

03_Zaplidnennya_zv
.docxЗапліднення
-
Дистантні взаємодії гамет.
-
Контактні взаємодії гамет.
-
Акросомна та кортикальна реакція.
Основні поняття: запліднення, гіногамні сперміолізіни, андрогамні сперміолізіни, хемотаксис, дістантні взаємодії, акросомна реакція, кортикальна реакція, сінгамія, пронуклеус.
Запліднення – це процес злиття двох статевих клітин (гамет), внаслідок чого виникає нова особина, генетичні потенції якої беруть початок від обох батьків. Воно виконує дві функції: статеву – комбінування генів двох батьків, передача генів від батьків нащадкам, та репродуктивну – ініціацію в цитоплазмі яйця тих реакцій, які дозволяють продовжувати розвиток нового організму.
Запліднення супроводжується відновленням диплоїдної генетичної структури та активує яйцеклітину до подальшого розвитку. Диплоїдна клітина – зигота, яка утворилася в результаті запліднення, є початковим етапом розвитку нового організму. Під час запліднення сперматозоїд виконує дві функції:
-
активація яйця – спонукання яйця до розвитку – не специфічна функція, активуючими факторами можуть бути хімічні або механічні агенти (розвиток яйцеклітини без участі сперматозоїду називається партеногенезом);
-
внесення до яйцеклітини генетичного матеріалу батька – є специфічної функцією сперматозоїда.
Для будь-якої тварини, навіть при природному партеногенезі, запліднення є обов’язковим процесом в індивідуальному розвитку.
Процес запліднення складається з трьох послідовних фаз:
-
дистантні взаємодії (зближення гамет), здійснюються на деякій відстані, до зіткнення гамет;
-
контактні взаємодії (активації яйцеклітини та сперматозоїда), які відбуваються при безпосередньому зіткненні поверхонь гамет;
-
процеси, які протікають після входження сперматозоїда в яйце – сінгамия (злиття гамет: плазмогамія, каріогамія).
Взаємодії чоловічих і жіночих гамет під час запліднення називають дистантними, до них відносять:
-
хемотаксис – здатність сперматозоїдів рухатися по градієнту концентрації речовин, що виділяються яйцеклітиною, показаний для багатьох безхребетних (гідроїдні, молюски, глошкіряні, напівхордові);
-
реотаксис – здатність сперматозоїдів рухатися проти току рідини в статевих шляхах самиці;
-
стереотаксис – здатність сперматозоїдів рухатися у напрямку до більш крупного, ніж він сам, об’єкту, яким є яйце.
Другий тип взаємодії спермію та яйця – це активація спермію матеріалом драглистої оболонки, яка викликає акросомну реакцію. Акросомна реакція може бути визвана не тільки контактом сперматозоїда з оболонкою яйця, але й зіткненням з будь-якою твердою поверхнею, або підвищенням концентрації Ca2+. Вона зводиться до дуже швидких змін в акросомному апараті сперматозоїда, які призводять до вивільненя речовин (ферментів) акросомної гранули та викиданню акросомної нитки у бік твердого субстрата або поверхні яйця (рис. 1).
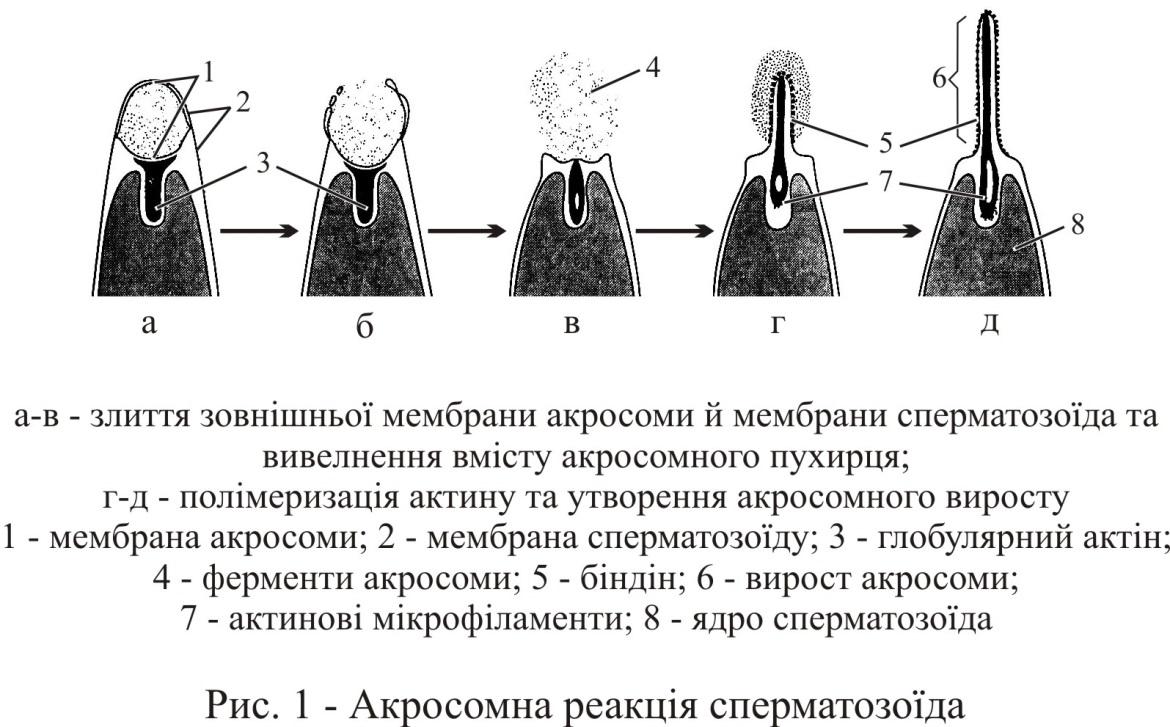
Починається акросомна реакція, коли апікальна частина головки сперматозоїда вступає в контакт з драглистою оболонкою яйцеклітини, зокрема її глікопротеінами, що призводить до відкриття Ca2+ та Na+ каналів. В результаті надходження іонів кальцію до сперматозоїду відбувається злиття плазмолеми з передньою частиною мембрани акросоми. Внаслідок цього акросома розкривається, протеази вивільнюються та потрапляють на третичну (у ссавців на драглисту zona pellucida) оболонку та розчиняють її.
В результаті надходження Na+ із зовнішнього середовищадо клітини, відбувається відток H+ (через Na+/H+-обмінник), пілвищується pH, що призводить до вибухоподібного переходу G-актина у F-актин. В результаті полімеризації актина задня частина мембрани акросоми утворює безліч паличкоподібних виростів – акросомних мікроворсинок (поліхети) або один довгий виріст – акросомну нитку (глошкірі). Підвищення pH також призводить до активації протеїнову АТФ-азу у шійки сперматозоїду, що призводить до підвищення рухової активності джгутика.
Акросомний виріст на своїй апікальній мембрані несе відоспецифічний білок біндін. На поверхні яйця розташовуються рецептори біндіна. «Упізнавання» яйцеклітиною свого виду полягає в з’єднанні рецепторів яйця з біндінами сперматозоїдів. Завдяки біндінам здійснюється цитологічний блок міжвидового схрещування.
У ссавців активація сперматозоїда відбувається без утворення акросомного виросту. Дисоціація мембран сперматозоїда та яйцеклітини відбувається уздовж голівки сперматозоїда. Ферменти акросоми розчиняють променистий вінець, після чого сперматозоїд вступає в контакт з блискучою оболонкою, яка утворена трьома типами білків: ZP3 і ZP2 знаходяться паралельно поверхні яйця, а білок ZP1 «зшиває» їх та розташован перпендикулярно поверхні яйця. Контакт з ZP3 – це перша частина акросомної реакції. Далі відбувається вилиття ферменту акросоми проакрозіну, який взаємодіє з ZP2 та лізує блискучу оболонку.
Закінчується реакція активації сперматозоїда злипанням задньої мембрани акросоми сперматозоїда та яйця, в результаті чого формується єдина зовнішня мембрана, що обмежує канал, через який ядро сперматозоїда та проксимальна центріоль проникають в цитоплазму яйця. Мембрана сперматозоїда, яка вбудовується в мембрану яйця, деякий час продовжує зберігати особливі властивості, зокрема має підвищену проникність для іонів Na+. В результаті мембранний потенціал яйцеклітини з негативного на деякий час стає слабо позитивним. Саме в цей час яйце стає непроникним для інших сперматозоїдів. Це явище отримало назву швидкого блоку поліспермії. Механізм швидкого блоку пояснюється наявністю в мембранах яйця та сперматозоїда рецепторів, чутливих до знака заряду.
В процесі активації яйцеклітини безпосередньо беруть участь мембрана яйця та кортекс. В кортексі зосереджені глобулярний білок (G-актин) та велика кількість мембранних пухирців – кортикальних гранул – структур, багато в чому гомологічних акросомі сперматозоїда.
Молекулярні механізми активації яйцеклітини пов’язані з включенням в дію інозитолфосфатної системи регуляції, основними компонентами якої є PIP2 (похідне вуглеводу інозиту фосфатіділінозитол-4,5-діфосфат) та фермент фосфоліпаза С. Знаходячись в неактивній формі, фосфолипаза С активується в результаті контакту між мембранами сперматозоїда й яйцеклітини та розщеплює PIP2 на діацилгліцерол (DAG), що залишається в мембрані, та інозитолтрифосфат (IP3), що виходить в цитоплазму. DAG за допомогою іншого мембранного ферменту, протеінкінази С, стимулює роботу мембранного механізму, що транспортує іони Na+ всередину клітини та протони – назовні (Na+/H+-обмінник). Зростання внутрішньоклітинного рН стимулює синтез білку в цитоплазмі яйцеклітини та синтез ДНК в чоловічому й жіночому ядрах.
У свою чергу IP3, що виділився в цитоплазму яйцеклітини, стимулює вивільнення іонів Са2+ з відсіків ендоплазматичного ретикулуму. В результаті в цитоплазмі виникає вузька хвиля підвищеної концентрації Са2+, яка рухається від точки входження сперматозоїда до протилежної точки поверхні яйцеклітини. Вузькість хвилі пояснюється тим, що безпосередньо після свого звільнення іони кальцію знову зв’язуються, інакше уся активність клітини буде паралізована.
Короткочасний підйом концентрації Са+ в цитоплазмі запускає цілий ряд процесів, пов'язаних з активацією яйцеклітини: синтез білку, реплікація ДНК, екзоцитоз кортикальных гранул. Сукупність цих прроцесів називаєьтся кортикальною реакцією, яка до тогож забезпечує розвиток повільного блоку поліспермії.
Внаслідок екзоцитозу кортикальних гранул у вузький простір між плазматичною мембраною яйцеклітини та жовтковою оболонкою, що щільно примикає до неї, вивільняються наступні речовини:
-
протеолітичний фермент вітелінова деламіназа, яка розриває зв’язки між плазматичною мембраною та жовточну оболонкою;
-
протеолітичний фермент спермрецепторна гідролаза, яка звільняє осілу на жовточно оболонку сперму від зв’язків з оболонкою;
-
осмотично активний глікопротеід, який вбирає воду в простір між жовточною оболонкою та плазматичною мембраною, викликаючи їх розшарування, внаслідок чого між ними виникає обширний простір – перівітеліновий простір;
-
фактор, що перетворює жовточну оболонку на непроникну для надмірних сперматозоїдів – оболонку запліднення;
-
структурний білок гіалін, який утворює шар над плазматичною цитомембраною, в подальшому розвитку він сприяє підтримці правильного взаємного розташування бластомерів в процесі дроблення.
У ссавців механізм екзоцитозу такий же, як у інших тварин. Проте блок поліспермії у різних видів створюється по-різному. В більшості випадків процеси запліднення відрегульовані так, що ймовірність проникнення в яйце більше одного сперматозоїда була зведена до мінімуму. У деяких груп тварин (акулових риб, рептилій, птахів) проникнення декількох сперматозоїдів не приводить до небажаних наслідків: спостерігається природна або фізіологічна поліспермія. Як правило, в їх яйцеклітину проникає до декількох десятків сперматозоїдів, але з ядром яйцеклітини взаємодіє тільки один. Інші або деякий час діляться з участю своїх центріолей, рано чи пізно гинучи, або розсмоктуються, не починаючи ділення. Передбачається, що у птахів ці надмірні спермії несуть трофічну функцію до утворення відповідної спеціалізованої тканини зародка. У інших (морський їжак, ссавці) оболонка запліднення стає непроникною так швидко, що в яйцеклітину встигає проникнути лише один сперматозоїд. Поліспермія у таких тварин рідкість, а якщо вона виявляється, то порушує нормальний розвиток.
Синтез білків, що починається вже в перші хвилини після активації яйцеклітини, йде виключно на матрицях (молекулах мРНК, що знаходяться до активації яйцеклітини у зарепресированному стані у складі інформосом) та рибосомах, які були накопичені в період оогенезу. Присутність ядра яйцеклітини або сперматозоїда для активації синтезу білку не потрібна. Активація здійснюється, таким чином, на рівні трансляції та вимагає, з одного боку, дерепресії інформосом, а з іншого – асоціації окремих рибосом в полірибосоми. Ці процеси активуються підвищенням pН та Са2+.
У більшості тварин сперматозоїд входить в яйце цілком, включаючи хвостову частину, у деяких джгутик залишається на поверхні. Але і в тих випадках, коли джгутик сперматозоїда заходить всередину яйця, він відділяється й розсмоктується. В подальших переміщеннях компонентів спермія усередині яйця бере участь центріоль. В період завершення жіночого мейозу компактизоване ядро головки сперматозоїда перетвориться в своєрідне ядро, зване чоловічим пронуклеусом. При цьому воно поступове збільшується в об’ємі, набухає, хроматин розпушується та набуває тонкогранулярної будови. Починається занурення чоловічого пронуклеусу, при цьому центріоль займає положення попереду ядра у напрямі занурення, а навколо виникає характерне «полярне сяйво». Судячи з усього, центріоль перетворюється на орган руху чоловічого пронуклеусу всередині яйця.
Схожі зміни – набухання та розпушування хроматину – відбуваються в гаплоідному ядрі яйцеклітини, яке формується після мейозу – в жіночому пронуклеусі. Перш ніж зближуватися, пронуклеуси виконують складні рухи. Спочатку чоловічий пронуклеус рухається всередину яйця перпендикулярно до поверхні та незалежно від положення жіночого пронуклеусу. Цей відрізок шляху називається «шляхом проникнення». В процесі руху прснуклеусів та розпушування хроматина в кожному з них репліцируется ДНК. Кінцева стадія зближення пронуклеусів – утворення метафазної пластинки ділення зиготи. Після того, як це перше ділення зиготи здійснюється по мітотичному типу, в ядрах перших двох клітин зародка, що утворилися, об’єднуються чоловічі та жіночі хромосоми. Кожне таке ядро диплоідно, тобто містить 2n хромосом та 2С ДНК.
Безпосередньо після проникнення сперматозоїда (або дії партеногенетичного агента) в яйцях різних груп тварин починаються інтенсивні переміщення складових частин цитоплазми (ооплазми). Іноді при цьому спостерігається розшарування, сегрегація складових частин – ооплазматична сегрегація. Ці процеси створюють певну просторову організацію майбутнього зародка. Ооплазматична сегрегація у різних видів протікає неоднаково: у деяких вона завершується до початку дроблення (в дозріваючому яйці й зиготі), у інших продовжується і в період дроблення.
Контрольні питання:
-
Які фази можна виділити в процесі запліднення?
-
Які фактори забезпечують зближення сперматозоїда з яйцем?
-
В чому полягають особливості акросомної реакції?
-
В чому полягають особливості кортикальної реакція?
-
Які речовини приймають участь у заплідненні?
-
Що таке сінгамія, каріогамія?
-
Ооплазматична сегрегація.
-
Як вирішується проблема поліспермії?
-
Повільний та швидкий блоки поліспермії.