

06_Neirulyaciya_zv
.docxНейруляція. провізорні органи хребетних
-
Загальна характеристика нейруляції, її біологічний смисл.
-
Формування нейтральної пластинки.
-
Утворення нервового жолобка та нейтральної трубки.
-
Перетворення мезодерми під час нейруляції.
-
Провізорні органи.
Основні поняття: нейруляція, нейрула, нервова пластинка, нервовий вал, нервова (медулярна) трубка, нервовий гребінь, невроцель, нейропор, склеротоми, міотоми, провізорні органи, амніон, хоріон, жовточний мішок, алантоіс, імплантація, плацента.
Після гаструляції у всіх метамерно побудованих тварин починається процес виділення й сегментації осьової мезодерми. У хордових на цьому етапі розвитку закладається та утворюється центральна нервова система (ЦНС). Через наочність її перетворень і важливості самої ЦНС весь період отримав назву нейруляція (рис. 1), а зародок – нейрули. Передній розширений відділ нервової трубки згодом стане головним, а задній – спинним мозком особини. Крім скручування нейральної пластинки в цей період відбуваються інші комплексні перетворення: оформлюється хорда, виділяється мезодерма майбутніх сомітів, починається сомітогенез. В цей період розвитку і у хордових, і у сегментованих безхребетних спостерігається багато загальних найважливіших подій: формується комплекс осьових структур, відбувається початкове відособлення та остаточне розташування в організмі закладок всієї решти органів – похідних экто-, енто- та мезодерми. Таким чином, на цьому етапі розвитку у тварин реалізується план будови організму, який має загальні риси. Вказані загальні гомологічні події в розвитку членистих безхребетних і всіх хордових дозволяють використовувати термін «нейруляція» як синонім поняття «закладка осьових структур і реалізація плану будови» та називати так цей період розвитку, пам’ятаючи, що у безхребетних немає центральної нервової системи, структурно гомологічної хордовим тваринам.
Формування нейральної пластинки. Протягом всієї нейруляції продовжуються вентро-дорсальні рухи клітин в екто- й мезодермальному листках. Конвергентна інтеркаляція викликає розтягування дорсальної сторони зародка в передньо-задньому напрямі, причому тулубний відділ витягується значно сильніше, ніж головний.
Клітки підповерхневого шару ектодерми (гіпоектодерма) зміщуються на спину зародка та утворюють стовбчастий епітелій. В результаті цього спинна сторона зародка сплощується та підводиться, утворюється нервова пластинка. Межі нервової пластинки встановлюються в результаті індукційних взаємодій клітин. Індукційні взаємодії, при яких осьова мезодерма впливає на презумптивну нейроектодерму, та спонукає її формувати нейральні структури, називають нейральною, або первинною, ембріональною індукцією. В результаті поверхня зародка розділяється на три зони – нейроектодерму, покривну ектодерму та прикордонну область клітин нервового гребеня.
Утворення нервового жолобка та нервової трубки. По краях нервової пластинки утворюється подковоподібний нервовий вал. Нервова пластинка усередині валу починає швидко звужуватися та згинатися в жолоб. Цьому сприяють як конвергентні рухи клітин і подовжнє розтягування, так і зміни форми клітин. Уздовж серединної лінії пластинки утворюються колбоподібні клітини та подовжній жолобок (рис. 2). Клітини обох шарів нервової пластинки витягуються перпендикулярно пласту, їх апікальні поверхні скорочуються, при цьому клітини поверхневого шару занурюються вглиб і перемішуються з клітинами шару гіпоектодерми. Ці центральні клітини міцно були пов’язані з закладкою хорди, яка лежить нижче. Краї ж нервової пластинки підіймаються вгору під тиском сомітної мезодерми, яка лежить нижче.
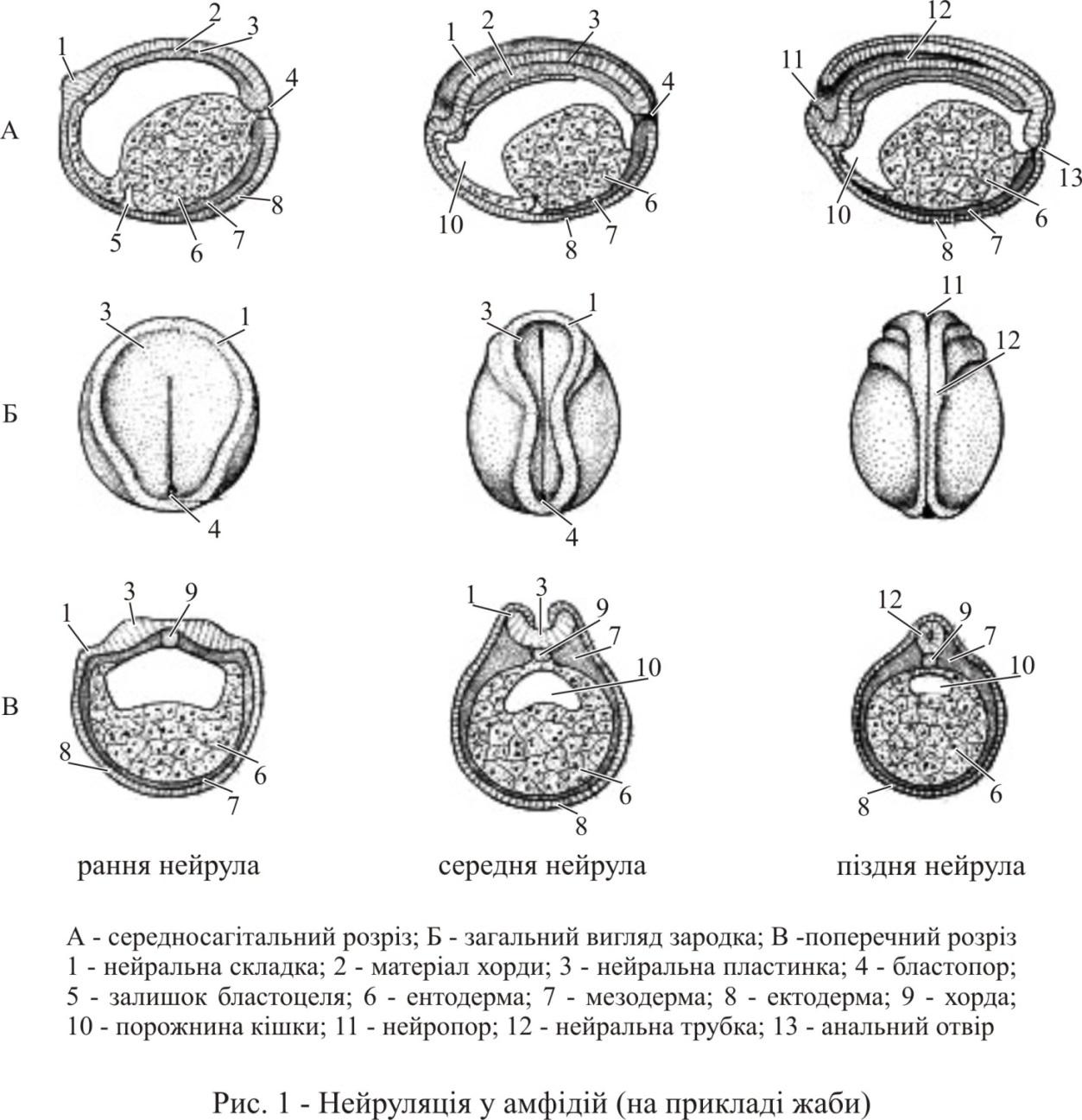

Нервові вали зближуються і приходять в контакт. Нейруляція закінчується, коли нервовий жолоб замикається в нервову (медулярну) трубку, покривна ектодерма зливається над нею в суцільний двошаровий епітелій, а з матеріалу нервових валів утворюється окремий подовжній тяж клітин – нервовий гребінь, або гангліозна пластинка (рис. 2). Порожнина всередині нервової трубки називається невроцелем. Передня ділянка презумптивного головного мозку досить довго не закривається. Отвір на передньому кінці нервової трубки називається нейропором (або невропором). Задній відділ нервової пластинки при замиканні в трубку прикриває своїм зводом бластопор. Він знаходиться тепер на задньому кінці медулярної трубки та стає нервово-кишковим каналом (у деяких хребетних, зокрема у хвостатих амфібій, нервово-кишковий канал не виникає). В області шиї та тулуба нервова трубка диференціюється в спинний мозок, а передня частина нервової трубки перетворюється на головний мозок. Передня ділянка нервової пластинки у всіх хребетних, особливо у людини, значно ширша, з самого початку формування нервової трубки діаметр її неоднаковий в різних ділянках. Значно розширена передня, головна ділянка, стінки трубки тут товщі, ніж в задній частині. Дорсальна та вентральна ділянки стінки медулярної трубки більш тонкі, ніж бічні ділянки, що розростаються. Канал нервової трубки перетворюється на центральний спинномозковий канал розвинутого мозку.
В різних ділянках переднього відділу нервової трубки завдяки нерівномірному зростанню дуже рано стають помітними в певних ділянках потовщення, утворюються карманоподібні випинання, значні в ділянках майбутніх мозкових півкуль. Одночасно з тієї ж причини зовні рано утворюються в певних місцях неглибокі стиснення нервової трубки та виразно позначаються три мозкові міхури: передній (prosencephalon), середній (mesencephalon) і задній (rombencephalon). Передній мозковий міхур надалі підрозділяється на зачаток великого мозку (telencephalon) і зачаток проміжного мозку (diencephalon). Mesencephalon, розвиваючись і диференціюючись, перетворюється на середній мозок. Rombencephalon при подальшому розвитку підрозділяється на зачаток мозочка й моста (metencephalon) та зачаток довгастого мозку (myelencephalon).
Довгастий мозок переходить в спинній. Верхня стінка myelencephalon залишається при подальшому розвитку мозку тонкою, і в ній не утворюються нервові клітини. Велика частина верхньої стінки згодом вростає разом з судинною оболонкою в порожнину четвертого шлуночка мозку, утворюючи судинне сплетення четвертого шлуночка (plexus chorioideus). Довгастий мозок утворюється в результаті розростання й розвитку бічних і нижніх стінок myelencephalon. Оскільки при розвитку довгастого мозку у всіх хребетних на його поверхні утворюються жолобки (борозенки), то він розділяється на велике число сегментів – нейромерів. Особливо виразна сегментація у зародків риб. Однак вона не пов’язана з метамерним розвитком черепних нервів, а носить тимчасовий характер, і від неї не зберігається сліду в дефінітивному мозку.
Ще до виникнення виразного
підрозділу переднього мозкового міхура
на зачаток великого і проміжного мозку
на його бічних стінках випинаються очні
міхури (рис. 3).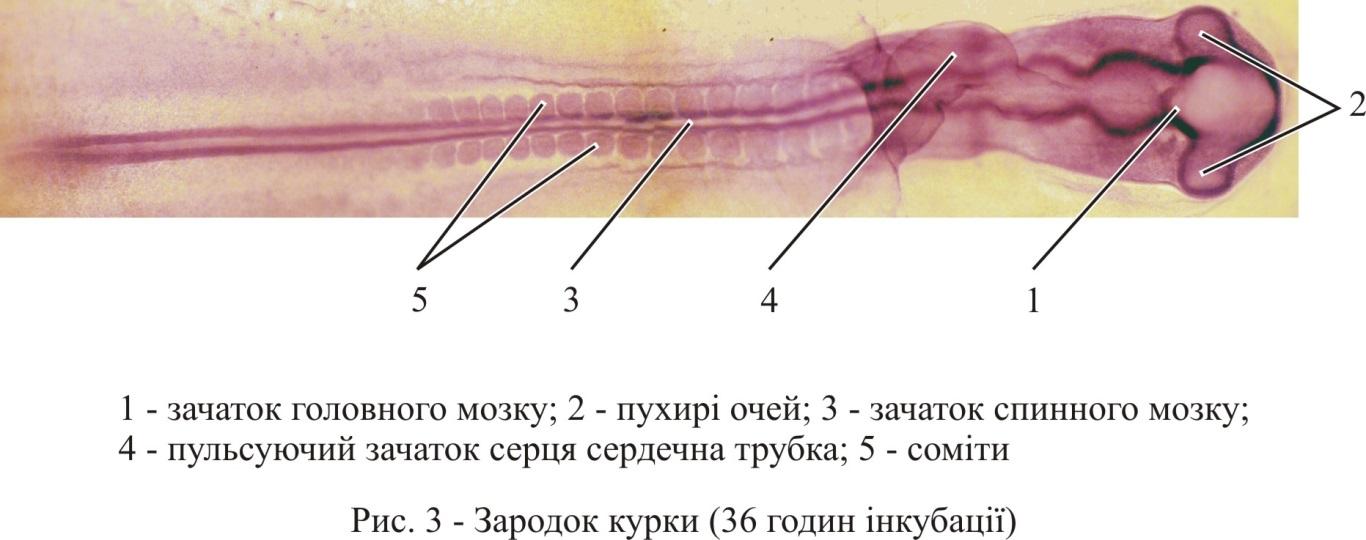
Дорсальна частина нервової трубки витягується сильніше, ніж вентральна, внаслідок чого задній кінець трубки виноситься за бластопор. Потім відбувається згинання трубки на деякій відстані від бластопору. Вершина вигину, яки утворився, стає зачатком хвоста. Ділянка, що лежить між вершиною зачатка хвоста та бластопором, диференціюється як м’язи хвостової області.
У різних тварин, залежно від ступеня складності нервової системи, є значні анатомічні й гістологічні особливості, а також особливості в топографії різних ділянок центральної нервової системи.
У нижчих хребетних, наприклад у акулових риб, форма дефінітивного мозку мало відрізняється від мозку їх зародків. У вищих хребетних, особливо у людини, мозок ранніх зародків за своєю морфологією мало відрізняється від мозку дорослих риб або амфібій, але дефінітивний мозок, особливо його клітинна й тканинна структури, різко відрізняється від мозку нижчих представників хребетних. Прогресивна еволюція організмів не обов’язково супроводжувалася ускладненням будови всіх органів. Так, в деякому відношенні очі у птахів або деяких комах досконаліші, ніж очі ссавців.
Вже в процесі замикання нервової трубки клітини нервового гребеня набувають рухливість і мігрують дорсовентрально. Ці клітини демонструють широкий спектр диференціювань. В головному відділі нащадки клітин нервового гребеня виявляються у складі кісток, хрящів і гладкої мускулатури лицьової частини, кісточок середнього вуха, одонтобластів зубних зачатків, сполучної тканини лицьової частини, щитовидної й слинних залоз, а також диференціюються в периферичні нейрони та глію. В тулубному відділі клітини нервового гребеня утворюють пігментні клітини шкіри – меланоцити, а також нейрони й глію периферичної нервової системи, включаючи дорсальні сенсорні ганглії та ганглії симпатичної й парасимпатичної систем.
Перетворення мезодерми в ході нейруляції. Мезодерма, яка займає місце з боків від хорди і розповсюджується далі між шкіряною ектодермою й ентодермою вторинної кишки, підрозділяється на дорсальну та вентральну області. Дорсальна частина сегментована та представлена парними сомітами (рис. 4). Закладка сомітів йде від головного до хвостового кінця. Вентральна частина мезодерми, що має вид тонкого шару клітин, називається бічною пластинкою. Соміти сполучені з бічною пластинкою проміжною мезодермою у вигляді сегментованих ніжок сомітів.
Усі області мезодерми поступово диференціюються. На початку формування соміти мають конфігурацію, характерну для епітелію з порожниною всередині. Під індукційною дією, яке іде від хорди та нервової трубки, вентромедіальні частини сомітів – склеротоми – перетворюються на вторинну мезенхіму, виселяються з соміту та оточують хорду й вентральну частину нервової трубки. Надалі з них утворюються хребці, ребра та лопатки.
Дорсолатеральна частина сомітів з внутрішньої сторони утворює міотоми, з яких надалі розвиваються поперечносмугасті скелетні м’язи тіла й кінцівок. Зовнішня дорсолатеральна частина сомітів утворює дерматоми, які дають початок внутрішньому шару шкіри – дермі. З області ніжок сомітів із зачатками нефротом і гонотом утворюються органи виділення й статеві залози.
Права та ліва несегментовані бічні пластинки розщеплюються на два листки, які обмежують вторинну порожнину тіла – цілом. Внутрішній листок, прилеглий до ентодерми, називають вісцелярним. Він оточує кишку з усіх боків і утворює брижу, покриває легеневу паренхіму та м’яз серця. Зовнішній листок бічної пластинки прилеглий до ектодерми називається парієнтальним. Надалі він утворює зовнішні листки очеревини, плеври та перикарду.
Порожнина бластоцеля в ентодермі на вентральній стороні зародка редукується.
Таким чином, в процесі нейруляції виникає комплекс осьових органів нервова трубка – хорда – кишка, що є характерною рисою організації тіла всіх хордових. Однакове походження, розвиток і взаємне розташування осьових органів виявляють їх повну гомологію та еволюційну спадкоємність.
При поглибленому розгляді
та порівнянні процесів нейруляції у
конкретних представників типу хордових
виявляються деякі відмінності, які
пов’язані в основному з особливостями
будови яйцеклітин, способу дроблення
та гаструляції. Звертають увагу відмінна
форма зародків і зсув часу закладки
осьових органів один до одного, тобто
гетерохронія.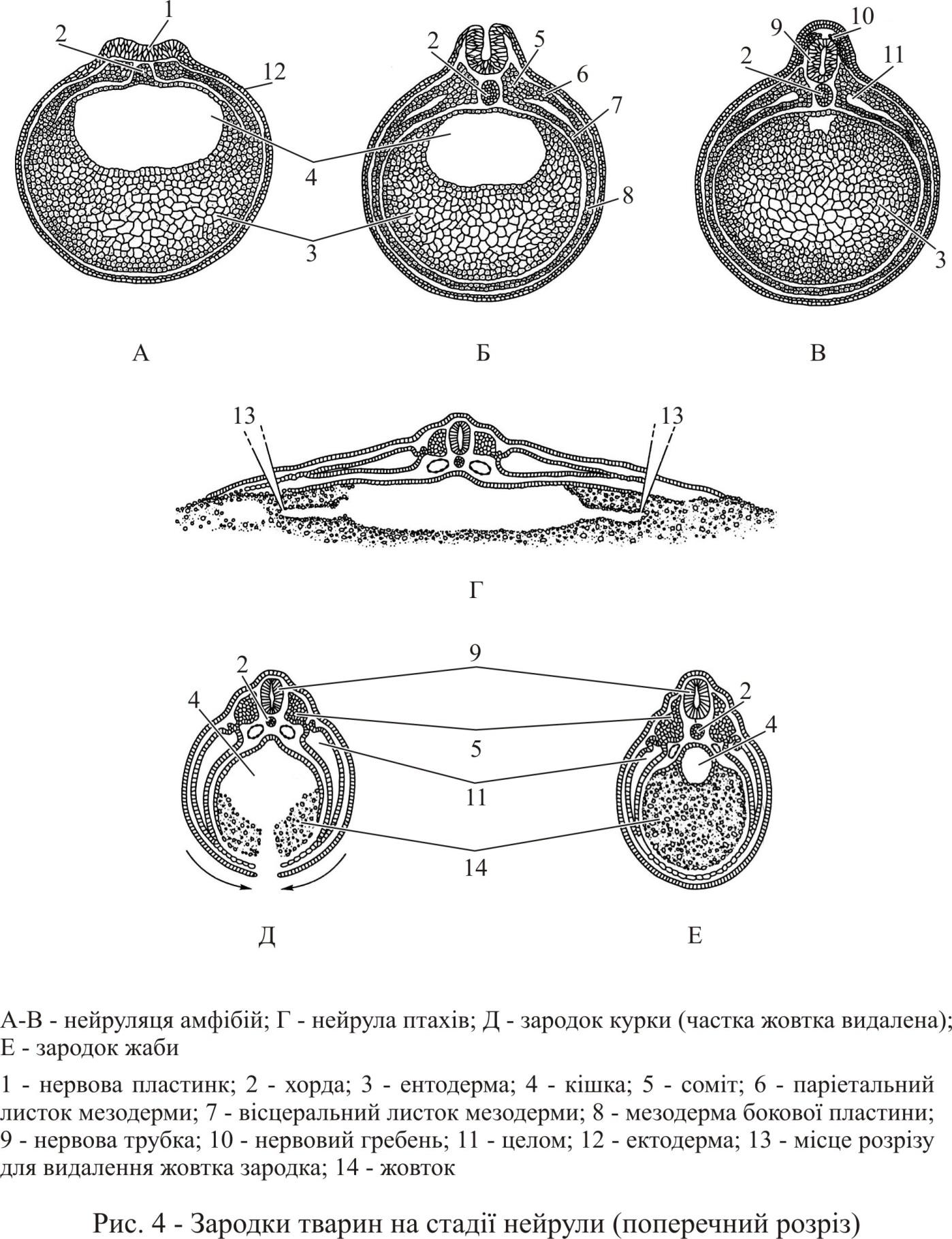
Провізорні, або тимчасові, органи утворюються в ембріогенезі ряду представників хребетних для забезпечення життєво важливих функцій, таких, як дихання, живлення, виділення, рух тощо. Недорозвинені органи самого зародка ще не здатні функціонувати за призначенням, хоча обов’язково відіграють певну роль в системі цілісного організму, що розвивається. Як тільки зародок досягає необхідного ступеня зрілості, коли більшість органів здатна виконувати життєво важливі функції, тимчасові органи розсмоктуються або відкидаються.
Час утворення провізорних органів залежить від кількості поживних речовин, які були накопичені в яйцеклітині та в яких умовах середовища відбувається розвиток зародка. Наприклад, у безхвостих земноводних в яйцеклітинах накопичена достатня кількість жовтка, розвиток відбувається у воді, зародок здійснює газообмін і виділяє продукти дисиміляції безпосередньо через оболонки яйця, досягає стадії пуголовка, на якій утворюються провізорні органи дихання (зябра), травлення та руху, пристосовані до водного способу життя. Перераховані личинкові органи дають можливість пуголовку продовжити розвиток. По досягненні стану морфофункціональної зрілості органів дорослого типу тимчасові органи зникають в процесі метаморфозу.
У плазунів і птахів запасів
жовтка в яйцеклітині більше, але розвиток
йде на суші. У зв’язку з цим дуже рано
виникає потреба в забезпеченні дихання
та виділення, а також в захисті від
висихання. У них вже в ранньому
ембріогенезі, майже паралельно з
нейруляцією, починається формування
провізорних органів, таких, як амніон,
хоріон і
жовточний мішок
(рис. 5). Трохи пізніше формується алантоіс.
У плацентарних ссавців
ці ж провізорні органи утворюються ще
раніше, оскільки в яйцеклітині дуже
мало жовтка. Розвиток таких тваринних
відбувається внутрішньоутробний,
утворення провізорних органів у них
співпадає за часом з періодом гаструляції.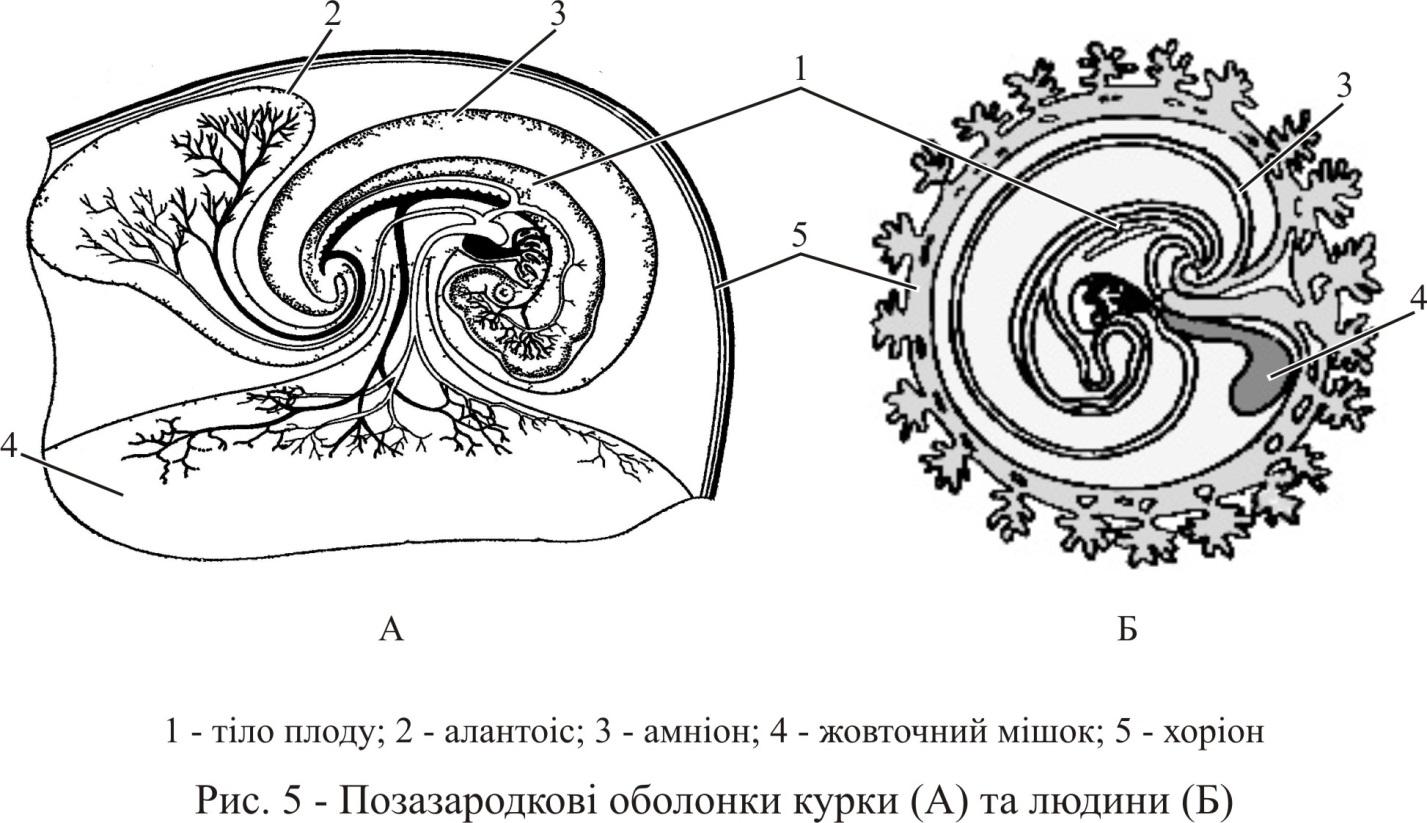
Наявність або відсутність амніона та інших провізорних органів лежить в основі розподілу хребетних на дві групи: Anamnia й Amniota. Еволюційно більш стародавні хребетні (круглороті, риби та амфібії), які розвиваються виключно у водному середовищі, не потребують додаткових водних та інших оболонок зародка й складають групу анамній. До групи амніот відносять первинно наземні хребетні (плазуни, птахи та ссавці), тобто ті, у кого ембріональний розвиток протікає в наземних умовах. Вони є вищими хребетними, оскільки мають скоординовані та високоефективні системи органів, що забезпечують їм існування в найскладніших умовах, якими є умови суші. Ці класи налічують велику кількість видів, які повторно перейшли у водне середовище. Таким чином, вищі хребетні змогли освоїти всі середовища. Подібна досконалість була б неможливою, у тому числі без внутрішнього запліднення та спеціальних провізорних ембріональних органів.
В будові й функціях провізорних органів різних амніот багато спільного. Характеризуючи в найзагальнішому вигляді провізорні органи зародків вищих хребетних, які ще називають зародковими оболонками, слід зазначити, що всі вони розвиваються з клітинного матеріалу зародкових листків, що вже сформувалися. Деякі особливості є в розвитку зародкових оболонок плацентарних ссавців.
Амніон – ектодермальний мішком навколо зародка заповнений амніотичною рідиною. Амніотична оболонка спеціалізована для секреції та поглинання амніотичної рідини, яка омиває зародок. Амніон відіграє певну роль в захисті зародка від висихання та від механічних пошкоджень, створюючи для нього найсприятливіше і природне водне середовище. Амніон має й мезодермальний шар з внезародковою соматоплеврою, який дає початок гладким м’язовим волокнам. Скорочення цих м’язів викликають пульсацію амніона, а повільні коливальні рухи, які передаються при цьому зародку, сприяють тому, що його зростаючи частини не заважають один одному.
Хоріон (сероза) – зовнішня зародкова оболонка, прилегла до шкаралупи або материнських тканин, виникаюча, як й амніон, з ектодерми та соматоплеври. Хоріон служить для обміну між зародком і навколишнім середовищем. У яйцеродних видів основна його функція – дихальний газообмін; у ссавців він виконує більш обширні функції, беручи участь окрім дихання в живленні, виділенні, фільтрації та синтезі речовин, наприклад гормонів.
Жовточний мішок має ентодермальне походження, покритий вісцеральною мезодермою та безпосередньо пов’язаний з кишковою трубкою зародка. У зародків з великою кількістю жовтка він бере участь у живленні. У птахів, наприклад в спланхноплеврі жовточного мішка, розвивається судинна сітка. Жовток не проходить через жовточну протоку, яка з’єднує мішок з кишкою. Спочатку він переводиться в розчинну форму під дією травних ферментів, які продукуються ентодермальними клітинами стінки мішка. Потім потрапляє в судини та з кров’ю розноситься по всьому тілу зародка.
У ссавців немає запасів жовтка, тому збереження жовточного мішка може бути пов’язано з важливими вторинними функціями. Його ентодерма служить місцем утворення первинних статевих клітин, мезодерма дає формені елементи крові зародка. Крім того, жовточний мішок ссавців заповнений рідиною з високою концентрацією амінокислот і глюкози, що вказує на можливість обміну білків в ньому.
Доля жовточного мішка у різних тварин дещо різна. У птахів до кінця періоду інкубації його залишки вже знаходяться всередині зародка, після чого він швидко зникає і до кінця 6-ї доби після вилуплення повністю розсмоктується. У ссавців жовточний мішок розвинений по-різному. У хижаків він порівняно великий, з сильно розвиненою сіткою судин, а у приматів швидко зморщується та зникає без залишку до пологів.
Алантоїс розвивається дещо пізніше за інші позазародкові органи. Він є мішкоподібним виростом вентральної стінки задньої кишки. Отже, він утворений ентодермой зсередини і спланхноплеврой зовні. У рептилій і птахів алантоїс швидко доростає до хоріону і виконує декілька функцій. Перш за все, це вмістище для сечовини і сечової кислоти, які є кінцевими продуктами обміну азотовмісних органічних речовин. В алантоїсі добре розвинена судинна сіть, завдяки чому разом з хоріоном він бере участь в газообміні. При вилупленні зовнішня частина алантоїса відкидається, а внутрішня – зберігається у вигляді сечового міхура.
У багатьох ссавців алантоїс теж добре розвинений, і разом з хоріоном утворює хоріоалантоїсну плаценту. Термін плацента означає тісне накладення або злиття зародкових оболонок з тканинами батьківського організму. У приматів і деяких інших ссавців ентодермальна частина алантоїса рудіментарна, а мезодермальні клітини утворюють щільний тяж, що простягається від клоачного відділу до хоріону. По мезодермі алантоїсу до хоріону ростуть судини, за допомогою яких плацента виконує функції виділення, дихання й живлення.
Для вищих ссавців характерне більш-менш щільне прикріплення зародка до стінок матки, що наступає через декілька днів після початку розвитку (у миші на 6-у, у людини на 7-у добу), коли зародок знаходиться на стадії бластоцисти. В основі процесу прикріплення, званого імплантацією, лежить занурення вторинних ворсинок хоріону в стінку матки. В результаті утворюється особливий орган – плацента, який має зародкову частину (ворсинки хоріону) і материнську частину (більш менш змінені стінки матки). До зародкової частини плаценти можна віднести також алантоїдну ніжку, яка має особливе значення для кровопостачання у нижчих ссавців (сумчасті), у яких материнська частина плаценти нерозвинена.
У вищих ссавців по глибині занурення ворсинок хоріону зародка й ступеня їх проникнення в слизову оболонку матки (гістологічна класифікація) розрізняють наступні типи плацент (рис. 6).
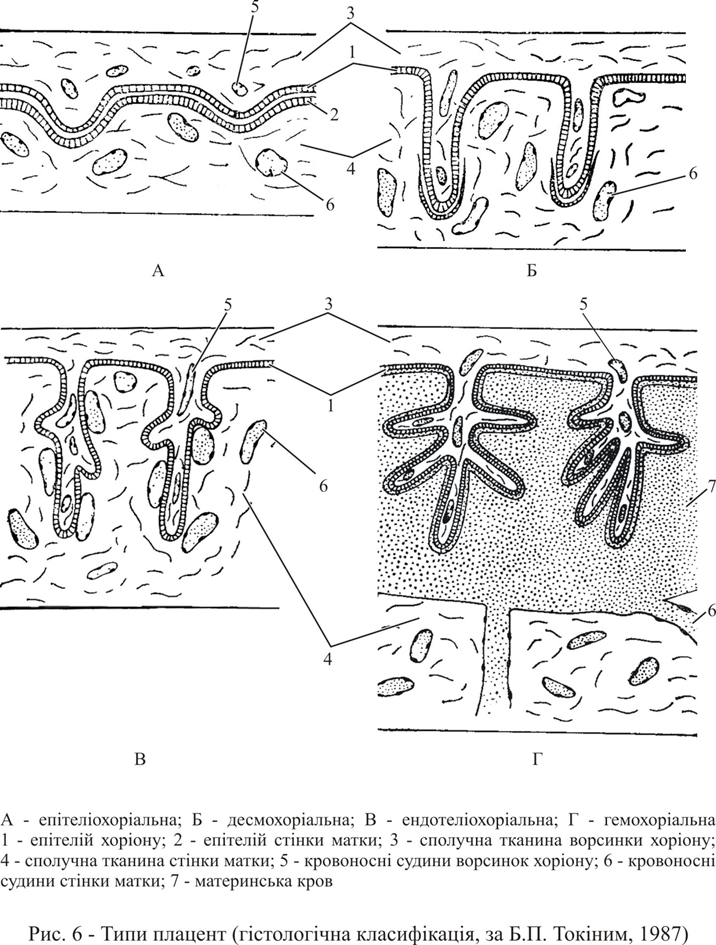
Епітеліохоріальна плацента (напівплацента) зустрічається у деяких копитних, лемурів і китоподібних. Ворсинки хоріону не проривають навіть епітеліального вистилання матки, а лише занурюються в складки її слизової оболонки. При пологах ворсинки хоріону відокремлюються від епітелію матки, не руйнуючи її.
Десмохоріальна плацента жуйних влаштована так, що ворсинки хоріону в місці контакту руйнують слизову оболонку матки та занурюються в її сполучний шар, але не досягають стінок кровоносних судин матки. Область контакту зародка з тканинами матері здійснюється в районах котиледон, невеликих майданчиків, розкиданих по поверхні обширного плодового міхура, що перевершує в сотні раз об’їм самого зародка. Після пологів в стінці матки залишаються ділянки, позбавлені епітелію. Подальша регенерація відбувається дуже швидко.
При ендотеліохоріальній (вазохоріальній) плаценті хижих між судинами плоду та матері встановлюється контакт, ворсинки хоріону проникають через увесь сполучнотканинний шар слизової оболонки матки та відділяються від її судин тільки їх ендотеліальною стінкою. При імплантації відбувається не тільки руйнування слизового епітелію матки, але й деструктивні процеси в сполучній тканині: трофобласт ворсинок хоріона опиняється в безпосередньому контакті з ендотелієм судин.
Найтісніший зв’язок судин плоду та матері здійснюється в гемохоріальній плаценті приматів і деяких інших рядів (рукокрилі, комахоїдні). Тут ворсинки хоріону також проривають ендотелій кровоносних судин слизового шару матки та безпосередньо стикаються з материнською кров’ю. Таким чином, кров матері й плоду розділена між собою лише тонкою зовнішньою оболонкою ворсинок хоріону та стінками капілярних судин зародка. Встановлено, що клітини ворсинок хоріону активно заковтують шляхом піноцитозу цілі крапельки крові матері. Будова гемохоріального типу плацент варіює у різних тварин. У мавп і людини ворсинки хоріону складно розгалужуються, і тим самим створюється величезна поверхня зіткнення ворсинок з кров’ю. Тому така плацента називається ворсинково-гемохоріальною плацентою. Збільшення поверхні контакту трофобласту з кров’ю в інших випадках (наприклад, у гризунів) досягається шляхом сильного розгалуження випинань трофобласту, які створюють складний лабіринт каналів, по яких тече материнська кров; тому такого типу плацента називається лабіринтово-гемохоріальною плацентою.
Природно, що при такому зв’язку зародка з матір’ю пологи супроводжуються сильною кровотечею, відпадає епітелій слизистої оболонки матки. Стінка матки має хорошу здатністю до регенерації, тому відбувається її швидке відновлення.
Крім вищенаведеної гістологічної класифікації плацент існує їх анатомічна (морфологічна) класифікація, заснована на розташуванні ворсинок по поверхні хоріону (рис. 7). Згідно цієї класифікації розрізняють:
-
дифузну плаценту – ворсинки утворюються по всій поверхні хоріону (свиня);
-
часткову (котиледонну, множинну, острівну) плаценту – ворсинки хоріону зібрані в групи (котиледони), яким відповідають певним чином трансформовані ділянки слизової оболонки матки (плацентоми), між котиледонами поверхня хоріону гладка (жуйні);
-
зонарну (пояскову) плаценту – ворсинки хоріону зосереджені всередині невеликої ділянки, яка опоясує плодовий міхур (хижі);
-
дискоїдальну плаценту – ворсинки сконцентровані в одній ділянці, яка має форму диска (у людиноподібних мавп і людини, у мавп і макак – бідискоїдальна).
Анатомічні типи плацент приблизно відповідають гістологічним типам. Так, зонарні плаценти є в той же час десмохоріальними, дискоїдальні – гемохоріальними тощо.
До основних функцій плаценти
відносяться: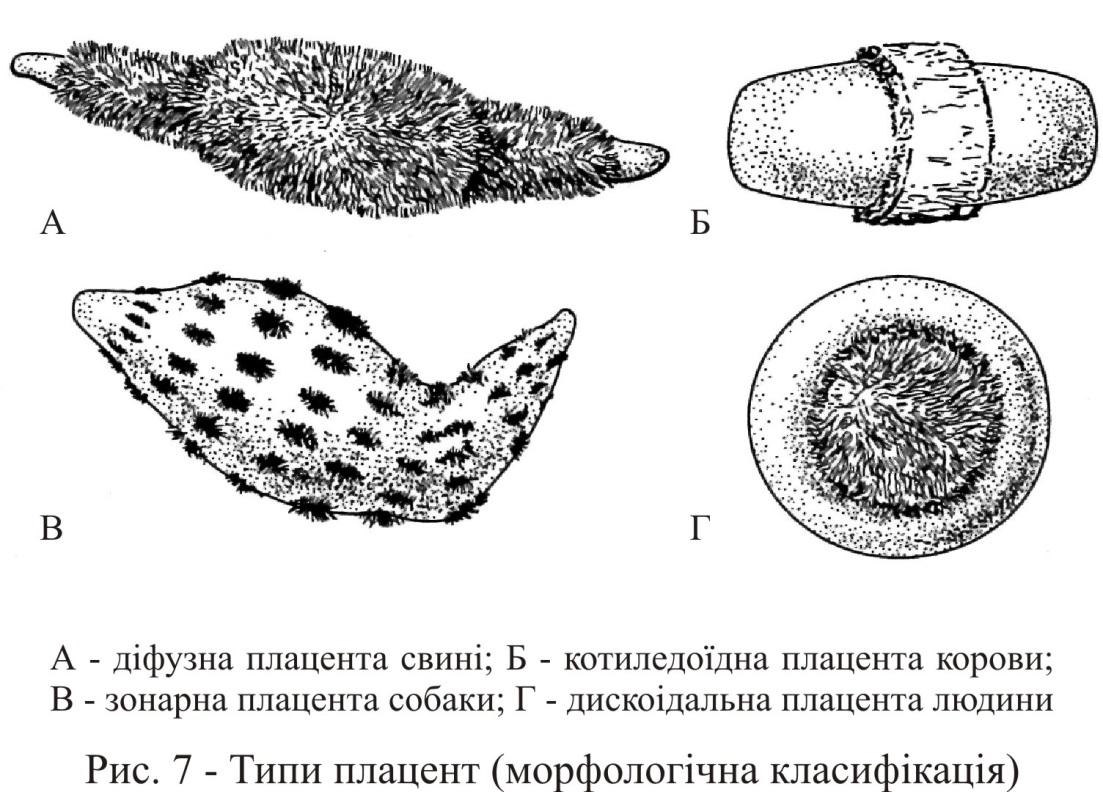
-
Функція газообміну між плодом і матір’ю. Посередником газообміну служить особливий фермент – трансферін. Він переносить молекули кисню від гемоглобіну материнської крові до гемоглобіну плоду. Останній (фетальний гемоглобін) має більшу спорідненість з киснем, ніж гемоглобін матері.
-
Функція живлення. На ранніх стадіях розвитку після імплантації ворсинки хоріону одержують поживні речовини від клітин слизової оболонки матки, зруйнованих при імплантації та утворенні плаценти (гістотрофіка). Потім основною функцією плаценти стає отримання поживних матеріалів з материнської крові (гемотрофіка). Крім поживних речовин через плаценту в кров плоду можуть поступати чужорідні речовини. При цьому відмічається висока вибірковість відносно проникнення різних речовин: одні з них проникають в кров плоду (наприклад, антибіотики тетрациклінового ряду), інші – ні.
-
Антитоксична функція. Плацента здатна видаляти ряд токсичних речовин та отрут як за рахунок діяльності ферментів (аналогічних ферментам печінки), так і просто шляхом викиду токсинів в кров матері. Плацента, як правило, захищає плід від інфекції – через неї не проходять бактерії, грибки, пухлинні клітини.
-
Гормоноутворююча функція. Хоріон плоду є потужною ендокринною залозою. Тут утворюються гонадотропін, адренокортикотропний гормон, соматотропін (сприяючий збільшенню маси матки й плоду) та інші гормони, що поступають в кров материнського організму. До плоду не поступають стероїдні гормони хоріону: естроген, андроген і прогестерон. В другій половині вагітності плацента стає основним джерелом гормонів, які забезпечують нормальне протікання вагітності й пологів.
-
Функція згортання крові. Плацента виділяє як речовини, сприяючі згортанню крові, так і фібрінолітичні речовини, що виключають утворення тромбів.
Контрольні питання:
-
Загальна характеристика нейруляції, її біологічний смисл.
-
Як проходить формування нейтральної пластинки?
-
Як утворюється нервовий жолобок та нейтральна трубка?
-
Які перетворення мезодерми проходять під час нейруляції?
-
Провізорні органи, їх походження та функції.
-
Плацента. Типи плацент. Функції плаценти.