

Elektronnaya_metodichka_po_anatomii_3_modul
.pdfникогда не страдают. Чаще гидромиелия возникает в шейном отделе спинного мозга. Порок может быть как самостоятельным, так и сопровождать внутреннюю гидроцефалию, обусловленную атрезией срединной и латеральных апертур IV желудочка.
Дипломиелия – удвоение спинного мозга в области шейного или поясничного утолщения. Реже удваивается весь спинной мозг. Оба мозга лежат в одном ложе, состоящем из мягкой и твердой мозговых оболочек, местами соединяются с помощью глиальной ткани, довольно хорошо развиты и имеют все компоненты, характерные для спинного мозга. Порок крайне редкий, возникает до закрытия нервной трубки в результате раздельного смыкания каждой ее половины
Головной мозг
Наблюдается множество вариаций числа, формы и размеров борозд и извилин коры полушарий большого мозга. В лобной доле отмечено отсутствие следующих борозд: верхней лобной (1%), нижней лобной (16%), прецентральной (6%). Межтеменная борозда отсутствует в 2% случаев, постцентральная – в 25%, нижняя височная борозда – в 43% случаев. Многие борозды коры полушарий раздваиваются. Латеральная борозда раздваивается в задней своей части в 40% случаев, разделяется на 3-4 части – в 6% случаев. Верхняя и нижняя лобные борозды в 13% случаев соединяются в одну борозду. На нижней поверхности лобной доли иногда определяется надглазничная поперечная борозда. Постцентральная борозда иногда сливается с межтеменной бороздой, задней частью латеральной борозды (в 31% случаев). В 56% случаев над поясничной бороздой проходит одноименная параллельная ей борозда. В 40% случаев имеется дополнительная дуговая борозда предклинья. Верхняя затылочная борозда бывает раздвоена (в 55%) или утроена (в 12%) в виде двух-трех поперечных борозд. Средняя височная борозда иногда замещена несколькими радиальными или расходящимися бороздами.
Иногда обонятельный тракт имеет в себе продольный канал. Изредка между областью расхождения ножек свода и валиком мозолистого тела имеется небольшая уплощенная четырехугольная и закрытая снизу щель (треугольная щель), основание этой щели обращено кпереди.
Варьируют размеры и форма таламуса - редко наблюдаются два межталамических сращения. Размеры сосцевидных тел варьируют. Конфигурация, взаимоотношения гипоталамических ядер, их размеры изменчивы. Глубина межножковой ямки, количество отверстий заднего продырявленного вещества варьируют. Наблюдаются вариации протяженности и размеров черного вещества и красного ядра. Глубина базилярной борозды моста может быть различная. Форма моста, толщина средних ножек мозжечка индивидуально изменчивы. Наблюдается одностороннее или двухстороннее отсутствие слуховых полосок, косой или латеральный их ход на поверхности продолговатого мозга. Количество извилин мозжечка варьирует от 127 до 244. Латеральнее передней поверхности нижнего отдела червя может наблюдаться маленькая добавочная долька-пирамидка. Описаны дополнительные клочки мозжечка, фиксированные к червю самостоятельными ручками.
Пороки конечного мозга в результате несмыкания нервной трубки – дизрафии краниальной области.
В основе пороков этой группы лежит нарушение развития эктодермального и мезодермального листков, вследствие чего такие пороки часто сопровождаются нарушениями развития мозговых оболочек, костей черепа и мягких покровов головы.
Анэнцефалия – отсутствие большого мозга, костей свода черепа и мягких тканей. Часто повреждается и задний мозг. На месте мозгового вещества обычно располагается богатая кровеносными сосудами соединительная ткань с кистозными полостями, выстланными медуллярным эпителием, глия, единичные нервные клетки, остатки сосудистых сплетений. Анэнцефалия, как правило, сопровождается выраженной гипоплазией надпочечников и аплазией нейрогипофиза.
43

IV. ПРОВОДЯЩИЕ ПУТИ ГОЛОВНОГО И СПИННОГО МОЗГА
Проводящие пути – цепь нейронов, соединяющих функционально однородные участки серого вещества в ЦНС, занимающих в белом и сером веществе головного и спинного мозга определенное место и проводящих одинаковый импульс.
Проводящие пути являются частью сложных рефлекторных дуг, которые соединяют между собой различные отделы центральной нервной системы и обеспечивают двухстороннюю функциональную связь между отдельными структурами головного и спинного мозга. Они отличаются многочисленностью, сложностью строения и надежностью функционирования.
Взависимости от величины, формы и направления нервного импульса проводящие пути полу-
чают название: путь (tractus), пучок (fasciculus), волокна (fibrae), спайка (commissura), петля (lemniscus) или лучистость (radiatio).
Все проводящие пути ЦНС подразделяют на три группы: 1. Проекционные.
2. Комиссуральные.
3. Ассоциативные.
Впроцессе становления проводящей системы в онтогенезе первоначально формируются проекционные пути, а затем комиссуральные и ассоциативные.
Ассоциативные проводящие пути соединяют участки коры в пределах одного полушария. Различают:
А. Короткие волокна, дугообразные волокна, fibrae arcuatae cerebri, которые соединяют корковые поля соседних извилин.
Б. Длинные волокна, которые соединяют корковые поля отдаленных извилин (верхний про-
дольный пучок, fasciculus longitudinalis superior, нижний продольный пучок, fasciculus longitudinalis inferior, пояс, cingulum, крючковидный пучок, fasciculus uncinatus).
Рис.12. Проводящие пути.
1- ассоциативные, 2 – комиссуральные, 3 – проекционные.
Коммиссуральпые пути соединяют симметричные части правого и левого полушарий. К ним принадлежат:
1.Мозолистое тело, corpus callosum.
2.Передняя спайка, commissura anterior. Относится к обонятельному мозгу.
3.Спайка свода, commissura hippocampi (fornicis). Соединяет корковые поля гиппокампа правого и левого полушарий.
4.Задняя спайка, comissura posterior.
44
5.Спайка поводков, comissura habenulare.
6.Межталамическое сращение, adhesio interthalamica.
Проекционные пути соединяют кору с подкорковыми образованиями головного мозга и спинным мозгом.
Проекционные проводящие пути связывают кору головного мозга с его нижележащими отде-
лами (короткие проводящие пути) и со спинным мозгом (длинные проводящие пути).
По направлению проведения нервного импульса проекционные пути подразделяют на две группы:
•афферентные (восходящие, центростремительные, чувствительные), которые проводят нервный импульс от рецепторов, воспринимающих информацию из внешнего мира или внутренней среды организма к различным отделам головного мозга и к коре полушарий;
•эфферентные (нисходящие, центробежные, двигательные), передающие импульс от коры головною мозга и других его отделов на периферию.
ЧУВСТВИТЕЛЬНЫЕ (АФФЕРЕНТНЫЕ) ПРОВОДЯЩИЕ ПУТИ
Под чувствительностью понимают способность организма воспринимать воздействия раздражителей внешней и внутренней среды.
Вид чувствительности определяется характером рецептора, воспринимающего раздражение. Среди рецепторов различают экстерорецепторы (тактильная, болевая, температурная), располагающиеся в коже и слизистых оболочках.; проприорецепторы (мышечно-суставная, вибрационная, чувство давления и веса), которые находятся в мышцах, сухожилиях, связках, суставных капсулах; и висцерорецепторы (чувствительность внутренних органов и сосудов), расположенные в различных внутренних органах и сосудах.
В зависимости от вида чувствительности афферентные проводящие пути подразделяют на:
•экстероцептивные (контактные – общей чувствиельности и дистантные – видовой чувствительности,
•проприоцептивные (корковые – сознательные и мозжечковые – бессознательные),
•интероцептивные.
Те из них, которые заканчиваются в коре полушарий большого мозга, получили название проводников сознательной чувствительности, или проводящих путей коркового направления.
Для чувствительных путей коркового направления характерно:
1.Наличие 3-х нейронов.
2.I-е нейроны представлены псевдоуниполярными нейронами, тела которых расположены в спинно-мозговом узле, а периферические отростки образуют рецепторы.
3.Тела II-х нейронов располагаются в ядрах спинного или продолговатого мозга.
4.Аксоны II-х нейронов образуют (как правило) перекрест.
5.Большинство аксонов II-х нейронов идут к зрительному бугру в составе медиальной петли.
6.Волокна этих путей располагаются, как правило, в дорсальных отделах ствола мозга.
7.Тела III-х нейронов располагаются в латеральных отделах зрительного бугра.
8.Все они проходят в средней трети задней ножки внутренней капсулы.
9.Аксоны III-х нейронов по пути в кору образуют или проходят в составе лучистого венца.
ЧУВСТВИТЕЛЬНЫЕ ПРОВОДЯЩИЕ ПУТИ КОРКОВОГО НАПРАВЛЕНИЯ Первые (рецепторные) нейроны чувствительных проводящих путей вынесены за пределы го-
ловного и спинного мозга в чувствительные ганглии. Это спинномозговые узлы или чувствительные ганглии черепных нервов.
Tractus spino-bulbo-thalamo-corticalis – чувствительный, сознательный, 3-хнейронный, полностью перекрещенный, проводит проприоцептивную чувствительность от мышц, суставов и связок. От кожи проводит тактильное чувство (чувство стереогнозии – узнавание предметов на ощупь). 1-й нейрон располагается в спинномозговом узле и представлен ложноуниполяоной клеткой: ее дендрит в составе спинномозгового нерва идет на периферию и доходит до кожи, мышц, суставов, связок, где заканчивается рецептором. Аксон 1-гонейрона идет в составезадних корешков, ноневступает в серое вещество задних рогов, а идет в задних канатиках спинного мозга. Аксоны нижних 19 сегментов спинного мозга образуют путь Голля – тонкий – fasciculus gracilis, он расположен медиально, а от верхних 12 сегментов – путь Бурдаха – клиновидный – fasciculus cuneatus. Эти пути идут в продолговатый мозг, где заканчиваются в одноименных ядрах, которые являются вторыми нейронами. Отро-
45
стки (аксоны) вторых нейронов делают в межоливном слое продолговатого мозга полный перекрест
(decussatio lemniscorum) и далее образуют медиальную петлю lemniscus medialis (tractus bulbothalamicus). Волокна медиальной петли идут через мост, средний мозг, где располагаются в покрышке, латеральнее красного ядра, и заканчиваются в клетках латерального ядра (nucleus lateralis) зрительного бугра (thalamus) промежуточного мозга. Здесь располагается тело 3-го нейрона. Из зрительного бугра отростки третьих нейронов через заднюю ножку внутренней капсулы направляются к коребольших полушарий (корковыеконцы анализаторов) в виде3-хпучков:
1-й подходит к передней центральной извилине и заканчивается в дольке Беца (проводит проприоцептивную чувсвительность);
2-й заканчивается в постцентральной извилине (проводит тактильную чувствительность от кожи).
3-й заканчивается в верхней теменной дольке (проводит чувство стереогнозии).
Tractus spino-thalamo-corticalis - чувствительный, сознательный, 3-хнейронный, полностью перекрещенный, проводит эекстероцептивную чувствительность от кожи и видимых слизистых.
Тела I-х нейронов располагаются в спинномозговых узлах. Периферические отростки псевдоуниполярных клеток подходят к рецепторам кожи туловища и конечностей, а центральные отростки в составе задних корешков входят в задние рога серого вещества, где проводники тактильной чувствительности переключаются на клетках студенистого вещества; проводники болевой и температурной чувствительности переключаются на клетках собственногоядра заднегорога. Аксоны II-х нейронов от этих ядер переходят на противоположную сторону вступают в боковые и передние канатики белого вещества. Аксоны клеток собственного ядра заднего рога проходят в боковых канатиках, составляя tr. spino-thalamicus lateralis. Аксоны клеток студенистого вещества проходят в передних канатиках, составляя tr. spino-thalamicus anterior. Пройдя через спинной мозг, продолговатый мозг, входят в состав медиальной петли; в ее составе проходят покрышку моста, покрышку ножек мозга и заканчиваются в латеральном ядре таламуса, где лежат тела III-х нейронов этого пути. Их аксоны составляют tr. thalamo-corticalis, который проходит в задней ножке внутренней капсулы и заканчивается в коре постцентральной извилины (проводники стереогноза – в верхней теменной дольке). Часть проводников тактильной чувствительности проходит также в составе задних канатиков спинного мозга, вместе с проприоцептивными путями.
Медиальная петля образована аксонами вторых нейронов проводящих путей проприо- и экстероцептивной чувствительности противоположной стороны.
ПРОПРИОЦЕПТИВНЫЕ ПУТИ КМОЗЖЕЧКУ
Проприоцептивные пути к мозжечку составляют часть рефлекторного аппарата, осуществляющего функцию равновесия (бессознательную координацию движений). Они проводят бессознательное мышечно-суставное чувство от рецепторов аппарата движения, участвуют в регуляции мышечного тонуса.
Tractus spino-cerebellaris anterior (Говерса).
Чувствительный, бессознательный, 2-хнейронный, дважды перекрещенный. Проводит проприоцептивную чувствительность от мышц, суставов и связокк мозжечку.
1-й нейрон находится в спинномозговом ганглии и представлен псевдоуниполярной клеткой. Дендриты идут на периферию в составе спинномозгового нерва и заканчиваются рецепторами в мышцах, суставных сумках и связках. Аксоны в составе задних корешков идут в спинной мозг, в промежуточную зону и заканчиваются в nucleus intermediomedialis. Здесь располагаются вторые нейроны. Аксоны вторых нейронов полностью перекрещиваются и переходят на противоположную сторону в составе comissura alba anterior. Затем они заходят в верхний мозговой парус и здесь делают второй перекрест, возвращаясь на свою сторону. Таким образом, путь становится дважды перекрещенным. Через верхниеножки он заходит в мозжечок и заканчивается в коречервя.
Tractus spinocerebellaris posterior(Флексига).
Чувствительный, 2-хнейронный, бессознательный, неперекрещенный путь. Проводит бессознательную проприоцептивную чувствительность от мышц, суставов, связок. 1-й нейрон располагается в спинномозговом ганглии и представлен псевдоуниполярной клеткой. Дендриты идут на периферию в составе спинномозговых нервов, а аксоны в составе задних корешков заходят в задние рога спинного мозга и заканчиваются в nucleus thoracicus (Кларка-Штилинга). Здесь лежит второй нейрон. Отростки 2-го нейрона идут в боковых канатиках на своей стороне, поднимаются вверх, заходят в продолговатый мозг и через нижниеножки мозжечка заходят в мозжечок, заканчиваясь в коре червя. Оба эти
46
пути (posterior и anterior) участвуют в поддержании равновесия, сохранении мышечного тонуса, мышечной координации, преодолении инерции и силы тяжести.
Кроме описанных выше путей специфической чувствительности существуют пути неспецифической чувствительности, которые связаны с ретикулярной формацией. Все специфические чувствительные пути, проходя к коре больших полушарий, отдают коллатерали к ядрам ретикулярной формации, в которой чувтсивтельные импульсы утрачивают свою специфичность и следуют в кору лишь в качестве энергетического компонента, вызывая ее диффузное возбуждение. Эти импульсы тонизируют коруи делают ее способной к восприятию специфических нервных импульсов.
ДВИГАТЕЛЬНЫЕ(ЭФФЕРЕНТНЫЕ) ПРОВОДЯЩИЕ ПУТИ
Эфферентные пути проводят нервный импульс из различных центров головного мозга к рабочему органу. У человека прямые эфферентные пути начинаются лишь из коры больших полушарий, где располагаются тела так называемых центральных нейронов. Эти пути являются проводниками сознательных двигательных импульсов к скелетной мускулатуре и объединяются понятием пирамидная система. Промежуточные ядра ствола (красное ядро, ядра крыши среднего мозга, ядра ретикулярной формации, ядро оливы, латеральное преддверное ядро) рассматриваются как начало эфферентных проводящих путей, которые посылают бессознательные двигательные импульсы на скелетную мускулатуру и входят в состав экстрапирамидной системы. Мозжечок осуществляет связь со спинным мозгом через указанные стволовые ядра. Все нисходящие пути заканчиваются на телах и дендритах нейронов двигательных ядер передних рогов серого вещества спинного мозга или двигательных ядер черепных нервов (периферические нейроны).
Таким образом, всеэфферентныепроводящиепути подразделяются на двегруппы:
1.Пирамидныепути.
2.Экстрапирамидныепути.
ПИРАМИДНЫЕПУТИ
Пирамидные пути проводят сознательные (волевые) двигательные импульсы, а также тормозные импульсы от коры полушарий большого мозга к нейронам двигательных ядер черепных нервов и к нейронам двигательных ядер передних рогов сероговещества спинногомозга.
Для пирамидных путей характерно:
1.Наличие2-х нейронов.
2.I-й нейрон – пирамидныеклетки Беца (Vслой коры предцентральной извилины).
3.В полушарии головного мозга волокна проходят в составе лучистого венца и внутренней капсулы, занимая коленои передние2/3 задней её ножки.
4.В мозговом стволе волокна следуют в его вентральной части, переходя последовательно из ножек мозга в мост и продолговатый мозг.
5.На границе со спинным мозгом 80% волокон, переходя на другую сторону, образуют нижний двигательный перекрест (decussatiopyramidum).
6.В спинном мозгепирамидныепути занимают егопередниеи боковые канатики.
7.Тела II-х нейронов располагаются в двигательных ядрах передних рогов спинного мозга или в двигательных ядрах черепных нервов.
В зависимости от места назначения пирамидныепути подразделяются на двегруппы:
• корково-ядерный путь, tractus corticonuclearis (corticobulbaris);
• корково-спинномозговой путь, tractus corticospinalis (pyramidalis) .
Tractus corticonuclearis (corticobulbaris).
Это двигательный, сознательный, частично перекрещенный путь. Проводит двигательные сознательные импульсы от коры головного мозга к мышцам головы, лица и шеи через ядра черепномозговых нервов.
1-й нейрон расположен в прецентральной извилине и дольке Беца (lobulus paracentralis). Он представлен пирамидными клетками, находящимися в V, VI слоях коры. Дендриты участвуют в образовании внутримозговых связей, а аксоны идут в белое вещество головного мозга, и через колено внутренней капсулы выходят из полушарий. Затем они идут в составе ножек мозга, моста и продолговатогомозга. Волокна заканчиваются в стволе мозга на двигательных ядрах черепных нервов. Ковсем ядрам кроме ядер VII и XII пар подходят частично перекрещенные волокна, а к ядрам этих черепных нервов подходят волокна, толькос противоположной стороны (т.е. полностью перекрещенные).
2-ые нейронырасположены в ядрах ЧН:
47

В среднем мозге находятся ядра:
III пары – nucleus motorius n. oculomotorius; IVпары – nucleus motorius п. trochlearis;
В мостуядра:
Vпары – nucleus motorius n. trigemini; VI пары – nucleus motorius n. abducentis; VII пары – nucleus motorius n. facialis;
В продолговатом мозге ядра:
IX, X, XI пары –nucleus ambiquus;
XI пары – nucleus motorius п. accessorii; XII пары – nucleus motorius n. hypoglossi.
Отростки 2-х нейронов идут на периферию в составе черепных нервов и подходят к поперечнополосатым мышцам лица, щек, головы, на которых образуют двигательные бляшки.
Рис. 13. Нисходящие пути спинного и головного мозга (полусхематично).
1 - neurofibrae commissurales, 2 - fibrae corticostriatae, 3 - radiationes thalamicae, 4 - tractus corticotegmentalis, 5 - fibrae striopallidae, 6 - ventriculus tertius, 7 - ansa et fasciculus lenticulares, 8 - fibrae rubrothalamicae, 9 - pedunculus cerebri, 10 - tractus cerebellobulbaris, 11 - nuclei nervorum cranialium, 12 - nucleus dentatus, 13 – crebellum, 14 – pons, 15 – oliva, 16 - tractus rubrospinalis, 17 - tractus corticospinalis (pyramidalis) lateralis, 18 - tractus corticospinalis (pyramidalis) anterior, 19 - medulla oblongata, 20 – pyramis, 21 - nucleus ruber, 22 - fibrae corticonucleares, 23 – thalamus, 24 - fibrae corticorubralis, 25 - nucleus lentiformis, 26 - capsula interna, 27 – claustrum, 28 - fibrae corticospinales, 29 -nucleus caudatus, 30 – cortex, 31corona radiata, 32corpus callosum.
Tractus corticospinalis (pyramidalis).
Сознательный, двигательный, двухнейронный, полностью перекрещенный. Проводит сознательные импульсы от корыкпоперечно-полосатыммышцам.
1-й нейрон расположен в коре в передней центральной извилине и в дольке Беца и является пирамидной клеткой V, VI слоя. Дендриты формируют в коре синапсы, а аксон идет в белое вещество полушарий и выходит через переднюю часть задней ножки внутренней капсулы. Затем он идет в основании ножек мозга. По базальной части Варолиева моста входит в продолговатый мозг, идет в пирамиды продолговатого мозга. На границе со спинным мозгом делает частичный перекрест (decussatio pyramidum)
иделитсяна 2пути:
1)перекрещенныеволокнаидут в боковыхканатиках, образуяtr. corticospinalis lateralis;
2)неперекрещенныеволокна идут в передних канатиках, образуяtr. corticospinalis anterior. Оба пути идут в передниерога, и заканчиваютсяна клетках двигательных ядер, которыеявляются 2-ми нейро-
48
нами. Неперекрещенные волокна (tr. corticospinalis anterior) делают перекрест в белой спайке comissura alba. Таким образом, путь стал полностью перекрещенным. Аксоны 2-х нейронов выходят из спинного мозга передними корешками, а затем в составе спинномозговых нервов идут на периферию и заканчиваютсядвигательными бляшками в мышцах.
ПУТИЭКСТРАПИРАМИДНОЙСИСТЕМЫ
Экстрапирамидная система является более старой по сравнению с пирамидной двигательной системой. У нижнихпозвоночных(урыб)экстрапирамидная система является ведущей, обеспечиваяих относительно простые движения. Затем, по мере развития переднего мозга и дифференцирования коры полушарий, у животных образуется новая, пирамидная, корковая система, она соответствует новой форме двигательных актов, все более ограничивающихся определенной группой мышц. Экстрапирамидная же система у человека обеспечивает тонус мышц, состояние готовности их к сокращению, автоматические движения.
Главным центром экстапирамидной системы является полосатое тело (striatum et pallidum). На пути к спинному мозгу волокна от этих ядер переключаются на промежуточных центрах: красном ядре, черном веществе, подталамическом ядре, зубчатом ядре мозжечка и т.д. От striatum (хвостатое ядро и скорлупа чечевицеобразного ядра) волокна следуют к pallidum (globus pallidus) это стрио-паллидарные во-
локна. От pallidum идут: tr. pallido-rubralis (к красному ядру), tr.pallido-nigralis (к черному веществу), tr.pallido-subthalamicus (к n.subthalamicus, corp.Luisii) Oт nucl. subthalamicusследует путь к красномуядру
(th. subthalamico-rubralis). Черное вещество, повидимому, также связано с красным ядром. Т.о. красное ядро – ведущий промежуточный центр экстрапирамидной системы. От него идет tr. rubrospinalis (путь Монакова)кспинномумозгу.
При поражении различных центров экстрапирамидной системы у человека наблюдаются гипокинезии либогиперкинезы, скованностьдвижений, дрожательный паралич,паркинсонизм.
Чувствительные импульсы передаются на полосатое тело через таламус: tr.thalamo-strialis, tr. thalamo-pallidalis, которые идут от медиальногоядра таламуса. Через таламус на экстрапирамидную систему передается влияние коры б.п. (tr. corticothalamicus – к медиальному ядру таламуса, а от него tr. thalamostrialis, tr. thalamopallidalis.
Tractus rubrospinalis (Монакова).
Двигательный, бессознательный, полностью перекрещенный. Проводит бессознательные импульсы от красного ядра ко всем поперечно-полосатым мышцам. В красное ядро собираются волокна от всей экстрапирамидной системы(полосатоетело, чернаясубстанция, зубчатоеядро).
1-й нейрон располагается в красном ядре покрышек ножек мозга. Аксоны здесь полностью перехо-
дят на противоположную сторону – полный передний покрышечный перекрест (Фореля), decussatio tegmentalis anterior. После перекреста волокна заходят в мост, продолговатый мозг, а затем в боковые канатики спинного мозга. В стволе мозга от пути отходят волокна к двигательным ядрам черепных нер-
вов (III, IV, V, VI, VII, IX, X, XI, ХIIпары).
В спинном мозге волокна посегментно заходят в передние рога и заканчиваются на клетках двигательных ядер. Таким образом, вторые нейроны расположены как в двигательных ядрах черепных нервов, таки в ядрахпереднихрогов спинногомозга.
Аксоны вторыхнейронов идут в составечерепныхнервов и спинно-мозговых нервов на периферию к поперечно-полосатым мышцам. По этому пути осуществляются автоматические движения, бессознательные(идтина красномядре, наподкорке).
Tractus tectospinalis.
Это двигательный, бессознательный, двухнейронный, полностью перекрещенный. Проводит бессознательные двигательные импульсы от верхних и нижних бугорков пластинки четверохолмия (подкорковыецентры слуха и зрения) ковсем поперечно-полосатым мышцам. 1-енейронырасполагаются в бугорках пластинки четверохолмия. Их аксоны здесь же переходят на противоположную сторону – зад-
ний покрышечный перекрест (Мейнерта), decussatio tegmeni posterior. Волокна идут в мост, продол-
говатый мозг, передние столбы спинного мозга, заканчиваются на клетках двигательных ядер передних рогов. Попути часть волокон заходит в двигательныеядра черепных нервов. Таким образом, 2-е нейроны представлены как клетками двигательных ядер черепных нервов, так и передних рогов спинного мозга. В составе черепных нервов и спинномозговых нервов отростки 2-х нейронов заканчиваются двигательными бляшками на поперечно-полосатых мышцах. По этому пути осуществляются бессознательныедвигательныереакции на световыеи звуковыераздражения(сигналмашины, фотовспышка).
49
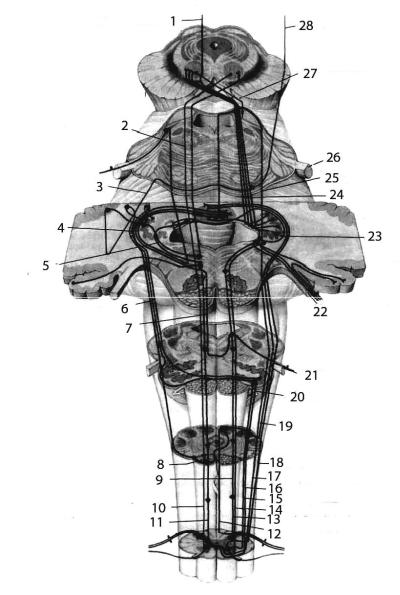
Рис. 14. Восходящие и низходящие пути ствола головного мозга и мозжечка.
1 - tractus dentatothalamicus, 2 - tractus rubrospinalis, 3 - fibrae pontocelebellares, 4 - tractus spinalis n. trigemini, 5 - мозжечково-ядерный путь, 6 - tractus olivocerebellaris, 7 -
ядерно-мозжечковый путь, 8 - nuclei arcuati, 9 - tractus bulboreticulospinalis, 10 - tractus rubrospinalis, 11 - tractus pontoreticulospinalis, 12 - fasciculi gracilis et cuneatus, 13 - tractus bulboreticulospinalis, 14 - tractus vestibulospinalis, 15 - tractus olivospinalis, 16 - tractus spinoolivaris, 17 - tractus spinocerebellaris anterior, 18 - tractus spinocerebellaris posterior, 19 -
волокна от тонкого и клиновидного пучков к мозжечку, 20 - n. Hypoglossus, 21 - n. Vagus, 22 - n. Vestibulocochlearis, 23 - tractus vestibulocerebellaris, 24 - tractus cerebellorubralis, 25 - tractus spinocerebellaris anterior, 26 - n. Trigeminus, 27 - tractus rubrospinalis, 28 - fibraecorticopontinae.
Tractus vestibulospinalis.
Бессознательный, двигательный, 2-х нейронный, полностью перекрещенный. Проводит двигательные импульсы от латерального вестибулярного ядра к поперечно-полосатым мышцам. 1-й нейрон располагается в латеральном вестибулярном ядре. Волокна делают полный перекрест в продолговатом мозге и идут в белом веществе боковых и передних канатиков спинного мозга. Заканчиваются посегментно на клетках двигательных ядер спинного мозга (2-е нейроны). От них в составе передних корешков, потом спинно-мозговых нервов волокна идут к поперечнополосатым мышцам. По этому пути осуществляется координация движений сохранениеположениятела впространстве.
СВЯЗЬ КОРЫ ГОЛОВНОГО МОЗГА С КОРОЙ МОЗЖЕЧКА
1). Лобно-мосто-мозжечковый путь – tractus fronto-ponto-cerebellaris; 2). Височно-мосто-мозжечковый – tractus temporo-ponto-cerebellaris; 3). Затылочно-мосто-мозжечковый – tractus occipito-ponto-cerebellaris.
1-й нейрон находится в лобной, височной, затылочной долях соответственно. Дальше волокна идут через внутреннюю капсулу, базальную часть ножек мозга. Заходят в базальную часть моста, где делают полный перекрест и заканчиваются на nuclei proprii. Это 2-й нейрон. Отростки 2-го нейрона проходят в составесредних мозжечковых ножек идут в корумозжечка, гдеи заканчиваются.
50
V. ПЕРИФЕРИЧЕСКИЙ ОТДЕЛ СОМАТИЧЕСКОЙ НЕРВНОЙ СПИННОМОЗГОВЫЕ НЕРВЫ
Учитывая, что центральными отделами соматической (анимальной) нервной системы являются спинной и головной мозг, то соответственно периферическими составляющими этих отделов выступают спинномозговые и черепные нервы.
ФИЛОГЕНЕЗ СПИННОМОЗГОВЫХ НЕРВОВ
У низших животных, например ланцетника, передний и задний корешки спинного мозга представляют самостоятельные ветви, не образуют смешанного спинномозгового нерва. У рыб задний и передний корешки соединяются вне позвоночного канала, хотя спинномозговой узел заднего корешка располагается в этом канале. Спинномозговые нервы водных животных иннервируют соответствующие миотомы и дерматомы, которые были связаны с метамерами тела, послужившими зачатками при образовании конечностей. У акул, селяхий и других водных животных формируется объединенное шейно-плечевое и пояснично-крестцовое сплетения. Только у амфибий отмечаются самостоятельные шейное и плечевое, поясничное и крестцовое сплетения. Даже у пресмыкающихся, которые лишены конечностей, закладываются шейное и плечевое сплетения. Самостоятельные поясничные и крестцовые сплетения возникают урептилий. Этот факт свидетельствует, чтопредки данных видов животных имели конечности, которые с изменением образа жизни утратили функцию и атрофировались. У млекопитающих принцип строения периферической нервной системы во многом имеет общие черты с таковым учеловека.
ОНТОГЕНЕЗ СПИННОМОЗГОВЫХ НЕРВОВ
Центральная нервная система и двигательные волокна развиваются из нервной трубки, чувствительные волокна — из нейробластов ганглионарных пластинок. В составе нервной трубки и ганглионарных пластинок имеются нейробласты—молодые неразвившиеся нейроны, способные к миграции. На 3-4-й неделе внутриутробного развития отростки нейробластов, находящихся в вен- тро-латеральном отделе нервной трубки, отделяются от нее и образуют передние корешки спинномозговых нервов. Эти корешки врастают в рядом лежащие миотомы. Нейробласты двух ганглионарных пластинок посылают отростки в двух направлениях: одни — к спинному мозгу, образующие задние корешки спинномозговых нервов, другие — на периферию тела или к внутренним органам {пищеварительная, дыхательная, мочеполовая, эндокринная и сердечно-сосудистая системы), имеющим рецепторы. Первоначально отростки передних и задних корешков располагаются самостоятельно и только на 5-6-й неделе внутриутробного развития сливаются в спинномозговой нерв, который находится в анатомической и функциональной связи с соответствующим склеротомом, дерматомом и миотомом. В это же время отмечается перемещение миотомов туловища и конечностей, что приводит к переплетению нервов и формированию нервных сплетений. На 4- 5-м месяце эмбрионального развития из клеток эктодермы формируются миелиновые оболочки. Миелинизация заключается в том, что клетки энтодермы, вытягиваясь около нервных волокон, формируют нейролемму, где откладывается миелин. В первую очередь миелинизиру-ются передние и задние корешки верхних отделов спинного мозга, черепные нервы и те проводящие пути центральной нервной системы, которые имеют более филогенетически древнее происхождение, например tractus rubrospinalis. Волокна пирамидного пути покрываются миелиновыми оболочками только после рождения.
Передние корешки спинного мозга образованы аксонами мотонейронов передних рогов спинного мозга, следовательно, являются двигательными. Задние корешки образованы центральными отростками клеток спинномозговых узлов и являются чувствительными. Периферические отростки клеток спинномозговых узлов начинаются рецепторами на периферии.
Передние и задние корешки сближаются друг с другом и после узла в межпозвоночном отверстии образуют смешанный спинномозговой нерв (n. spinalis). Общее число спинномозговых нервов - 31 пара: 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1 копчиковый (соответствуют сегментам спинного мозга). По выходе из отверстия каждый нерв делится на четыре ветви. Две из них: передняя (r. ventralis) и задняя (r. dorsalis) являются продолжением нерва; другие две: менингеальная идет к оболочкам спинного мозга, а соединительная (r. communicantes) идет на соединение с узлом симпатического ствола.
Воспаление переднего или заднего корешков спинного мозга называется радикулит; ствола СМН – фуникулит; периферических ветвей – неврит.
51

Рис. 15. Схема основных чувствительных и двигательных нейронов спинномозговых нервов и их связей со спинным мозгом.
1- кожа, 2- нервные волокна от кожных рецепторов, 3- задняя ветвь, 4- передняя ветвь, 5- скелетная мышца, 6- ганглий заднего корешка, 7- вегетативная ветвь, 8- симпатический ганглий, 9- передний корешок, 10кишечник.
Задние ветви всегда тоньше соответствующих передних. Каждая из задних ветвей (за исключением I шейного нерва) делится на медиальную и латеральную ветви. Задниешейные ветви иннервируют кожу и мышцы спины, расположенные в области шеи. Первая задняя ветвь - двигательная и называется подзатылочным нервом (n. suboccipitalis), который иннервирует короткие затылочно-позвоночные мышцы (большую и малую задние прямые мышцы головы, верхнюю и нижнюю косые). 2-я задняя ветвь самая значительная из всех задних шейных ветвей. Ее медиальная кожная ветвь прободает сухожилиетрапециевидной мышцы и появляется под кожей, как большой затылочный нерв (n. occipitalis major) и иннервирует кожу затылочной области. Остальные задние шейные ветви очень короткие, иннервируют кожуи аутохтонныемышцы спины.
Задние ветви грудных и поясничных нервов также иннервируют кожу и собственные мышцы спины. Ветви от верхних поясничных нервов достигают кожи ягодичной области и называются верхними нервами ягодиц (nn. clunium superiores). Задние ветви крестцовых нервов очень тонкие, выходят через задние крестцовые отверстия и образуют средние ветви ягодиц (nn. clunium medii). Задняя ветвь копчикового нерва иннервирует кожу в области копчика и анального отверстия.
Менингеальные ветви возвращаются через межпозвоночные отверстия в позвоночный канал и иннервируют оболочки и зубчатые связки спинного мозга. Они состоят из чувствительных и симпатических волокон.
Соединительные ветви вступают в нерв из узлов симпатического ствола и состоят из симпатических волокон, которые обеспечивают трофику скелетных мышц и иннервацию кожи.
Передние ветви спинномозговых нервов иннервируют кожу и мышцы передней стенки туловища и конечности. Их волокна в отличие от задних ветвей переплетаются и образуют нервные сплетения, в которых происходит обмен волокон из различных сегментов. Различают четыре больших сплетения: шейное, плечевое, поясничное и крестцово-копчиковое. Лишь передние ветви 12 грудных нервов не образуют сплетений. Они вместе с артериями и венами располагаются в межреберных промежутках, между наружными и внутренними межреберными мышцами, у нижнего края вышележащего ребра, в его борозде и называются межреберными.
Первый межреберный нерв тонкий, т.к. большая часть этой ветви входит в состав плечевого сплетения; XII нерв лежит под соответствующим ребром и называется подреберным нервом. Шесть пар верхних межреберных нервов по межреберным промежуткам достигают грудины, а шесть нижних переходят на переднюю брюшную стенку, располагаются там между поперечной и внутренней косой мышцами живота и проникают во влагалище прямых мышц живота. Межреберные нервы иннервируют собственныемышцы груди, всемышцы живота, кожугруди (молочную железу) и живота.
52