

201403170953311
.pdfМИНИСТЕРСТВО ЗДРАВООХРАНЕНИЯ РЕСПУБЛИКИ БЕЛАРУСЬ
БЕЛОРУССКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ
1-я КАФЕДРА ДЕТСКИХ БОЛЕЗНЕЙ
А. В. СОЛНЦЕВА, А. В. СУКАЛО
ДИАГНОСТИКА НЕДОСТАТОЧНОСТИ ГОРМОНА РОСТА У ДЕТЕЙ
Учебно-методическое пособие
Минск 2007
УДК 612.433.65.09–053.2 (075.8) ББК 57.31 я 73
С 16
Утверждено Научно-методическим советом университета в качестве учебно-методического пособия 14.06.2006 г., протокол № 7
Р е ц е н з е н т ы: канд. мед. наук, доц., зав. каф. пропедевтики детских болезней Белорусского государственного медицинского университета В. И. Твардовский; канд. мед. наук, доц. каф. эндокринологии Белорусской медицинской академии последипломного образования В. И. Шутова
Солнцева, А. В.
С 16 Диагностика недостаточности гормона роста у детей : учеб.-метод. пособие / А. В. Солнцева, А. В. Сукало. – Минск: БГМУ, 2007. – 40 с.
ISBN 985–462–622–9.
Обобщены современные представления о физиологической роли гормона роста, отражены сведения об особенностях периодов роста ребенка в зависимости от пола и возраста, аспектах техники антропометрии, оценки физического развития, инструментальной и лабораторной диагностики соматотропной недостаточности в детской практике.
Предназначено для студентов педиатрического факультета, врачей-стажеров, педиатров, эндокринологов, врачей общей практики.
|
УДК 612.433.65.09–053.2 (075.8) |
|
ББК 57.31 я 73 |
ISBN 985–462–622–9 |
© Оформление. Белорусский государственный |
|
медицинский университет, 2007 |
2
СПИСОК СОКРАЩЕНИЙ
ГР — гормон роста ГР-РГ — гормон роста-рилизинг гормон
ИФР-1 — инсулиноподобный фактор роста-1 ИФР-2 — инсулиноподобный фактор роста-2 АКТГ — адренокортикотропный гормон
ИФРСБ — инсулиноподобный фактор роста связывающий белок
Рост является одним из показателей, характеризующих состояние здоровья ребенка. Он регулируется такими факторами как питание, эмоциональное состояние, реактивность тканей и скелета, влияние окружающей среды, генетическая предрасположенность и гормональный фон.
РЕГУЛЯЦИЯ СИНТЕЗА И СЕКРЕЦИИ ГР
Ген, ответственный за синтез ГР, локализован на длинном плече 17-й хромосомы (17g22g24). Он кодирует аминокислотную последовательность прогормона, который в результате протеолиза превращается в активные формы ГР. Молекула ГР — одноцепочечный полипептид с двумя поперечными дисульфидными связями между цистеиновыми группами. Более 90 % секретируемого ГР представлено преимущественно в форме 22К–ГР с молекулярной массой 21 500 дальтон. Существуют и другие формы ГР с меньшей молекулярной массой (20 000 дальтон), но они обладают низкой биологической активностью по сравнению с молекулой 22 К, и их физиологическая роль в процессе роста ребенка незначительна.
ГР синтезируется и секретируется соматотропными клетками передней доли гипофиза. Общее суточное количество ГР в гипофизе достигает
5–10 мг.
Синтезируемый в аденогипофизе ГР поступает в кровь в импульсном режиме в количестве 5–20 нг/мл. Продолжительность пикового выброса варьируется от 2 до 6 ч. Количество ГР, секретируемого у здоровых детей, зависит от возраста, пола и стадии полового созревания и достигает 0,02– 0,06 МЕ/кг/сут. Медленная фаза полувыведения ГР из плазмы у детей составляет 20 мин, быстрая фаза — 4 мин. С возрастом отмечается уменьшение секреторной активности соматотропных клеток гипофиза. Начиная с 20-летнего возраста, наблюдается снижение количества секреторных импульсов ГР, их амплитуды и продолжительности, что приводит каждые десять лет к уменьшению уровня циркулирующего в организме ГР на 14– 15 % и укорочению фазы его полувыведения на 6 %.
У взрослых выявлены достоверные половые различия секреции ГР. Более высокая суточная продукция ГР отмечена у женщин среднего возраста (0,6 МЕ/сут) по сравнению с мужчинами-сверстниками (0,2 МЕ/сут).
3

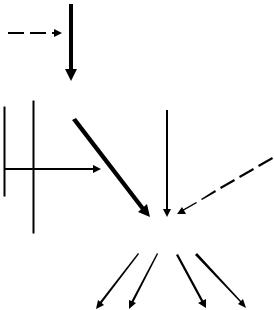
Синтез и секреция ГР контролируются гипоталамическими нейропеп-
тидами: ГР-РГ (соматолиберин) и соматостатином (рис. 1). ГР-РГ сти-
мулирует синтез и определяет амплитуду секреторного пика ГР. Активность его обусловлена взаимосвязью с рецепторами ГР-РГ, расположенными на соматотропных клетках гипофиза. Действие соматостатина проявляется во влиянии на частоту и время пульсаторной секреции и менее выражено — на синтез ГР. Взаимодействие этих двух гипоталамических гормонов играет важную роль в поддержании физиологической импульсной секреции ГР.
Рис. 1. Регуляция синтеза и секреции гормона роста
4
Предполагается, что помимо гипоталамических нейропептидов существует еще одна регуляторная система — ГР-рилизинг-пептиды (ГР-секре- тогоны). ГР-стимулирующая активность этих соединений и их небелковых аналогов значительно превышает таковую ГР-РГ. Возможный механизм действия ГР-рилизинг-пептидов связан с непосредственной стимуляцией соматотропных клеток гипофиза и с опосредованным действием на аркуатные ядра гипоталамуса с высвобождением ГР-РГ. Установлено, что ГР- рилизинг-пептиды вызывают деполяризацию соматотропных клеток, ускоряют внутриклеточное поступление кальция, действуя на уровне гипофиза как антагонисты соматостатина. Места связывания ГР-рилизинг-пептидов выявлены в гипоталамусе и гипофизе. В настоящее время в ткани гипофиза человека определен рецептор ГР-секретагонов, что является доказательством существования в организме неизвестного ГР-стимулирующего нейропептида, медиаторами действия которого являются ГР-рилизинг- пептиды и их небелковые аналоги.
Нейроэндокринная регуляция секреции ГР-РГ и соматостатина до настоящего времени полностью не раскрыта (табл. 1). Известно участие таких нейротрансмиттеров и нейропептидов, как серотонин, гистамин, дофамин, ацетилхолин, тиреотропин-рилизинг-гормон, гастрин, кальцитонин, нейропептид У, вазопрессин, кортикотропин-рилизинг-гормон, галанин. Эти факторы могут оказывать непосредственное влияние на секрецию ГР при различных физиологических состояниях (стресс, сон, кровотечение, голодание, гипогликемия, физическая нагрузка), лежащих в основе ГР-сти- муляционных тестов.
|
Таблица 1 |
Нейроэндокринная регуляция ГР (по M. B. Ranke, 1996) |
|
Стимуляторы секреции ГР |
Ингибиторы секреции ГР |
Агонисты α-рецептора |
Антагонисты α-рецептора |
Клонидин |
Фентоламин |
Аргинин |
|
Орнитин |
|
Вазопрессин |
|
Стресс |
|
Физическая нагрузка |
|
Гипогликемия |
|
Глюкагон |
|
Галанин |
|
Леводопа |
|
Агонисты β-рецептора |
Антагонисты β-рецептора |
Сальбутамол |
Пропроналол |
Допаминергические агонисты |
Допаминергические антагонисты |
Леводопа |
Пимазид |
Пиридостигмин |
|
Бромкриптин |
|
Серотонинергические агонисты |
Серотонинергические антагонисты |
Триптофан |
Метизергид |
5
Тиреоидные гормоны, глюкокортикоиды, половые гормоны также играют существенную роль в физиологической регуляции функции соматотропных клеток.
Известно, что гипотиреоз ведет к уменьшению как спонтанной, так и стимулированной секреции ГР, что связано с влиянием тиреоидных гормонов на процесс синтеза ГР-РГ и связывания с его соматотропными рецепторами. Кроме того, вероятно и непосредственное воздействие гормонов щитовидной железы на экспрессию гена ГР.
Половые гормоны являются активными модуляторами секреции ГР. Разное соотношение женских и мужских половых гормонов в организме определяет половой диморфизм выброса ГР во время пубертата. При мужском типе секреции отмечается только увеличение амплитуды пиков ГР при сохранении неизменными их частоты, продолжительности, а также периода полувыведения гормона. Женский тип выделения ГР отличается более частыми сниженными импульсами с наличием высокого межпикового уровня. Потенциальным механизмом влияния половых гормонов на секрецию ГР является воздействие на нейроны ГР-РГ и соматостатина в ядрах гипоталамуса. Установлено, что основным фактором, определяющим половое различие секреции ГР, служит концентрация эндогенного эстрадиола, выделяемого гонадами или образованного путем ароматизации из тестостерона. В ГР-РГ и соматостатинсодержащих нейронах найдены рецепторы эстрогенов.
Глюкокортикоидные гормоны при кратковременном влиянии или в физиологических концентрациях повышают выработку ГР путем активации экспрессии гена ГР и/или увеличения количества рецепторов к ГР-РГ. При длительном воздействии глюкокортикоидов отмечено уменьшение выброса ГР, обусловленное их стимулирующим эффектом на β2-адрено- рецепторы.
В последнее время в литературе активно обсуждается роль грелина (белка, обнаруженного в клетках желудка и гипоталамуса) как одного из ключевых компонентов в регуляции роста ребенка. В эксперименте на животных показано, что внутривенное введение грелина стимулирует аппетит, вызывает увеличение массы тела и ведет к ожирению, одновременно повышая плазменный уровень ГР.
Попадая в кровь, ГР взаимодействует с ГР-связывающим белком, и в связанном состоянии попадает в печень, где происходит активация выработки инсулиноподобных факторов роста 1 и 2 (ИФР-1 и ИФР-2) (рис. 2). Помимо печени ростовые факторы вырабатываются в надпочечниках, хрящевой ткани, легких, почках, половых железах. Аминокислотная последовательность этих пептидов на 50 % подобна таковой у инсулина. Большинство эффектов ИФР на клеточном уровне реализуются через рецептор к ИФР-1, представленный практически во всех органах и тканях.
6
Гипофиз
Хронический стресс
Инсулин Половые гормоны Кортизол Т4
 ГР Питание
ГР Питание
Иммунная система
Поражение печени Травмы
 ИФР-1
ИФР-1
Ткани мишени
Рис. 2. Регуляция действия ИФР-1
Биологическое действие факторов роста проявляется острыми анаболическими сдвигами в белковом обмене, позднее — в стимуляции пролиферации и дифференцировки клеток различных типов, в первую очередь, хрящевой и костной ткани. Острый эндокринный эффект ИФР проявляется, прежде всего, инсулиноподобным действием.
Оба ИФР регулируют синтез и секрецию гормонов клетками Лейдига, тиреоцитами, зернистыми и текальными клетками яичника, усиливают секреторную активность клеток коры надпочечников в ответ на стимуляцию АКТГ, выполняют роль медиатора ГР при синтезе таких важных субстратов, как ДНК, белки, гликозоаминогликаны и протеогликаны. Оба фактора роста участвуют в развитии плода, до 12 недель беременности значимую роль играет ИФР-2, во 2-м и 3-м триместрах — ИФР-1. В постэмбриональном периоде основное значение в регуляции роста имеет ИФР- 1. Его уровень у новорожденных составляет от 30 до 50 % уровня взрослых. В детстве сывороточная концентрация ИФР-1 медленно повышается, достигая максимальных значений в период пубертата (в 2–3 раза выше, чем у взрослых), и лучше коррелирует со степенью полового развития ребенка, чем с хронологическим возрастом.
В биологических средах 80 % ростовых факторов находятся в виде комплексов с ИФР-связывающими белками (ИФРСБ). К настоящему времени установлено 6 разновидностей ИФРСБ. Они имеют сходную структуру и специфически связываются только с ИФР, но не с инсулином. Наиболее распространен ИФРСБ типа 3. Найдены два комплекса ИФР-
7
ИФРСБ, имеющих молекулярную массу 40–50 и 150 кДа. В состав последнего входит еще один компонент — гликопротеин — кислотно-лабильная субединица, что значительно увеличивает стабильность соединения с удлинением периода полужизни комплексного ИФР до 12–15 ч (период полужизни не связанных ИФР менее 10 мин).
Физиологическая роль ИФР-связанных комплексов до конца не уточнена. Установлено, что трехкомпонентный комплекс циркулирует только в пределах сосудистого пространства, не проникая в лимфу или другие экстраваскулярные жидкости. Возможно, что исключительно внутрисосудистое нахождение ИФР играет важную роль в поддержании нормального уровня гликемии. В норме плазменные концентрации ИФР достоверно превышают уровень инсулина, но преимущественное содержание протеинов в составе трехкомпонентного комплекса препятствует проявлению их инсулиноподобного действия. Комплекс 40–50 кДа играет основную роль в реализации ростового эффекта ИФР. Он доставляет ИФР-1 к тканяммишеням, на поверхности которых находятся специфические рецепторы к ростовым факторам (табл. 2).
Таблица 2
Гормональная регуляция сывороточного уровня ИФР-1 и ИФРСБ-3
(по W. F. Blum, 1996)
Гормоны* |
Наблюдаемый эффект |
|
Гормон роста |
↑↑↑ |
|
Пролактин |
↑? |
|
Инсулин |
↑ |
|
Тиреоидные гормоны |
↑ |
|
Глюкокортикоиды |
↑ |
|
Андрогены |
↑ |
|
Эстрогены |
низкие дозы ↑ |
|
высокие дозы ↓ |
||
|
* Гонадотропные гормоны, тиреотропный гормон, АКТГ, паратгормон не оказывают прямого влияния на уровень ИФР-1 и ИФРСБ-3, но стимулируют продукцию ИФР-1 тканями-мишенями.
Также как и ГР, оба ИФР действуют на гипоталамус и аденогипофиз по принципу обратной связи, контролируя синтез соматолиберина и соматостатина и секрецию ГР.
8
ФИЗИОЛОГИЧЕСКАЯ РОЛЬ ГР
ГР обладает выраженным метаболическим влиянием на различные ткани и органы.
СТИМУЛЯЦИЯ РОСТА
Основной эффект ГР на детский организм проявляется в стимуляции продольного роста костей, преимущественно длинных трубчатых и в меньшей степени — губчатых костей (например, позвонков). В ходе этого процесса пролиферирующие хондроциты эпифизарной ростовой пластинки формируют хрящевую матрицу, которая в последующем подвергается дегенерации и замещается костной тканью по направлению к метафизу кости. Эпифизарная ростовая пластинка представляет собой хрящевую структуру между окостеневшим эпифизом и метафизом. Выделяют следующие зоны ростовой пластинки: зародышевая зона, зона пролиферации, гипертрофическая зона и зона кальцификации. Ширина пластинки пропорциональна интенсивности деления клеток и отражает скорость продольного роста кости. В процессе роста клетки зародышевой зоны, которая граничит с окостеневшим эпифизом, подвергаются дифференцировке и начинают распространяться в зону пролиферации. На конечной стадии клетки претерпевают дегенеративные изменения в гипертрофической зоне и зоне кальцификации и внедряются в метафиз кости. Минерализация начинается в области сосудов, которые расположены во внеклеточном матриксе между клетками гипертрофического слоя.
Рецепторы ГР определяются в разные возрастные периоды как в зародышевой, так и в пролиферативной и гипертрофической зонах ростовой пластинки. В настоящее время широко дискутируется гипотеза так называемого «двойного эффекта» ГР, в соответствии с которой ГР первоначально вызывает дифференцировку клеток-предшественников соединительной ткани, после чего ИФР-1 воздействует на рост клона этих клеток. В раннем возрасте основное влияние оказывает паракринный эффект образуемого в ростовой пластинке ИФР-1. В пубертатный период, когда отмечается значительное возрастание уровней ИФР-1, большое значение приобретает его эндокринное действие.
ВЛИЯНИЕ НА МЕТАБОЛИЗМ КОСТНОЙ ТКАНИ
ГР вместе с половыми гормонами и модуляторами фосфорно-каль- циевого обмена (паратгормон, витамин Д, кальцитонин) является одним из активных участников системы регуляции метаболизма костной ткани. ГР активирует обмен костной ткани, вызывая усиление остеобразования, и в меньшей степени — остеорезорбции. ГР играет важную роль в онтогенезе, стимулируя экспрессию протеогликанов зубного матрикса.
9
ВЛИЯНИЕ НА ЖИРОВОЙ ОБМЕН
ГР обладает выраженным липолитическим эффектом, который сохраняется даже при ограничении потребления калорий. Липолитическое действие проявляется, с одной стороны, в увеличении количества клетокпредшественников — преадипоцитов. С другой стороны, ГР вызывает ряд эффектов, направленных на уменьшение размеров зрелых жировых клеток, что приводит к общему снижению объема жировой ткани. Такими эффектами являются подавление активности липопротеинлипазы, гидролизирующей триглицериды до неэстерифицированных жирных кислот, которые повторно эстерифицируются и откладываются в адипоцитах. Также к числу липолитических эффектов относится и контринсулярное действие ГР.
ВЛИЯНИЕ НА УГЛЕВОДНЫЙ ОБМЕН
ГР оказывает двухфазное влияние на углеводный обмен. При введении физиологических доз ГР отмечается кратковременный инсулиноподобный эффект, который вызывает снижение уровня сахара в крови, затем за счет активации печеночной продукции глюкозы и уменьшения периферического клиренса углеводов отмечается контринсулярное действие. Длительное воздействие высоких доз ГР приводит к формированию выраженной инсулинорезистентности в сочетании с гиперинсулинемией и уменьшением синтеза гликогена в мышцах.
АНАБОЛИЧЕСКИЙ ЭФФЕКТ
Анаболический эффект ГР связан как с непосредственным влиянием собственно ГР на синтез белка, так и за счет ИФР-1. В мышечной ткани ГР действует на клетки-предшественники миобластов, вызывая их дифференцировку, и не влияет на клеточный объем. Эффект ИФР-1 на рост скелетных мышц изучен недостаточно, однако имеются данные о прямой зависимости содержания ИФР-1 и мРНК ИФР-1 от уровня ГР. В скелетных мышцах ГР действует на клетки-предшественники миобластов, вызывая их дифференцировку.
ПЕРИОДЫ РОСТА РЕБЕНКА
Условно выделяют следующие периоды роста ребенка: пренатальный и постнатальный, включающий младенчество, детство и пубертат.
ПРЕНАТАЛЬНЫЙ ПЕРИОД
Пренатальный период отличается активным уровнем процессов деления клеток и морфогенеза и характеризуется наиболее высокими темпами роста, которые не встречаются ни в одном другом периоде жизни. Выде-
ляют эмбриональную фазу роста (до 12 недель беременности) и феталь-
ную (после 12 недель беременности).
10