

04_Droblennya_zv
.docxТема 5. Дроблення
-
Загальна характеристика дроблення, його біологічний смисл.
-
Значення кількості та розподілу жовтка.
-
Типи дроблення.
-
Морфологія основних типів дроблення.
-
Типи бластул, зв’язок їх будови з морфологією дроблення.
Основні поняття: дроблення, ділення дроблення, борозни дроблення, ядерно-цитоплазматичне співвідношення, бластомер, макромер, мікромер, мезомер, бластоцель, бластодерма, бластула, тип дроблення, тип бластули, трофобласт, жовточний мішок, амніон, хоріон, презумптивні зачатки.
Дроблення – це ряд безперервно слідуючих одне за одним мітотичних ділень зиготи, в результаті яких одна клітина – запліднене яйце – перетворюється на багатоклітинний комплекс. Після об’єднання хромосомних наборів обох пронуклеусів починається мітотичне ділення ядра зиготи. За цим першим розподілом слідує серія ділень, так званих ділень дроблення, які мають певні особливості:
-
клітини зародка, що розділилися, не ростуть, тобто в проміжку між розподілами їх маса не збільшується – сумарний об’єм і маса всіх виниклих клітин не перевищують об’єм й масу яйцеклітини відразу після запліднення;
-
кількість ДНК в ядрах подвоюється після кожного ділення, як при звичайному мітозі (таким чином, кількість ДНК, яке припадає на зародок, постійно збільшується).
Клітини, що утворюються підчас дроблення називають бластомерами, а вдавленія протоплазми – борознами дроблення. В одних тварин (наприклад, жаб) борозна дроблення з’являється в області анімального полюса та розповсюджується до вегетативного. Одночасно борозна заглиблюється до центральної частини яйця. Ще до закінчення першого дроблення на поверхні з’являється друга борозна дроблення. Таким чином, бластомери у внутрішніх своїх частинах якийсь час залишаються з’єднаними. В інших яєць борозна дроблення майже одночасно опоясує все яйце.
Борозни, що проходять через анімальний та вегетативний полюси зиготи, називаються меридіальними. Борозна, що проходить перпендикулярно полярній осі зиготи через її середину, називається екваторіальною; борозни, паралельні екваторіальній, називаються широтними; борозни, що проходять паралельно поверхні зародка, називаються тангенціальними.
Спочатку бластомери прилягають один до одного, утворюючи скупчення клітин – морулу. Потім між клітинами утворюється порожнина – бластоцель, заповнена рідиною. Клітини відтісняються до периферії, утворюючи стінку бластули – бластодерму. Період дроблення завершується стадією розвитку організму – бластулою. Одна з найважливіших особливостей періоду дроблення – відсутність росту організму, який розвивається, не дивлячись на ділення, що відбуваються. В онтогенії бластомерів немає періоду зростання, перед дробленням вони не збільшуються в розмірах.
Видові особливості процесу дроблення визначаються двома основними параметрами:
-
кількістю й розподілом жовтка в цитоплазмі;
-
присутністю в цитоплазмі чинників, які впливають на орієнтацію мітотичного веретена та час його утворення.
Дроблення закінчується, коли у зародка відновлюється рівновага між ядром і цитоплазмою.
Дроблення буває голобластичним (повним), меробластичним (неповним) та абластичним (поверхневим) (рис. 1).
Голобластичне дроблення характерно для яйцеклітин, які мають незначну кількість жовтка, тобто для оліго-, мезоліцетальних та помірно телоліцетальних яйєць. Цей тип дроблення характеризується тим, що весь вміст зиготи входить до складу бластомерів, а зародок розвивається за рахунок внутріклітинної утилізації жовтка. Повне дроблення може бути повним рівномірним (оліголецитальні яйцеклітини) та повним нерівномірним (мезолецитальні яйцеклітини).
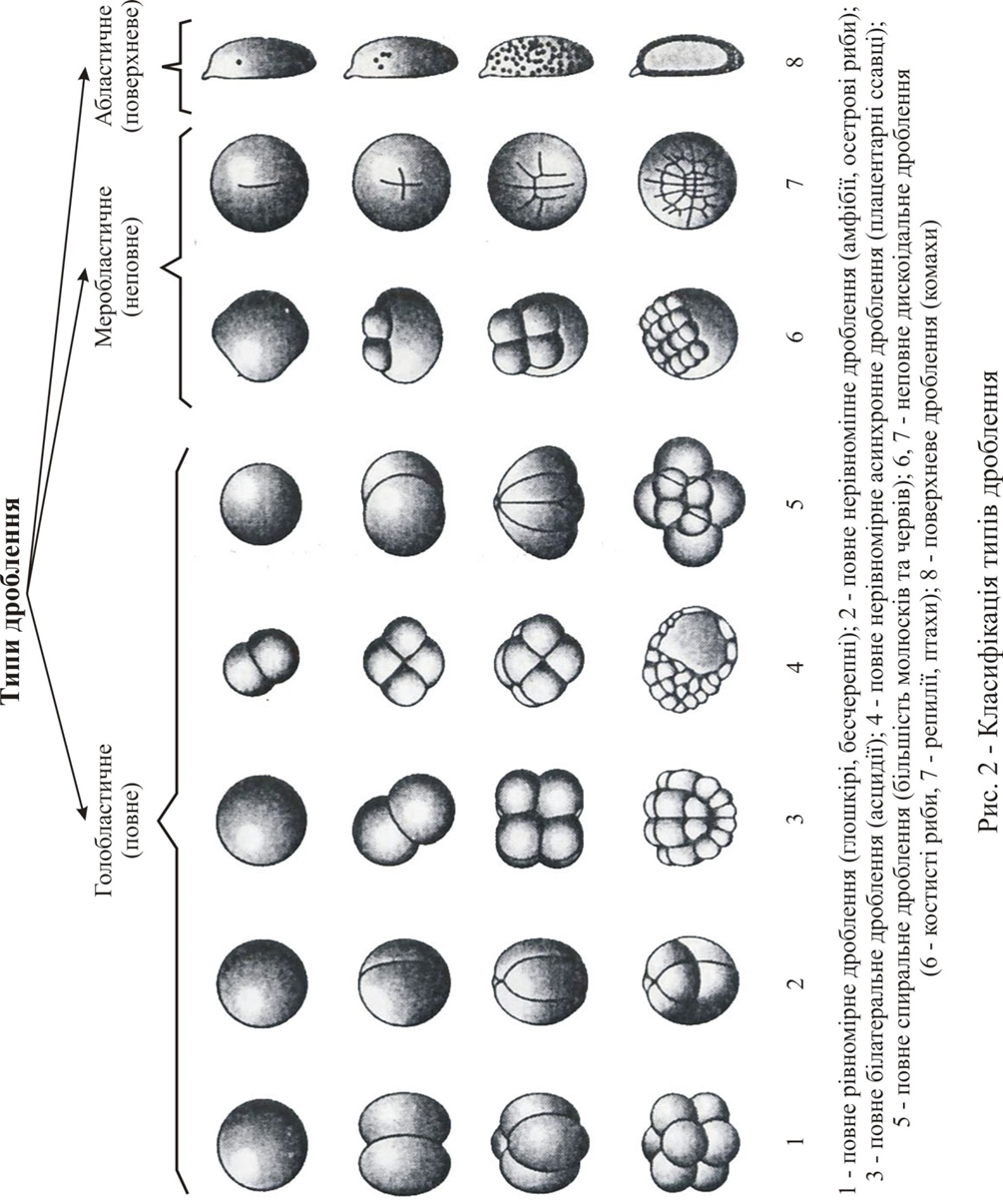
Загальною закономірністю голобластичного дроблення є взаємна перпендикулярність (ортогональність) перших трьох борозн дроблення.
Яйцеклітини з великою кількістю жовтка (полілецитальні) зазнають неповне, або меробластичне, дроблення. Цей тип дроблення характеризується тим, що лише з частини такої яйцеклітини формується зародок, а решта частини представляє поживний матеріал, що витрачається. Неповне дроблення може бути дискоідальним (телолецитальні яйцеклітини з великою кількістю жовтка) та поверхневим (центролецитальні яйцеклітини).
Абластичне дроблення характерно для центролецитальних яйєць комах. В цьому випадку цитокінез не відбувається та ділення цитоплазми відсутне. Діляться тільки ядра, які знаходяться у центрі яйця, звідки вони мігрують по цитоплазматичним тяжам на поверхню. Довгий час зародок має синцітіальну структуру. Потрапивши до поверхневої цитоплазматичної бластеми (періплазми), ядра утворюють синцітіальну бластодерму, яка пізднійше целюляризується та дає начало клітинній бластодермі зародка.
Характер дроблення залежить не тільки від кількості й розташування жовтка, але й від інших факторів. Тому дроблення також класифікують:
-
за об’ємом бластомерів, що утворюються:
-
рівномірне – об’єми бластомерів однакові;
-
нерівномірне – об’єми бластомерів неоднакові.
-
за тривалістю каріо- та цитотомії в різних бластомерах:
-
синхронне – дроблення починається і завершується у всіх бластомерах одночасно;
-
асинхронне – начало і час ділення в різних бластомерах неоднаково.
-
за взаємним розташування бластомерів:
-
радіальне – взаємне розташування бластомерів таке, що початкова полярна вісь яйця служить віссю радіальної симетрії зародка, що дробиться;
-
спіральне – прогресивне порушення симетрії яйця, що дробиться, в результаті спірального зміщення ділень бластомерів відносно один одного;
-
білатеральне – бластомери розташовані так, що через зародок можна провести тільки одну площину симетрії;
-
ротаційне – бластомери при другому дробленні діляться у взаємно перпендикулярних площинах;
-
анархічне – закономірність в розташуванні бластомерів у організмів одного виду відсутня.
У багатьох яйцеклітин ще на ранніх стадіях дроблення, як вже відмічалось раніше, внутрішні кінці бластомерів розходяться, і між ними порожнина дроблення – бластоцель. У деяких типів яйцеклітин бластоцель може досягати значних розмірів. В ході подальшого розвитку бластоцель перетворюється на первинну порожнину тіла, яка є основною порожниною тіла у нижчих безхребетних. У вищих безхребетних та хребетних вона майже повністю витісняється вторинною порожниною тіла (целомом), що виникає пізніше.
Бластоцель виконує дві функції:
-
дає можливість клітинам при гаструляции мігрувати всередину зародка;
-
існування бластоцелю запобігає взаємодії між клітинами, які знаходяться вище і нижче за нього.
Бластоцель – це перший відсік внутрішнього середовища організму, що виникає по ходу розвитку, та який відрізняється за іонним складом від зовнішнього середовища. Клітини стінок бластоцеля, що відгороджують його від зовнішнього середовища, утворюють між собою щільні контакти, які мають виборчу проникність для іонів. На зовнішній мембрані клітин, розташовані іонні канали для натрію, хлору та інших іонів, які забезпечують їх перенесення по градієнтах концентрації, а на внутрішній – іонні насоси, що працюють проти градієнта. В результаті Nа+ та Cl- перекачуються в бластоцель, де виникає їх надлишок, що створює в бластоцелі підвищений осмотичний тиск і забезпечує перенесення води, підвищуючи в ній тургорний тиск. Цей тиск розтягує поверхню зародка, що важливо для його наступного розвитку. Надлишок натрію в бластоцелі може впливати на швидкість клітинних циклів та стимулювати експресію генів в клітинах бластули.
Існує декілька типів бластул (рис. 2).
Целобластула характерна для голкошкірих та ланцетника. Ворма її сферична. Відрізняється цей тип бластул тим, що бластомери на анимальном і вегетативному полюсах абсолютно однакові за розміром, тонку бластодерму формує один шар циліндричних або кубічних клітин. Великий бластоцель займає центральне положення. Виділяють в целобластулі наступні зони: дах на анимальном полюсі, дно – на вегетативному, між ними знаходиться крайова або проміжна зона.
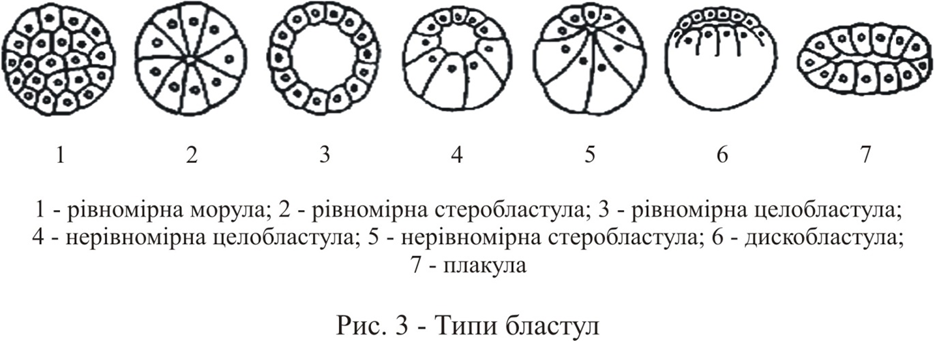
Бластула із стінкою рівномірної товщини і дуже маленьким центрально розташованим бластоцелем, яка зустрічається у деяких кишковопорожнинних, молюсків та черв’яків називається стеробластулою.
Целобластула и стеробластула можуть бути рівномірними та нерівномірними.
Зародковий розвиток деяких губок, черв’яків, асцидий призводить до утворення плакулы – двошарової пластинки, утвореної однорідними клітинами, між якими розташовується щілиноподібний бластоцель.
У деяких кишковопорожнинних, наземних олігозет, а також у ссавців на ранніх стадіях розвитку, бластоцель взагалі не виникає, і дроблення закінчується на стадії щільного скупчення клітин – морули.
Своєрідним типом бластули є стомобластула, характерна для зародкового розвитку вапняних губок. Вона має порожнину в центрі та отвір (фіалопор) на анимальном полюсі. Джгутикоутворюючі полюсы клітин обернені всередину. Після закінчення дроблення стомобластула вивертається навиворіт через фіалопор (екскурвація), внаслідок чого утворюється покрита джгутиками амфибластула.
Бластоциста характерна для ссавців. Утворюється після проходження стадії морули та має вигляд бульбашки, у складі якої є дві частини. Стінку формує один шар світлих клітин – трофобласт, а внутрішньоклітинну масу – темнозабарвлені клітини ембріобласт. Трофобласт забезпечує живлення, а з ембріобласту утворюється тіло зародка.
Амфибластула характерна для амфібій та деяких губок. Має сферичну форму. Бластомери різко розрізняються за розмірами та біохімічними властивостями вже на ранніх стадіях розвитку. На анимальном полюсі знаходяться менші за розміром клітини – мікромери, які пігментовані, мають меншу кількість жовтка, в порівнянні з розташованими в області вегетативного полюсу макромерами. На відміну від целобластули бластоцель в амфібластулі зміщено до анімального полюсу внаслідок більшої кількості жовтка на вегетативному. Бластодерма складається з декількох шарів клітин: анімальна частина стінки бластули (дах бластоцеля) тонша, в порівнянні з масивною вегетативною частиною (дно бластоцеля).
Дискобластула характерна для костистих риб, рептилій та птахів. В результаті дискоїдального дроблення утворюється одношарова бластодерма (бластодиск), з якого потім виникає 3-х або 4-х клітинний пласт. Бластодерма дискобластули (дах бластули) дещо вигинається над об’ємною масою жовтка, який не роздробився (дном бластули), а між ними формується бластоцель. На цій стадії чітко помітні дві області бластодиска: світле поле – відповідає тій його частині, яка розташовується над порожниною, темне поле – складається з клітин, розташованих по краю бластодиска та прилеглих до жовтка. До моменту відкладання яйця деякі з клітин бластодерми виселяються в підзародкову порожнину, де утворюють другий шар. Таким чином, на цій стадії зародок складається з епібласта (зовнішній шар клітин) та гіпобласта, що лежить під ним. Між ними знаходиться бластоцель.
Перібластула характерна для комах та є завершальною стадією поверхневого дроблення. Форма її еліпсоїдна. Порожнина відсутня. Бластодерма складаеться з олного, двох шарів клітин, які оточують жовток. Енергіди (ядра з острівцями цитоплазми, що їх оточують) мігрують на поверхню яйця, де вони оточуються новими плазматичними мембранами. Першими зміни зазнають ядра, які мігрували до заднього полюсу яйця та утворили полярні клітини зародка, які дають початок статевим клітинам дорослого організму. Клітини бластодерми утворюють скупчення на вентральній поверхні, означаючи місце, де виникнуть зародкові листки (зародкова смужка). Ця область дає початок усім клітинам комахи. Інші клітини бластодерми утворюють позазародкові оболонки.
Контрольні питання:
-
В чому полягають особливості дроблення?
-
Біологічний смисл дроблення.
-
Як впливає кількість та розподіл жовтка на дроблення?
-
За якими ознаками класифікують типи дроблення?
-
Дайте характеристику типам дроблення.
-
В чому полягають особливості голобластичного та меробластичного типів дроблення?
-
В чому полягає зв’язок будови бластул з морфологією дроблення?
-
В чому полягають особливості дроблення ссавців?