстраттар), катаболизмнің метаболиттері болып табылады, олардың дегидрленуі экзоэргиялық процесс. Тотығу реакциясында бөлінетін энергия толығымен жылу түрінде бөлінеді немесе жартылай АДФ-тың фосфорлануына жұмсалады.
Тотығып фосфорлану – бұл ұлпалық тыныс алу реакциясында бөлінген энергия есебінен АДФ пен Н3РО4 –тен АТФ-тің синтезделу реакциясы.
Хемиосмостық қосарлану процессі.
ХХ ғасырдың 60 жылдарының басында П.Митчелл бірнеше маңызды идеаларды постулат ретінде ұсынды.
1.Метаболиттің тотығуындағы энергия митохондрияда тотықсызданған эквиваленттер (Н,е-) түрінде болады, олар тыныс алу тізбегіне бағытталады. Мұнда электрондар редокс-градиенттер бойынша электрон ауыстырушылар арқылы соңғы реакцияда молекулалық оттегімен әсерлесіп су түзеді.
2.Редок-ауыстырушылар комплексі митохондрияның ішкі мембранасында жинақталған. Редок-градиенттің әртүрлі деңгейлеріне электрон ауыстырылуынан бөлінетін энергия протондарды матрикстен сорып шығаруға жұмсалады және митохондрияның ішкі мембранасында электрохимиялық потенциал түзіледі.
Хемиосмостық процесстің этаптарының жалпы сипаттамасы.
Хемиосмостық процесс метаболиттердің тотығуы энергиясын АТФ түзілуімен қосарландырады, ол митохондрияның ішкі мембранасында өтеді. Пируваттың лимон қышқылының циклінде тотығуы нәтижесінде түзілген энергия НАД+-пен ФАД-тардан НАДН-тан және ФАДН2 түзілуіне әкеледі немесе жоғары энергиялы электрондар мен тотықсыздандырушы эквиваленттер түрінде жинақталады.
Бұл электрондар молекуланың оттегімен байланысып оны активтендіріп Н2О түзілуіне және тотығып фосфорлану процесінде АТФ өндірілу үшін жұмсалады.
НАДН-н және ФАДН2 формасында резервтелген электрондар босатылады, митохондрияның ішкі мембранасында орналасқан тыныс алу тізбегінде тасымалданады.
Электрондардың бір комплекстен келесі комплекске өткенінде бөлінген энергия Н+ -ті матрикстен ішкі мембрана арқылы мембранааралық кеңістікке сорып шығарады.
Бұл процесс митохондрияның ішкі мембранасында электрохимиялық протондық градиенті қалыптастырады. Сондықтан, мембранааралық кеңістікке протондар концентрациясы жоғары болғандықтан олар протондық градиент бойынша кері матрикске ағылады (бір жағынан матрикстен еріс зарядтың да бағыттауымен). Бұл мембрана байланысқан АТФ-синтазаның (протондық каналдағы) қызметке әкеледі, нәтижесінде АДФ пен Н3РО4 –тен АТФ түзілуі өтеді.
Митохондриялық электрон тасымалдаушы тізбек (МЭТТ).
Алғашқы донор SH2 –ден электрондардың терминалдық акцептор О2-ге тасыфмалдану процесіне аралық жеткізушілер қатысады. Толық процесс тотығу-тотықсыздану реакцияларының тізбегін құрайды. Оның өту барысында аралық жеткізушілердің өзара әсерлесуі өтеді. Әрбір аралық жеткізушілер алғашқыда электрондар мен протондардың акцепторлары ролін атқарады. Электронды келесі жеткізушіге бергеннен кейін тотыққан қалыптан тотықсызданған формаға өтеді. Сонан кейін, ол электрондарды келесі жеткізушіге жеткізіп қайтадан тотыққан қалыпқа келеді. Соңғы стадияда жеткізуші электрондарды оттегіге береді, ал ол суға дейін тотықсызданады.
Тотығу-тотықсыздану реакцияларының кезектескен ретінің жиыны электрон тасымалдаушы тізбек немесе тыныс алу тізбегі деп аталады.
Жоғары сатыдағы ағзалардың тыныс алу тізбегіндегі аралық тасымалдаушылар ретінде келесі коферменттер болады: НАД+ (никотинамидадениндинуклеотид), ФАД және ФМН (флавинадениндинуклеотид және флавинмононуклеотид), кофермент Q (KoQ), гемқұрамды белоктар жиыны-цитохромдар (цх в, с1, с, а, а3) және гемсіз темірқұрамды белоктар (FeS, Fe2S2). Бұл тізбектегі барлық қатынасушылар төрт тотығу-тотықсызданушы жүйелерге немесе комплекске бөлінеді.