

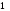
ЭМБРИОЛОГИЯ – наука об эмбриональном развитии организма как части онтогенеза человека.
Эмбриональный период включает время от момента оплодотворения яйцеклетки сперматозоидом и образования зиготы до рождения.
Стадии эмбриогенеза:
оплодотворение - цепь событий, которая завершается процессом слияния мужской и женской половых клеток и приводит к образованию одноклеточного зародыша – зиготы;
дробление; в результате делений дробления образуется многоклеточная бластула;
гаструляция → результат гаструляции образование зародышевых листков, организм на этой стадии развития называется гаструла; зародыш приобретает многослойное строение;
нейруляция и нотогенез → образование из зародышевых листков осевого комплекса зачатков органов;
гистогенез → формирование тканей;
органогенез → формирование органов;
системогенез → формирование систем органов.
Оплодотворению предшествует процесс образования мужских и женских половых клеток – ГАМЕТОГЕНЕЗ (ПРОГЕНЕЗ).
ГАМЕТОГЕНЕЗ
Первичные половые клетки гоноциты (гонобласты) впервые выявляются в стенке желточного мешка (в конце третьей недели жизни эмбриона человека). Гоноциты являются индифферентыми клетками: они не разделяются на мужские и женские клетки.
Гоноциты мигрируют в закладки половых желез. По пути они многократно делятся.
Гоноциты дифференцируются в сперматогонии (в семеннике) или в овогонии (в яичнике). Это процесс начинается на 6-ой недели жизни эмбриона.
СПЕРМАТОГЕНЕЗ происходит в извитых канальцах семенника.
В процессе сперматогенеза выделяют 4 фазы (периода):
размножение (пролиферация)
рост
созревание
формирование (спермиогенез).
Фаза размножения:
Мужские половые клетки представлены диплоидными клетками – СПЕРМАТОГОНИЯМИ, которые делятся МИТОТИЧЕСКИ.
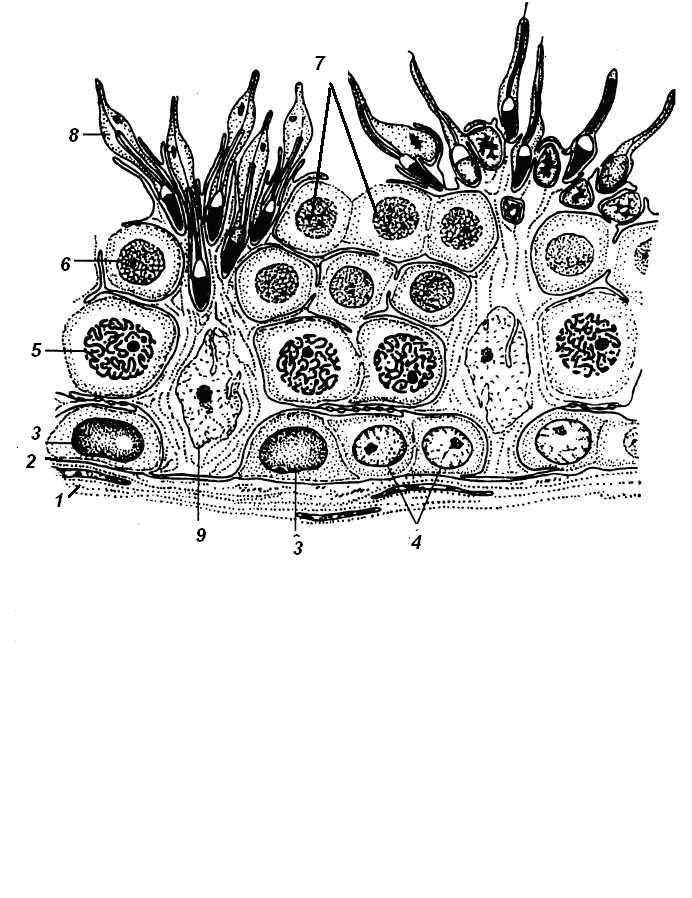
Рис.1. Участок стенки извитого канальца семенника (по Карлсон, 1983)
1 – миоидные клетки стенки; 2 – базальная мембрана; 3 - сперматогонии А;
4 – сперматогонии В; 5 – сперматоцит первого порядка; 6 – сперматоцит второго порядка; 7 – сперматиды на ранней стадии развития; 8 – сперматиды в конце развития; 9 – клетка Сертоли.
Различают 3 типа сперматогоний (рис.1):
тёмные А сперматогонии – истинные стволовые клетки, устойчивы к действию вредных факторов, делятся редко;
светлые А сперматогонии – полустволовые клетки, способны к частым митотическим делениям (результат деления: образуются две А- сперматогонии, либо одна А и одна В-).
светлые В сперматогонии, коммитированные клетки – готовятся к мейозу и вступают в фазу роста.
II. Фаза роста
В сперматогонии увеличиваются в объёме (в 4 раза) и вступают в профазу первого мейотического деления, перемещаясь из базального слоя по направлению к просвету канальца → СПЕРМАТОЦИТЫ 1-го порядка (диплоидные).
III. Фаза созревания
В результате 1-го деления мейоза (редукционного) образуются СПЕРМАТОЦИТЫ 2-го порядка, которые быстро вступают во 2-е деление мейоза (эквационное), образующиеся клетки называются СПЕРМАТИДЫ (гаплоидные).
Начиная со сперматогоний типа В, дочерние клетки при делении не отделяются полностью друг от друга, а остаются связанными с помощью цитоплазматических мостиков, образуя синцитий. Лишь зрелые сперматозоиды отделяются от синцития.
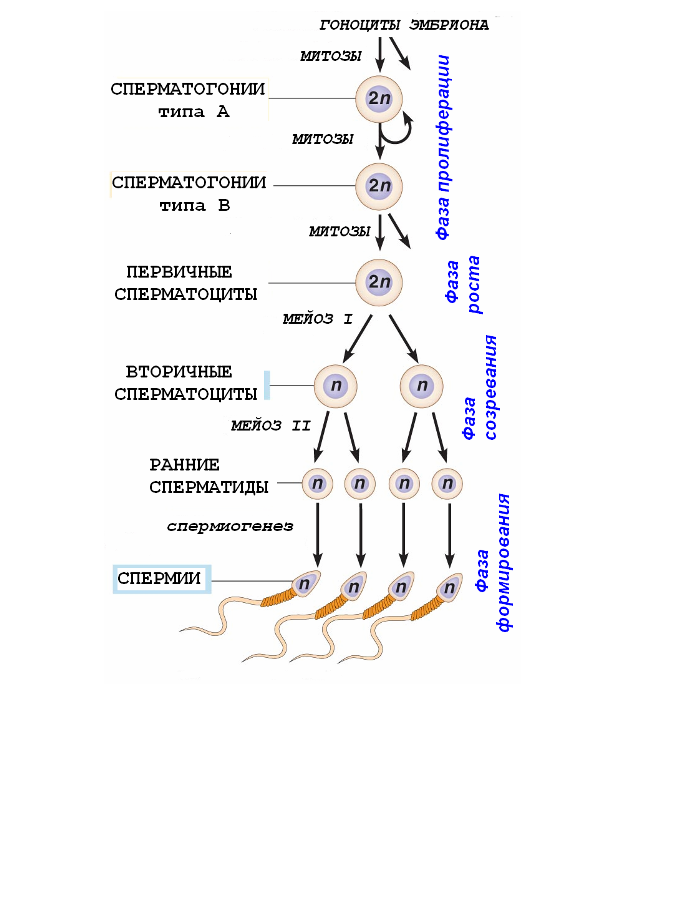
Рис.2. Фазы сперматогенеза
IY. Фаза формирования (спермиогенез)
Самая продолжительная фаза сперматогенеза (около 50 суток). В процессе формирования зрелых сперматозоидов из сперматид происходят (Рис.3):
конденсация ядерного хроматина; конденсация хроматина приводит к образованию суперкомпактного ядра, что увеличивает мобильность сперматозоида и защищает геном от повреждения;
образование акросомы из элементов комплекса Гольджи; акросома - специальный тип лизосомы, содержит гидролитические ферменты (такие как гиалуронидаза, нейраминидаза, кислая фосфатаза, протеазы) для разрушения оболочек яйцеклетки; акросома в виде «шапочки» покрывает переднюю половину ядра; наружная мембрана акросомы соприкасается с клеточной мембраной, а внутренняя – с мембраной ядра.
центриоли перемещаются к противоположному полюсу ядра; от дистальной центриоли образуется осевая нить (аксонема) жгутика.
митохондрии в виде спирали окружают часть аксонемы, образуя утолщенный участок – промежуточный отдел; митохондрии обеспечивают энергию для движения сперматозоида;
цитоплазма редуцируется до минимума;
после сброса излишек цитоплазмы, сперматозоиды отделяются от общей ассоциации клеток - синцития - и становятся свободными.
Рис.3. Важнейшие изменения, происходящие в сперматидах в период формирования
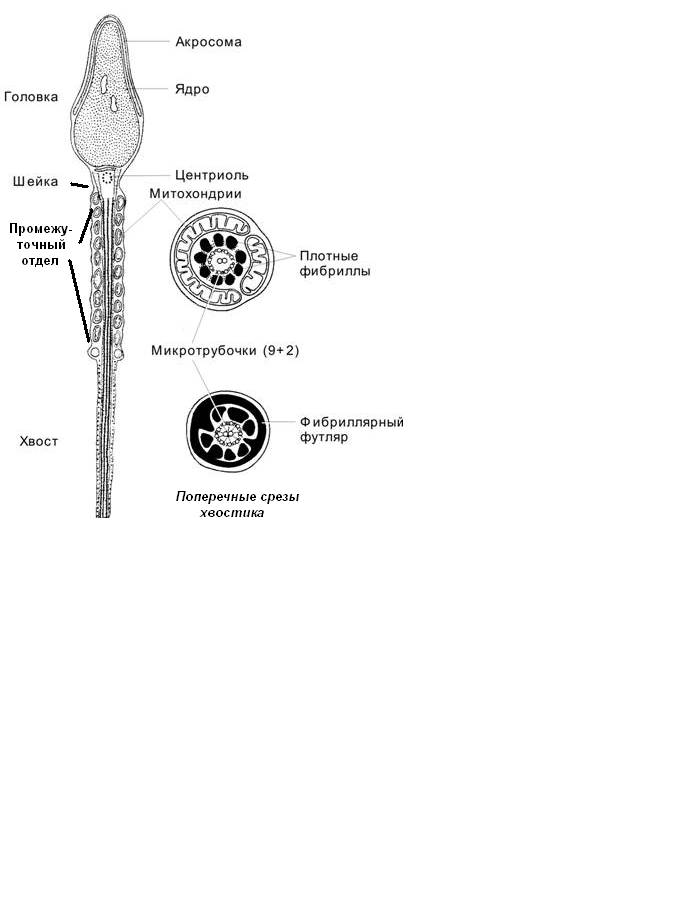
В ЗРЕЛОМ СПЕРМАТОЗОИДЕ выделяют (рис.4):
головку; содержит ядро и акросому.
шейку (связующий от
 дел),
где расположена центриоль;
дел),
где расположена центриоль;хвостик: в промежуточном отделе аксонему окружает митохондриальная спираль;
Рис.4.Схема строения сперматозоида
ОВОГЕНЕЗ
Образование женских половых клеток – овогенез – происходит в яичнике.
ОВОГЕНЕЗ включает 3 фазы:
размножение
рост
созревания
I. Фаза размножения
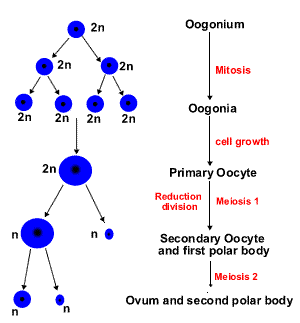
В отличие от сперматогенеза, фаза размножения в овогенезе протекает исключительно в эмбриональный период развития. Овогонии делятся путём митозов. Большинство образующихся дочерних клеток погибает (этот процесс называется атрезия), часть вступает в фазу роста (в профазу первого деления мейоза). Митотические деления овогоний прекращаются к 7 месяцу эмбриогенеза. При рождении все яйцеклетки в яичнике являются овоцитами первого порядка (первичными овоцитами).
II. Фаза роста
Фаза роста соответствует жизни первичного овоцита, находящегося в профазе первого деления мейоза. Фаза роста делится на два периода: малого (до полового созревания) и большого роста.
К моменту рождения в яичнике насчитывается около 2 млн первичных овоцитов; к периоду полового созревания большинство из них погибает, остаётся около 400 тысяч яйцеклеток.
Рост овоцитов происходит в фолликулах – структурах, в которых овоцит окружен эпителиальными фолликулярными клетками. В период малого роста имеется два типа фолликулов:
примордиальные фолликулы (овоцит окружен одним слоем уплощенных фолликулярных клеток) и
первичные фолликулы (появляется блестящая (прозрачная оболочка), фолликулярные клетки имеют кубическую или призматическую форму).
Вступление овоцитов в период большого роста происходит только под воздействием гормонов (фолликуло-стимулирующего гормона); этому периоду оогенеза соответствуют растущие (вторичные и третичные) фолликулы.
Период роста может продолжаться от 12 до 50 лет.
III. Фаза созревания
Первое деление мейоза завершается только перед овуляцией (овуляция -разрыв зрелого третичного фолликула и выброс яйцеклетки из яичника в брюшную полость). В результате мейоза I образуется овоцит второго порядка (вторичный овоцит) – гаплоидная клетка. Овоцит второго порядка вступает во второе деление мейоза, но это деление блокируется на стадии метафазы и завершается только при условии оплодотворения.
За один менструально-овариальный цикл (~ 28 дней) происходит овуляция обычно одной яйцеклетки. Таким образом, за репродуктивный период жизни женщины (~ 30-40 лет) только около 450 яйцеклеток достигают зрелости – готовности к оплодотворению.
Мейотическое деление в овогенезе отличается тем, что протекает с неравной цитотомией: в мейозе I хромосомы поровну распределяются между дочерними клетками, но только одна из клеток (вторичный овоцит) сохраняет почти всю цитоплазму; вторая клетка (первое полярное, или редукционное, направительное тельце) имеет очень мелкие размеры, небольшое ядро, минимум цитоплазмы.
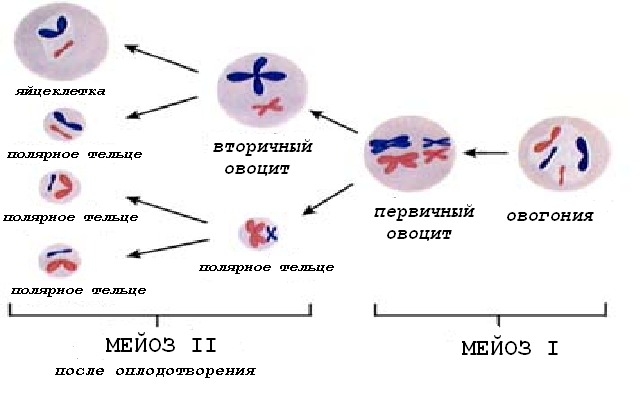
Рис.5. Фазы овогенеза
Зрелая яйцеклетка млекопитающих – крупная, в сравнении со сперматозоидом, неподвижная клетка. Особенность строения яйцеклетки – отсутствие центриолей и наличие характерных трофических включений – желточных гранул. По периферии яйцеклетки, под клеточной мембраной, равномерно распределены кортикальные гранулы, содержащие ферменты (гидролазы), необходимые для формирования оболочки оплодотворения после проникновения сперматозоида.
При овуляции яйцеклетка окружена (рис.6):
прозрачной оболочкой (zona pellucida), содержащей густую сеть тонких нитей, состоящих из гликопротеинов (фракции ZP1, ZP2, ZP3);
фолликулярными клетками, образующими лучистый венец (corona radiata).
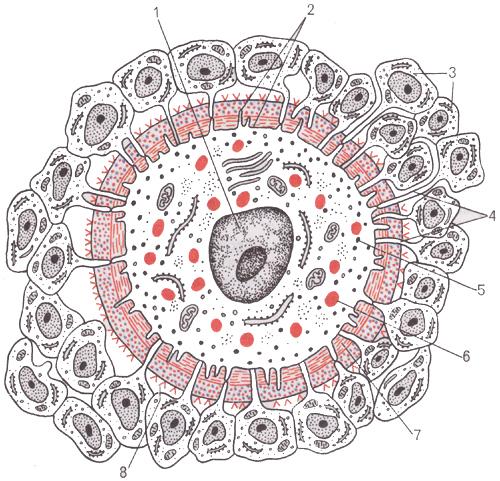
Рис.6. Строение яйцеклетки млекопитающего (по.
1 – ядро; 2 – 3 4 5 6 7 8
Основные различия в процессах сперматогенеза и овогенеза представлены в таблице 1.
Таблица 1
Основные различия процессов сперматогенеза и овогенеза
|
СПЕРМАТОГЕНЕЗ |
ОВОГЕНЕЗ |
|
Фаза размножения происходит только с момента полового созревания и продолжается в течение всей жизни |
Фаза размножения происходит только в эмбриональном периоде |
|
Фаза роста следует сразу за фазой размножения, относительно короткая (дни) |
Фаза роста очень длительная (от 10 до 50 лет); блок на стадии профазы мейоза I; подразделяется на малый рост (до полового созревания) и большой рост. |
|
Фаза созревания характеризуется равномерным делением сперматоцитов: 1 сперматогония даёт начало 4 зрелым сперматозоидам |
Фаза созревания характеризуется неравномерным делением цитоплазмы овоцитов: 1 оогония даёт начало 1 зрелой яйцеклетке и 3 полярным (редукционным) тельцам |
|
Есть фаза формирования |
Фаза формирования отсутствует; зрелая ♀яйцеклетка - в метафазе мейоза II |
|
Продолжается в течение всей жизни |
Прекращается после менопаузы |
ОПЛОДОТВОРЕНИЕ
Оплодотворение - цепь событий, завершающийся процессом слияния сперматозоида и яйцеклетки, который приводит к образованию одноклеточного зародыша – зиготы.
При овуляция вторичный овоцит, окруженный прозрачной оболочкой и лучистым венцом, выбрасывается из яичника в брюшную полость и попадает в ампулу маточной трубы, в которой может произойти оплодотворение. Если этого не происходит в течение первых 24 часов после овуляции, овоцит дегенерирует.
В эякуляте человека содержится 3х108 сперматозоидов. В женских половых путях они сохраняют способность к оплодотворению до двух суток. Только 200 из них достигают ампулярной части маточной трубы (яйцевода), где и происходит оплодотворение.
ФАЗЫ ОПЛОДОТВОРЕНИЯ:
дистантное взаимодействие;
контактное взаимодействие;
проникновение сперматозоида в яйцеклетку;
слияние пронуклеусов и образование зиготы
Процесс оплодотворения длится от 12 до 24 часов.
Дистантное взаимодействие
Сперматозоиды направленно мигрируют к воронке маточной трубы, где и происходит встреча с яйцеклеткой. При этом важную роль играют сокращения мышечных волокон стенки маточной трубы и хемотаксис. Хемоаттрактанты попадают в маточную трубу в составе фолликулярной жидкости лопнувшего фолликула, секретируются яйцеклеткой (гамоны) и фолликулярными клетками corona radiata.
В женских половых путях под влиянием факторов женского организма (слизистая секреция, рН и др.) происходит капацитация – физиологический процесс приобретения сперматозоидом способности к оплодотворению: окончательное созревание и активация. Во время капацитации происходят:
удаление гликопротеинов с цитолеммы головки сперматозоида;
перестройка цитолеммы (она становится более «текучей», «открытой» для проникновения ионов Са), что приводит к гиперактивации и усилению подвижности сперматозоида.
Для завершения капацитации требуется около 7 часов. Капацитацию можно воспроизвести in vitro.
Контактные взаимодействия
Для того чтобы произошло оплодотворение, сперматозоид должен последовательно преодолеть три барьера:
лучистый венец, из нескольких слоёв рыхло расположенных фолликулярных клеток;
прозрачную оболочку;
плазматическую мембрану (оолемму) яйцеклетки.
Сперматозоид довольно легко преодолевает лучистый венец, поскольку между его клетками нет прочных связей.
Более существенным барьером для сперматозоида является блестящая оболочка. При контакте с блестящей оболочкой сначала происходит связывание и прикрепление сперматозоида с рецепторами блестящей оболочки ZP2,. Благодаря рецепторам это прикрепление является видоспецифическим: сперматозоид и яйцеклетка должны обязательно принадлежать к одному виду млекопитающего.
После прочного связывания сперматозоида с ZP3 наступает акросомная реакция: мембрана акросомы во многих местах сливается с плазматической мембраной спермия в передней части головки, ферменты акросомы выделяются и расщепляют компоненты прозрачной оболочки, обеспечивая проникновение сперматозоида через образовавшийся канал к плазматической мембране яйцеклетки. Активные движения хвостика позволяют сперматозоиду мигрировать через прозрачную оболочку за 5-10 мин.
Касание одного из сперматозоидов оолеммы (плазматической мембраны яйцеклетки) приводит к образованию на поверхности воспринимающего бугорка. Оолемма содержит систему рецепторов, благодаря которым становится возможным слияние плазматических мембран головки сперматозоида и яйцеклетки, в результате чего головка спермия внедряется в ооплазму (в ооплазму входят только ядро сперматозоида и его центриоли).
Одновременно с началом взаимодействия двух гамет в яйцеклетке происходит кортикальная реакция, запускаемая быстрым повышением концентрации ионов кальция. Содержимое кортикальных гранул яйцеклетки выделяется в перивителлиновое пространство (пространство между оолеммой и блестящей оболочкой). Ферменты кортикальных гранул расщепляют рецепторы ZP2 и модифицируют ZP3, при этом блестящая оболочка теряет способность связывать другие спермии и блокирует акросомную реакцию других спермиев – формируется твердая оболочка оплодотворения, непреодолимая для спермиев. Оолемма также приобретает новые свойства, в том числе отрицательный заряд, отталкивающий спермии. Таким образом, оболочка оплодотворения обеспечивает блок полиспермии. Предупреждение полиспермии создаёт условия для наступления нормальной беременности.
Проникновение сперматозоида запускает завершение 2-го мейотического деления овоцита. В телофазе II формируются ядерные оболочки и образуются два гаплоидных ядра. Одно из них, окруженное тонким слоем цитоплазмы отделяется от овоцита в виде второго полярного тельца. Другое ядро, оставшееся в овоците, теперь называется женским пронуклеусом. Ядро проникшего сперматозоида также претерпевает ряд преобразований и набухает, формируя мужской пронуклеус. Оба пронуклеуса и центриоли сперматозоида перемещаются к центру ядра. В каждом из них начинается подготовка к митозу (конденсация и перестройка хроматина), происходит удвоение ДНК. После сближения их ядерные оболочки исчезают, а уже удвоенные хромосомы оказываются лежащими в цитоплазме. Формируется общее митотическое веретено и начинается первое деление дробления.
Результат оплодотворения – образование зиготы.
ЗИГОТА - оплодотворенная яйцеклетка и (или) новый организм, возникающий при слиянии двух гамет на одноклеточной стадии развития.
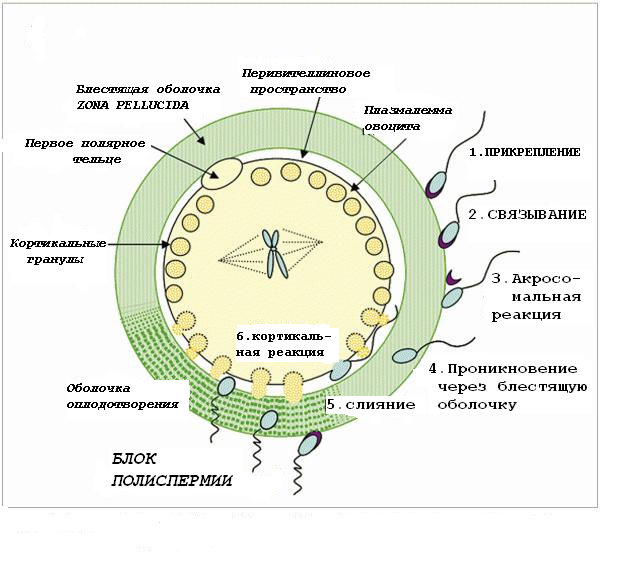
Рис.8. Последовательность контактных взаимодействий в процессе оплодотворения
БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ОПЛОДОТВОРЕНИЯ
Восстановление диплоидного набора хромосом;
Зигота содержит новую комбинацию хромосом и генетического материала, отличного от родительского;
Определение пола (в зависимости от оплодотворения сперматозоидом, несущего Х- или Y-хромосому);
Проникновение сперматозоида - сигнал для завершения 2-го мейотического деления овоцита;
Оплодотворение инициирует ряд быстрых митотических делений, называемых делениями дробления.
ТИПЫ ЯЙЦЕКЛЕТОК
Размеры и строение яйцеклеток различных представителей животного мира обусловлены количеством накопленных в цитоплазме питательных веществ, необходимых для формирования и роста зародыша. Эти питательные вещества содержатся в виде желточных включений (комплекса белков, липидов и углеводов). Количество желтка в цитоплазме яйцеклетки различно в зависимости от условий развития. Поскольку желток тормозит или даже препятствует делению цитоплазмы, количество и распределение желтка определяет тип делений дробления и образования бластулы.
Таким образом, классификации яйцеклеток основывается на количестве и характере распределения желточных включений в цитоплазме яйцеклетки.
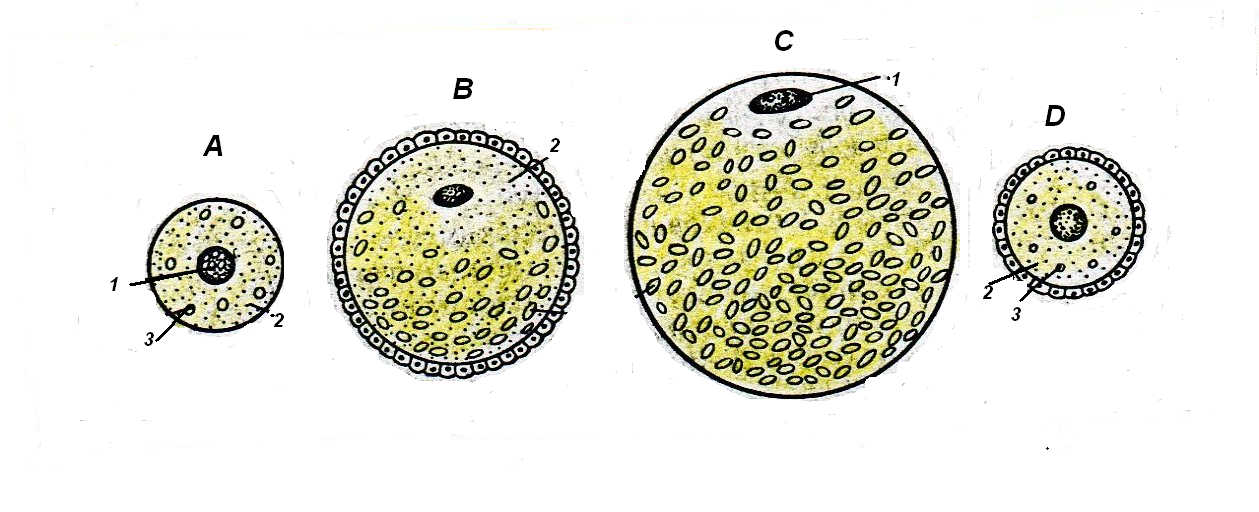
У различных животных в зависимости от количества питательных веществ в цитоплазме яйцеклетки могут быть (рис.7):
алецитальными – практически не содержат желтка;
олиголецитальными – с малым количеством желтка;
полилецитальными – с большим количеством желтка.
В олиголецитальных яйцеклетках небольшое количество желтка равномерно распределено в цитоплазме; такие яйцеклетки называются изолецитальными.
Полилецитальные яйцеклетки содержат асимметричное скопление желтка у одного из полюсов яйцеклетки, называемого вегетативным, ядро располагается ближе к противоположному анимальному полюсу. Эта полярность в распределении желтка может быть более умеренной - в мезолецитальных яйцеклетках (у амфибий) и более выраженной – в телолецитальных яйцеклетках (у рептилий, птиц).
В центролецитальных яйцеклетках желток равномерно распределён по всей цитоплазме, за исключением её периферического слоя (у многих членистоногих).
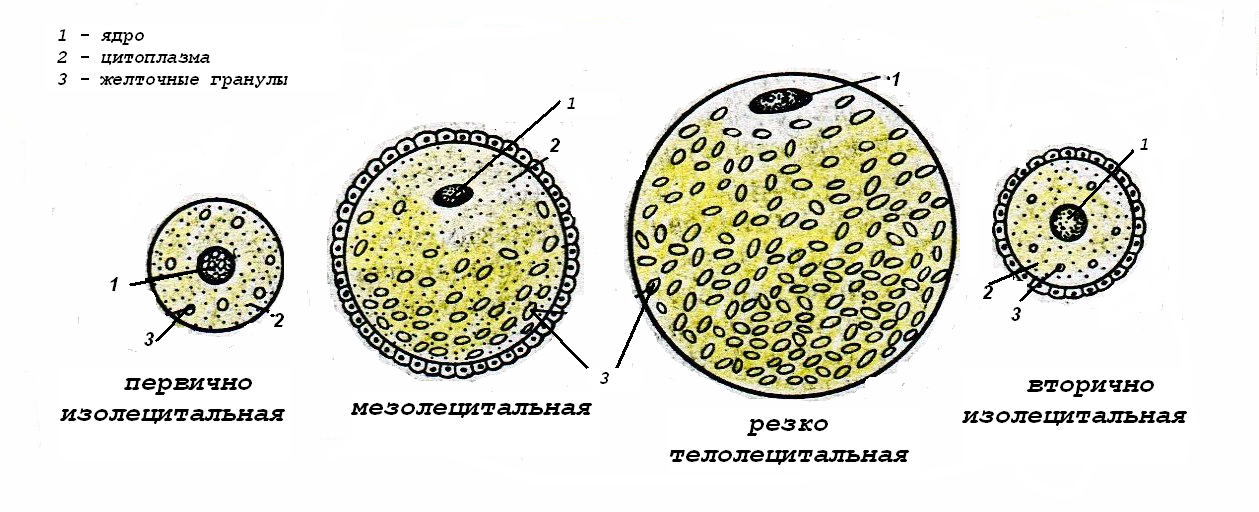
Рис.9. Типы яйцеклеток
У примитивных хордовых типа ланцетника, которые ведут водный способ существования и имеют стадию личинки – яйцеклетка первично изолецитального типа.
У некоторых рыб и амфибий, с развитием через личиночную стадию, яйцеклетки имеют умеренный запас питательных веществ – мезолецитальные яйцеклетки.
У рептилий и птиц, развитие которых происходит на суше и для развития требуется большой запас питательных веществ, яйцеклетки крупные, богатые желтком, - резко телолецитальные яйцеклетки.
В филогенезе предки высших млекопитающих также имели телолецитальные яйцеклетки, но в связи с переходом на внутриутробное развитие плода, к которому все необходимые вещества поступают из крови матери через плаценту, количество желтка было редуцировано, и яйцеклетка высших млекопитающих, включая человека, относится к вторично изолецитальному типу.
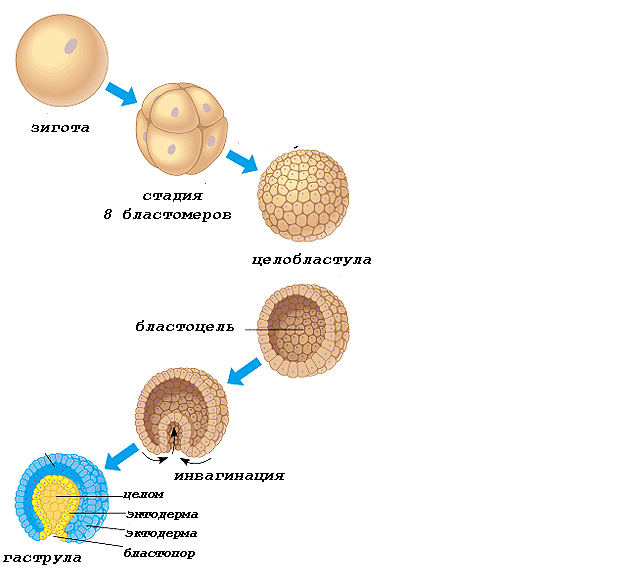
Дробление – стадия развития зародыша, которая следует за оплодотворением и включает в себя ряд последовательных митотических делений, называемых делениями дробления. За время короткой интерфазы происходит в основном только удвоение ДНК. РНК и белки практически не синтезируются (G1 фаза отсутствует). Клетки, образующиеся при делениях дробления, называются бластомерами.
Дробление происходит до тех пор, пока бластомеры не достигнут размеров, обычных для соматических клеток данного организма. Результат дроблений – образование бластулы – многоклеточного зародыша.
У разных видов животных процессы дробления характеризуются рядом видовых особенностей, обусловленных строением яйцеклетки.
Дробление может быть:
полным (голобластическим), когда весь объём ооцита делится на бластомеры;
неполным (меробластическом, частичным), когда делится только часть ооцита, его анимальный полюс, относительно свободный от желтка (характерно для резко телолецитальной яйцеклетки)
Кроме того, дробление может быть:
равномерным – если все бластомеры одинаковы;
неравномерным – если бластомеры, образующиеся при дроблении имеют неравные размеры
Тип бластулы зависит от типа дробления.
При полном равномерном синхронном дроблении первично изолецитальной яйцеклетки Ланцетника образуется целобластула (рис.10):
стенка целобластулы – бластодерма образована одним слоем клеток (однослойная);
бластомеры примерно одинакового размера;
в середине целобластулы есть полость – бластоцель.
Рис.10. Ранние стадии развития ланцетника
В результате полного неравномерного дробления мезолецитальной яйцеклетки амфибий (лягушки) образуется Амфибластула (рис.11):
на анимальном полюсе, которое содержало небольшое количество желтка клеточные деления происходят с большой скоростью и образуются мелкие бластомеры – микромеры;
на вегетативном полюсе, богатом желтком, клетки де
 лятся
медленно и образуются крупные бластомеры
– макромеры;
лятся
медленно и образуются крупные бластомеры
– макромеры;бластодерма (стенка бластулы) многослойная;
бластоцель смещён к анимальному полюсу;
анимальный полюс с многочисленными микромерами называется крышей бластулы, вегетативный с с макромерами образует дно; между ними краевая зона.
Тип дробления у птиц с полилецитальной резко телолецитальной яйцеклеткой - неполное (частичное) дискоидальное: деление ограничено небольшим участком на анимальном полюсе, остальная часть желтка остаётся нераздробленным. Образуется дискобластула (рис.12):
бластодерма многослойна, имеет форму круглой пластинки (диска) на анимальном полюсе;
щелевидный бластоцель находится между бластодермой и синцитиальном слоем на поверхности желтка (перибласт).
Характер дробления вторично изолецитальной яйцеклетки человека – полное неравномерное асинхронное. Образующая бластула на ранней стадии носит название морула (до стадии 64 бластомеров), а после появления полости – бластоцеля – бластоциста (рис.)
ГАСТРУЛЯЦИЯ
Гаструляция – это следующий за дроблением период развития зародыша. Скорость клеточных делений снижается, интерфаза между митозами удлиняется, а сами клетки начинают активно синтезировать РНК и белки.
Главная особенность этого периода – направленные активные перемещения делящихся клеток (клеточных потоков), которые приводят к глубокой перестройке зародыша и в конечном итоге к образованию трёх зародышевых листков:
эктодерма – наружный зародышевый листок;
энтодерма – внутренний зародышевый листок;
мезодерма - средний зародышевый листок, лежащий между экто- и энтодермой
Способы гаструляции у животных разных систематических групп сильно варьируют.
Описано четыре основных типа морфогенетических движений гаструляции:
инвагинация;
эпиболия;
деламинация;
иммиграция
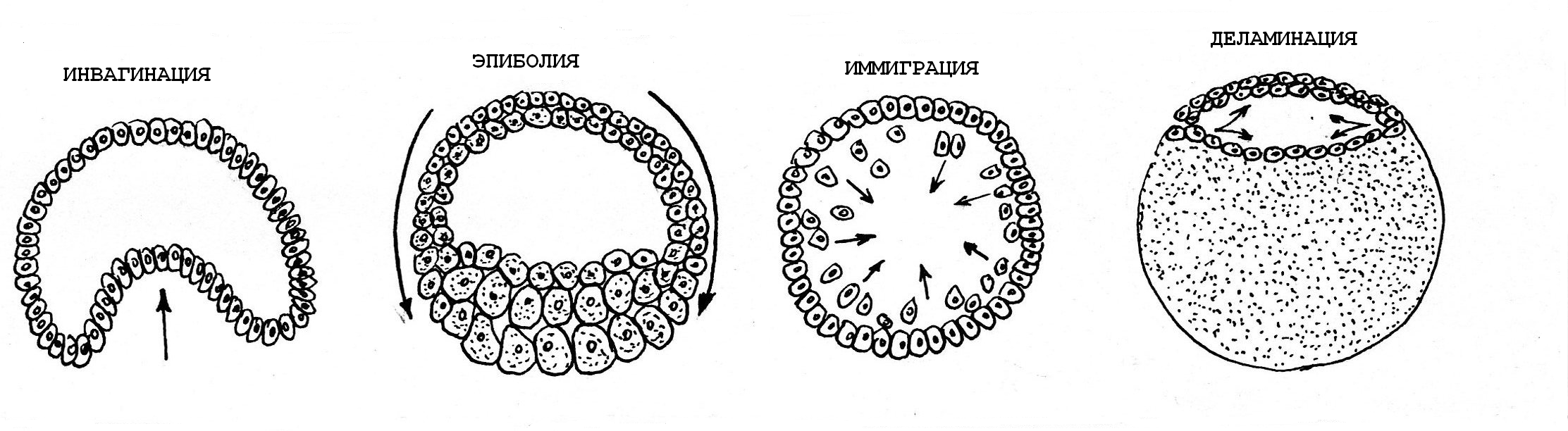
Рис.13. Основные механизмы гаструляции
Инвагинация – это впячивание бластодермы вегетативного полюса бластулы в полость бластоцеля, при этом образуется первичная кишка с полостью – гастроцелем, который сообщается с наружной средой отверстием – бластопором (первичным ртом). Инвагинация происходит при гаструляции у примитивных хордовых, таких как Ланцетник (рис.10).
Эпиболия – перемещение активно делящихся мелких клеток (микромеров) анимальной части бластодермы в направлении вегетативного полюса. Микромеры обрастают крупные клетки (макромеры) вегетативного полюса, и макромеры погружаются внутрь зародыша. Эпиболия, как тип движений гаструляции, характерна для эмбриогенеза амфибий (рис.11).
Деламинация – расщепление (расслоение) бластодермы на два пласта: наружный – эпибласт и внутренний – гипобласт (рис.14)
Иммиграция – перемещение отдельных клеток из бластодермы внутрь зародыша.
Если небольшие участки определенной области поверхности бластулы окрасить прижизненным красителем, можно проследить дальнейшую судьбу, дифференцировку клеток этого участка. Экспериментально показано, что клетки анимального полюса – клетки будущей эктодермы. По результатам таких исследований можно составить карту презумптивных зон.
ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ ПТИЦ
Тип яйцеклетки у птиц – полилецитальная, резко телолецитальная яйцеклетка.
Тип дробления зиготы - неполное (частичное, меробластическое) дробление (дробление ограничено лишь малой частью зиготы на анимальном полюсе, остальная часть зиготы не подвергается делению – нераздробившийся желток).
Тип бластулы – дискобластула: бластомеры располагаются в форме диска, лежащего на массе желтка (на перибласте).
Гаструляция в дискобластуле происходит в две фазы:
деламинация
иммиграция
Первая фаза гаструляции - деламинация заключается в расщеплении клеточной массы бластодиска на 2 листка: гипобласт (первичная энтодерма) и эпибласт (первичная эктодерма) (Рис.14).
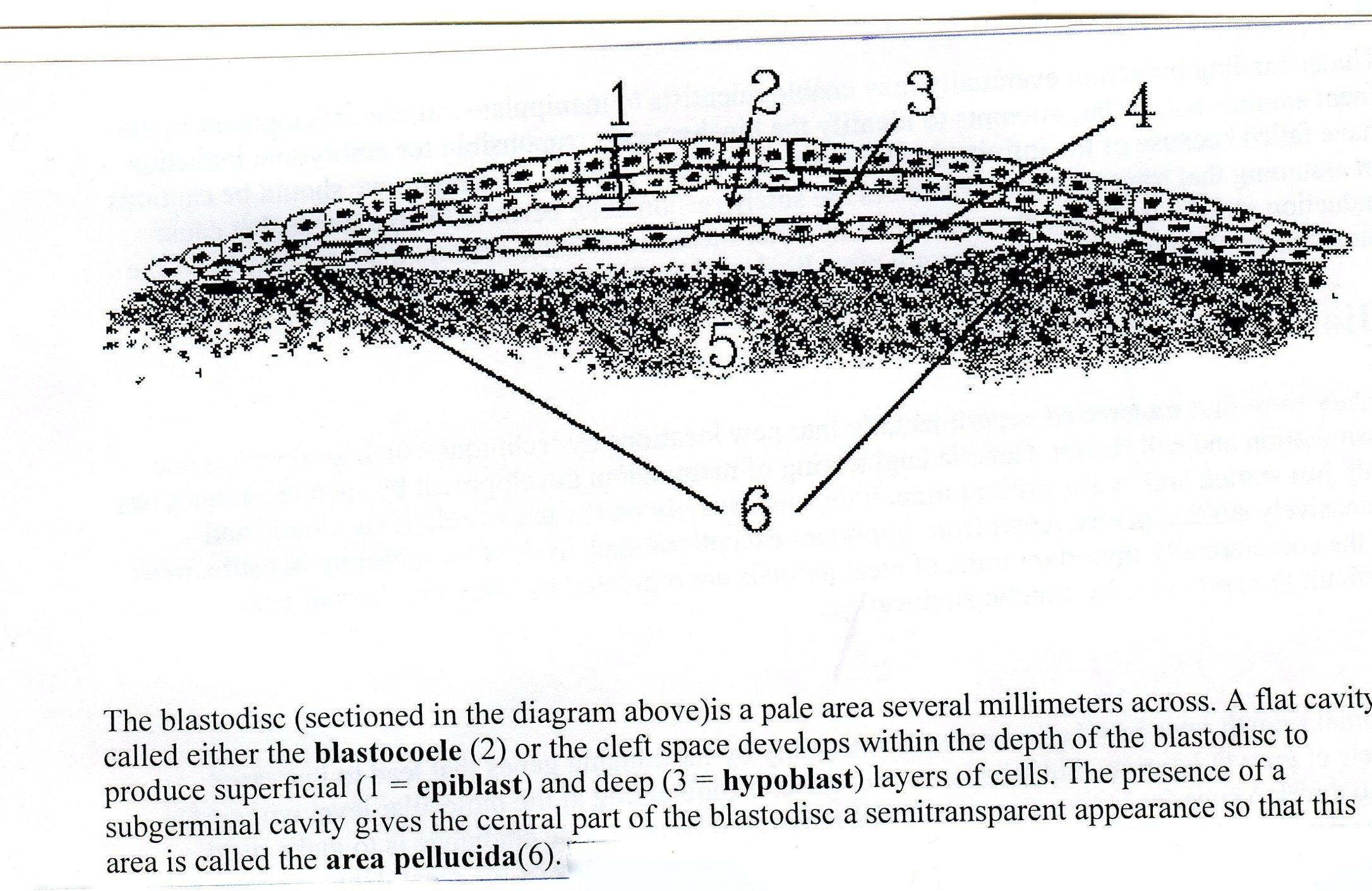
Рис.14. Деламинация:
эпибласт; 2. бластоцель; 3. гипобласт; 4. 5. – нераздробившийся желток.
Клетки внутреннего листка - гипобласта - смыкаются в правильную однослойную пластинку. Наружный слой бластодиска – эпибласт – клеточный пласт из более высоких призматических клеток. Центральный утолщенный участок бластодиска, состоящий из эпибласта и гипобласта, называется зародышевым щитком. Из зародышевого щитка образуется тело самого зародыша.
Вторая фаза гаструляции – иммиграция связана со сложным перемещением клеток эпибласта сначала к заднему (хвостовому) концу зародыша (рис.15). Клетки движутся двумя потоками по краям зародышевого диска, оба потока сталкиваются в хвостовом отделе у медианной (средней) линии, сливаются и затем направляются вперёд, к головному концу, образуя утолщенной клеточный тяж по средней лини – первичную полоску. Передняя расширенная часть первичной полоски называется первичный (гензеновский) узелок. Вдоль первичной полоски образуется углубление – первичная бороздка. Из первичной полоски клетки выселяются и вначале встраиваются в первичную энтодерму, раздвигая клетки гипобласта. Так формируется энтодерма зародыша. Далее клетки мигрируют из первичной полоски в обе стороны от неё, и встраиваются между первичной эктодермой и энтодермой, образуя средний зародышевый листок, - мезодерму. В процессе гаструляции происходит также образование мезенхимы – эмбрионального зачатка соединительной ткани. Мезенхима выселяется из всех зародышевых листком, главным образом из мезодермы.

Рис.15. Перемещение клеточных потоков
Из материала первичного узелка через первичную ямку мигрируют клетки, образующие клеточный тяж по средней линии зародыша – хорду.
Из зародышевых листков и мезенхимы происходит формирование зачатков всех тканей (гистогенез) и органов (органогенез) развивающегося организма. Вначале закладываются осевые зачатки органов, которые располагаются по оси симметрии тела.
Комплекс осевых органов включает:
нервную трубку
хорду
параксиальную мезодерму
кишечную трубку.
Нервная трубка формируется из первичной эктодермы под влиянием сигнальных молекул, вырабатываемых клетками хорды, таким образом хорда является первичным индуктором нейроэктодермы. Процесс образования нервной трубки называется нейруляцией (рис.16). В зоне образования нервной трубки в эктодерме вначале появляется утолщение – нервная пластинка. В нервной пластинке образуется углубление – нервный желобок, а по краям желобка утолщения – нервные складки. При смыкании нервных складок формируются нервная трубка и нервные валики (нервные гребни).
Из нервной трубки образуется центральная нервная система. Клетки нервного гребня активно мигрируют и дают начало периферической нервной системы, и некоторым другим структурам (мозговое вещество надпочечников, эндокриноциты диффузной эндокринной системы, клетки-меланоциты).
Остальная зародышевая эктодерма (эпидермальная эктодерма) является источником развития эпителия кожного покрова и его производных.
В мезодерме дифференцируются следующие части (рис.17):
сомиты (параксиальная мезодерма), которые располагаются справа и слева от нервной трубки и хорды;
нефротомы (сегментные ножки, промежуточная мезодерма), лежащие латеральнее сомитов;
спланхнотомы (латеральная мезодерма), наиболее латерально расположенные участки мезодермы.
Сомиты и нефротомы представляют собой сегментированную часть мезодермы.
Сомиты далее дифференцируются на:
дерматом, из которого формируется соединительнотканная часть кожи – дерма;
миотом, из которого развивается скелетная мышечная ткань;
склеротом – источник развития тканей скелета.
Нефротом служит источником для развития мочевыделительной и половой систем.
Спланхнотомы не сегментируются и расслаиваеюся на два листка – париетальный листок, прилежащий к эктодерме (соматическая мезодерма), и висцеральный листок, прилежащий к энтодерме (спланхническая мезодерма). Между париетальным и висцеральным листками формируется целомическая полость. Из листков спланхномезодермы происходит образование эпителия серозных оболочек – брюшины, плевры, перикарда.
Из мезенхимы развиваются:
кровь и лимфа;
сосуды;
кроветворные органы;
соединительная ткань;
гладкая мышечная ткань.
Из энтодермы образуется эпителий кишечной трубки.
Внезародышевые (провизорные) органы обеспечивают эмбриону условия для нормального развития и жизнедеятельности. Они функционируют в течение всего эмбриогенеза и исчезают по окончании эмбрионального развития, когда зародыш приобретает способность к самостоятельному существованию. Внезародышевые органы формируются из внезародышевых частей
Внезародышевые органы у птиц:
амнион;
серозная оболочка;
желточный мешок;
аллантоис.
Зародыш птиц первое время распластан из поверхности желтка, но в дальнейшем формируется туловищная складка. При этом все четыре листка (эктодерма, энтодерма, париетальный и висцеральный листки мезодермы) подгибаются под зародыш.
Амниотические складки, образованные из внезародышевых частей эктодермы и париетального листка мезодермы, приподнимаются сначала над головным концом, а затем над всем телом зародыша (рис.18). Эктодерма одной амниотической складки срастается с эктодермой другой. Париетальные листки мезодермы обеих складок также срастаются в единый слой. При этом формируются заполненный жидкостью пузырь – амнион, стенка которого образована амниотической оболочкой, состоящей из эктодермы, выстилающей его изнутри, и париетального листка мезодермы, лежащего снаружи. Эктодермальный амниотический эпителий секретирует амниотическую жидкость, обеспечивая водную среду для развития зародыша и защищая его от механических повреждений.
Желточный мешок образуется из внезародышевых частей энтодермы и висцерального листка мезодермы, которые постепенно обрастают поверхность желтка. Желточный мешок отделяется от первичной кишки в процессе формирования туловищной складки. Желток переводится в растворимую форму под действием ферментов, которые продуцируются энтодермальными клетками (желточной энтодермы), и используется зародышем для питания.
Серозная оболочка образована внезародышевыми частями париетального листка мезодермы (изнутри) и эктодермы (обращена наружу). Серозная оболочка обрастает весь зародыш и выполняет функцию дыхания (газообмена).
Аллантоис возникает в виде полого выроста вентральной стенки задней части кишечной трубки и состоит из энтодермы зачатка задней кишки и покрывающего его снаружи висцерального листка мезодермы с сосудами. Аллантоис разрастается, приближается к серозной оболочке и снабжает её сосудами. Функции аллантоиса:
аллантоис представляет собой мочевой мешок зародыша, где собираются продукты метаболизма;
участие в газообмене.
ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ ЧЕЛОВЕКА
Пренатальный период развития начинается с момента оплодотворения и образования зиготы и продолжается 280 дней.
Эмбриогенез подразделяется на:
начальный период (первая неделя)
эмбриональный период (со 2-й по 9 нед.)
плодный период (с 8-9 нед. до рождения).
Тип яйцеклетки человека – вторично олиголецитальная, изолецитальная.
Тип дробления зиготы, образованной изолецитальной яйцеклеткой – полное (голобластическое), то есть зигота полностью разделяется на два бластомера. Дальнейшие дробления – неравномерные, поскольку образуются 2 типа бластомеров: 1 тип - более мелкие и светлые, будущие клетки трофобласта, и 2 тип – более крупные и тёмные, будущие клетки эмбриобласта. Дробление происходит асинхронно и относительно медленно. Плоскость первого дробления проходит через редукционные тельца.
Более мелкие бластомеры в последующем образуют трофобласт, более крупные – эмбриобласт (внутреннюю клеточную массу).
На ранних стадиях (1-4 сутки после оплодотворения) бластула называется морулой (морула = «ягода»). Морула образована небольшим количеством бластомеров (до 32 бластомеров), и не имеет полости. Сначала бластомеры расположены рыхло, с довольно большими межклеточными пространствами (стадия 8 бластомеров). Затем происходит компактизация, и клетки сближаются. Мелкие бластомеры перемещаются на периферию бластулы, между ними устанавливаются плотные контакты (изолирующие) и образуют первую внезародышевую оболочку – трофобласт. Крупные бластомеры оказываются внутри, между ними устанавливаются коммуникационные контакты (щелевые, нексусы); эти клетки образуют эмбриобласт.
Дробление происходит в ходе перемещения зародыша (на этой стадии его называют преэмбрион или концептус) по маточной трубе. На 4-е сутки концептус попадает в полость матки.
С появлением полости (к 5-му дню эмбрионального развития) бластула называется бластоцистой. Эмбриобласт расположен эксцентрично, у полюса трофобласта.
Бластоциста остаётся свободной в полости матки до 6-7 дня э.р., затем погружается в слизистый секрет маточных желез на поверхности эндометрия (эндометрий – слизистая оболочка матки).
Имплантация – прикрепление и внедрение зародыша в слизистую оболочку матки начинается на 6-7-й день после оплодотворения.
Под действием маточного секрета, который выделяют клетки эпителия слизистой оболочки и маточных желез, прозрачная оболочка, окружающая бластоцисту, растворяется, и бластоциста прикрепляется к эндометрию. Эта стадия называется адгезией.
Как только трофобласт входит в контакт с эпителием эндометрия, клетки трофобласта начинают активно делиться, дочерние клетки сливаются, образуя единую цитоплазматическую массу, содержащую множество ядер, и не имеющую клеточных границ – симпластотрофобласт (синоним - синцитиотрофобласт). Таким образом, с началом имплантации трофобласт дифференцируется на цито- и симпластотрофобласт, который быстро разрастается, выделяет лизосомальные ферменты и разрушает ткани эндометрия матки.
Процесс проникновения через эпителий эндометрия и полное погружение в эндометрий - эта стадия называется инвазией - начинается с 7-го дня эмбриогенеза. Инвазия происходит благодаря активности симпластотрофобласта. Симпластотрофобласт выделяет литические ферменты, которые последовательно разрушают эпителий эндометрия матки, соединительную ткань собственной пластинки и, наконец, стенки кровеносных сосудов. Кровь из разрушенных сосудов изливается, образуя лакуны.
На 9-10-й день эмбрион оказывается полностью погруженным в эндометрий. Эпителий слизистой оболочки обрастает зародыш сверху, закрывая рану, образованную при имплантации.
Симпластотрофобласт адсорбирует продукты распада (лизиса) тканей матери. Продукты лизиса используются зародышем для роста. Такой тип питания зародыша называется гистиотрофным. Гистиотрофный тип питания характерен для первых двух недель жизни эмбриона.
Кроме того, симпластотрофобласт является местом образования гормонов, включая хорионический гонадотропин (гормон, стимулирующий образование прогестерона жёлтым телом). Этот гормон можно определить в крови или в моче женщин, начиная с 10-го дня беременности, что является основой теста на беременность.
Таким образом, роль симпластотрофобласта заключается в том, что:
благодаря действию лизосомальных ферментов он обеспечивает инвазию зародыша в стенку матки;
обеспечивает питание, необходимое для роста зародыша в первые две недели жизни, абсорбируя продукты распада тканей матки;
выполняет эндокринную функцию.
ВТОРАЯ НЕДЕЛЯ ЖИЗНИ ЭМБРИОНА
1-я фаза гаструляции – деламинация – происходит на 7-ой день жизни эмбриона. При этом эмбриобласт расщепляется на эпибласт и гипобласт. Гипобласт в будущем даст начало внезародышевой (желточной) энтодерме.
После деламинации края эпибласта, изгибаясь, срастаются с образованием небольшой полости, формируя амниотический пузырёк. Клетки гипобласта делятся, перемещаются и смыкаются, образуя стенку желточного пузырька. Таким образом, на этой стадии развития различают двухслойный зародышевый диск, при этом эпибласт образует дно амниотического пузырька, а гипобласт – крышу желточного пузырька.
Из эпибласта выселяются клетки, дающие начало внезародышевой мезодерме. Внезародышевая мезодерма образует губчатую структуру, образованную тяжами клеток. Постепенно внезародышевая мезодерма обрастает стенки амниотического и желточного пузырька, формируя амниотическую оболочку (изнутри выстлана внезародышевой эктодермой, снаружи – внезародышевой мезодермой) и желточный мешок (изнутри выстлан внезародышевой энтодермой, снаружи – внезародышевой мезодермой).
При подрастании мезодермы к трофобласту формируется хориональная оболочка (внезародышевая мезодерма + цито- и симпластотрофобласт) и небольшие полости сливаются в единую полость хориона (синоним - внезародышевый целом).
Зародыш связан с формирующимся хорионом через амниотическую ножку, образованную внезародышевой мезодермой.
Внезародышевая мезодерма врастает в выросты трофобласта, и первичные ворсинки хориона становятся вторичными (на 12-13-е сутки).
Вторую неделю эмбриогенеза часто называют периодом «двоек»:
образуются два слоя – эпибласт и гипобласт, которые составляют зародышевый диск;
развиваются два пузырька – амниотический и желточный;
дифференцируются два слоя трофобласта – цито- и симпластотрофобласт.
Таким образом, в конце первой недели – начале второй недели пренатального развития происходят такие очень важные и совпадающие по времени их протекания события (процессы), как:
имплантация;
первая фаза гаструляции;
появление внезародышевой мезодермы и развитие внезародышевых органов: амниона, желточного мешка, хориона.
В связи с этим, конец первой недели жизни эмбриона относится к критическим (определяющим) периодом развития.
ТРЕТЬЯ НЕДЕЛЯ ЭМБРИОНА
На 14-15–й день жизни эмбриона происходи вторая стадия гаструляции, которая происходит по механизму иммиграции.
Клетки, иммигрирующие в зоне гензеновского узелка, образуют хордомезодермальный отросток, первоначально растущий под эпибластом по направлению к головному концу по срединной линии зародыша. Клетки этого отростка дифференцируются в головную мезодерму и хорду. В период регрессии первичная полоска укорачивается, что приводит к перемещению первичного узелка в каудальном (хвостовом) направлении и, следовательно, росту хорды в этом направлении.
Роль хорды в процессе развития:
хорда - ось будущего позвоночника
хорда - первичный индуктор и организатор: под влиянием сигнальных молекул клеток хорды происходит дифференцировка первичной эктодермы в нейроэктодерму и образование нервной пластинки и нервной трубки.
nucleus pulposus
Формирование комплекса осевых зачатков, нейруляция и дифференцировка зародышевых листков при развитии млекопитающих происходит аналогично тому, как это происходит при развитии птиц.
В ходе образования нервной трубки после смыкания нервных валиков образуется нервный гребень (нервный крест), дающий начало целому ряду структур и клеток, в частности периферической нервной системе, мозговому веществу надпочечников и др. (перечислены в таблице ).
Под индукционным влиянием хорды и нервной трубки сомит дифференцируется на дерматом, миотом и склеротом (вентро-медиальная часть сомита). Из миотома (внутренний слой клеток дорсолатеральной части сомита) развивается скелетная мускулатура. Дерматом наружный слой клеток дорсолатеральной части сомита) даёт начало соединительной ткани кожи – дерме. Клетки склеротома направленно мигрируют в места окончательной локализации и дифференцируются в клетки хрящевой и костной ткани, формируя позвонки, рёбра, лопатки – осевой скелет.
Мезенхима – эмбриональная соединительная ткань, образована Клетки мезенхимы выселяются из всех зародышевых листков, наиболее активно из висцерального листка спланхнотома. Из мезенхимы образуются нескольких типов тканей и структур:
кровь;
все типы соединительной ткани;
гладкая мышечная ткань,
кровеносные сосуды и эндокард сердца;
микроглия центральной нервной системы.
Первые кровеносные сосуды и очаги (островки) кроветворения обнаруживаются в стенке желточного мешка.
Кровеносные сосуды врастают в хориональные ворсинки, которые с появлением сосудов становятся третичными (такие ворсинки появляются на 3-й неделе эмбрионального развития).
С 20-го дня эмбриогенеза начинается обособление тела зародыша от внезародышевых оболочек - образуется туловищная складка.
ВНЕЗАРОДЫШЕВЫЕ ОРГАНЫ В ЭМБРИОГЕНЕЗЕ ЧЕЛОВЕКА:
амнион
желточный мешок
аллантоис
хорион
плацента
Амнион
Амнион образует замкнутую полость вокруг зародыша.
Функции амниона:
создание водной среды определенного химического состава и давления для свободного развития эмбриона и плода;
защита от механических и гравитационных стрессов;
предотвращает слипание плода с окружающими тканями.
Стенка амниона образована амниотическим эпителием (внезародышевой эктодермой, которая развивается из эпибласта) изнутри и внезародышевой мезодермой снаружи. Постепенно полость амниона разрастается. К 7-й неделе развития амниотическая мезодерма входит в контакт с мезодермой хориона (амнио-хориональная оболочка). Кроме того, амниотический эпителий обрастает амниотическую ножку. Амнион функционирует до момента рождения (плодный пузырь). К концу беременности заполнен 1-1,5 литрами амниотической жидкости (околоплодные воды).
Желточный мешок
Стенка желточного мешка изнутри образована внезародышевой энтодермой. Её формируют интенсивно делящиеся клетки гипобласта, которые перемещаются по внутренней поверхности трофобласта. Снаружи внезародышевая энтодерма обрастает внезародышевой мезодермой.
У человека желточный мешок функционирует только на ранних стадиях развития (7-8 недель).
Функции желточного мешка:
стенка желточного мешка - место первых очагов кроветворения и образования кровеносных сосудов (на 3-й неделе развития);
стенка желточного мешка - место появления первичных половых клеток (гонобластов).
после 7-8 недели подвергается регрессии, остаётся в виде тяжа клеток в пупочном канатике, направляющего кровеносные сосуды к плаценте.
Аллантоис
Аллантоис развивается на 16-17-е сутки в виде небольшого выроста задней стенки желточного мешка, поэтому имеет те же оболочки, что и желточный мешок: внезародышевая энтодерма изнутри и внезародышевая мезодерма снаружи. Аллантоис врастает в амниотическую ножку, в его стенке развиваются пупочные кровеносные сосуды, которые он подводит к хориону. Таким образом, аллантоис выполняет ту же функцию, что и регрессирующий желточный мешок – они играют роль проводников и направляют рост сосудов плода к плаценте. На втором месяце эмбрионального развития аллантоис редуцируется и вместе с остатками желточного мешка образует клеточный тяж в составе пупочного канатика. Кроме того, аллантоис участвует в развитии мочевого пузыря.
Хорион
В формировании хориона различают три периода: предворсинчатый (7-8-е сутки), период образования ворсинок (до 50-х суток), период котиледонов (с 50 по 90-е сутки).
Зрелый хорион образован хориональной пластиной (внезародышевая мезодерма) и выростами пластины – ветвящимися третичными ворсинками, покрытыми трофобластом. Часть хориона, разрушающая стенку матки и участвующая в образовании плаценты, формирует сложноразветвленные ворсинки и носит название ворсинчатый хорион. Остальную поверхность составляет гладкий хорион. Самые крупные ворсинки, отходящие от хориональной пластины, носят название стволовых ворсинок. Стволовые ворсинки обильно ветвятся, самые мелкие веточки носят названия терминальных ворсинок. Кровеносные сосуды в терминальных ворсинках представлены капиллярами плода. Все ворсинки покрыты снаружи трофобластом. Ворсинки, внедряющиеся в базальную пластинку эндометрия, называются якорными ворсинками. Обычно стволовые ворсинки являются якорными.
Плацента
Плацентация – период эмбриогенеза, на протяжении которого происходит развитие плаценты, один из критических периодов эмбриогенеза, соответствует 3-6 неделям беременности.
Плацента – единственный орган, состоящий из клеток двух генетически различных организмов: плодной части (хорион с ворсинками) и материнской части.
Плодная часть плаценты – ворсинчатый хорион (хориональная пластина с ворсинками). Ворсинки хориона погружены в лакуны, заполненные кровью матери.
Материнская часть плаценты представлена измененной слизистой оболочки матки, которая называется эндометрием. Эндометрий, кроме самого глубокого базального слоя, отторгается при рождении ребёнка, поэтому эти структуры получили название децидуальной (отпадающей) оболочки. В зависимости от расположения относительно места имплантации различают:
Decidua parietalis (пристеночная) – эндометрий, выстилающий полость матки за исключением участка имплантации;
Decidua capsularis (сумочная) – часть эндометрия, которая окружает развивающийся эмбрион, образуя поверх него капсулу, и отделяет зародыш от полости матки (до 16-й нед.)
Decidua basalis (основная), материнская часть плаценты, та часть эндометрия, которая находится между плодом и базальным слоем эндометрия.
Итак, материнская часть плацента представлена:
базальной пластинкой (decidua basalis) эндометрия;
лакунами, заполненными материнской кровью.
Кровь матери и ребенка не смешивается. Их разделяет гематоплацентарный барьер. Компоненты гематоплацентарного барьера, разделяющего кровь матери и кровь плода (рис. ):
эндотелий капилляра плода;
базальная мембрана в стенке капилляров плода;
соединительная ткань ворсинок (с клетками-макрофагами);
базальная мембрана трофобласта;
цитоторофобласт;
симпластотрофобласт.
Ворсинки, обращенные к decidua basalis, распределены неравномерно, группами – котиледонами. Котиледон – структурно-функциональная единица сформированной плаценты. Котиледон образован стволовой ворсинкой и её разветвлениями. Котиледоны частично разделены соединительнотканными септами (плацентарными перегородками), отходящими от базальной пластинки (рис. )
К концу беременности плацента имеет форму диска.
Связь между циркуляциями крови плода и матери осуществляется через пупочный канатик.
Функции плаценты:
трофическая - из организма матери к плоду поступают самые разнообразные питательные вещества, электролиты, витамины, некоторые гормоны;
дыхательная - транспорт кислорода в кровь плода и перенос углекислого газа в кровь матери;
выделительная - из крови плода в кровь матери поступают продукты метаболизма и выделяются через почки матери;
защитная - препятствует развитию иммунного конфликта, проникновению лекарственных веществ – барьер не абсолютный;
эндокринная – здесь происходит синтез ряда гормонов и других биологически активных веществ (хорионический гонадотропин, прогестерон, фактор роста фибробластов, трансферрин, пролактин, релаксин и другие), имеющих важное значение для нормального течения беременности и развития плода.
Таблица 2
|
Обмен через плацентарный барьер | |
|
Из материнской крови в кровь плода (снабжение) |
Из крови плода в материнскую кровь (выделение) |
|
|
Гематоплацентарный барьер не является абсолютным и проницаем для ряда веществ и возбудителей болезней (таблица 3)
Таблица 3
|
Возбудители болезней |
Вирусы (ВИЧ, цитомегаловирус, краснухи, оспы, ветряной оспы, кори, полиомиелита) Бактерии (туберкулез, Treponema) Простейшие (Toxoplasma) |
|
Наркотические и лекарственные средства |
Кокаин, алкоголь, кофеин, никотин, зоокумарин, дифенин тетрациклин противораковые препараты, анестетики, успокаивающие, анальгетики |
|
Антитела |
Антитела к резус-фактору |
ТИПЫ ПЛАЦЕНТЫ млекопитающих (рис.):
Эпителиохориальная (пример – плацента свиньи): ворсинки хориона проникают в углубления слизистой оболочки (маточные железы), не разрушая их;
Десмохориальная плацента (копытные): ворсинки хориона контактируют с соединительной тканью эндометрия матери;
Эндотелио- (вазо-)хориальная (хищники) плацента: ворсинки хориона контактируют со стенкой сосудов эндометрия матери, не разрушая стенки.
Гемохориальная плацента (плацента человека): ворсинки хориона разрушают эпителий, соединительную ткань, стенки сосудов матери и омываются кровью, заполняющие лакуны.
Таблица 4
Дифференцировка зародышевых листков
|
Зародышевый листок |
Производные во взрослом организме | |||
|
ЭКТОДЕРМА |
Покровная эктодерма |
эпидермис кожи и его производные (волосы, ногти, потовые и сальные железы); молочные железы; эпителиальная выстилка органов полости рта; энамелобласты (секреция органического матрикса эмали зуба); слюнные железы; хрусталик глаза; аденогипофиз; выстилка наружного, среднего и внутреннего уха; эпителий анального канала
| ||
|
Нейроэктодерма |
Нервная трубка |
центральная нервная система (все нейроны и глия); сетчатка; нейрогипофиз; эпифиз | ||
|
Нервный гребень |
периферическая нервная система и другие структуры (таблица | |||
|
МЕЗОДЕРМА |
Параксиальная (СОМИТЫ) |
Дерматом |
Дерма кожи | |
|
Миотом |
Скелетная мускулатура туловища, конечностей, головы и шеи; глазные мышцы; мышечное тело языка. | |||
|
Склеротом |
Позвонки, рёбра, лопатки | |||
|
Промежуточная (НЕФРОТОМ) |
Эпителий почки и органов репродуктивной системы
| |||
|
Латеральная (СПЛАНХНОТОМ) |
Выстилка серозных оболочек полостей тела (брюшина, перикард, плевра) Миоэпикардиальная пластика (миокард, эпикард) Корковое вещество надпочечников | |||
|
ЭНТОДЕРМА |
эпителиальная выстилка желудочно-кишечного тракта, печень, поджелудочная железа, желчный пузырь; эпителиальная выстилка дыхательной системы (трахея, бронхи, лёгкие), строма тимуса, эндокриноциты щитовидной и паращитовидной железы, эпителий мочевого пузыря, женской уретры и большей части мужской уретры. | |||
|
МЕЗЕНХИМА |
Кровь, лимфа, соединительная ткань, гладкая мышечная ткань, кровеносные сосуды, эндокард | |||
Таблица 5
|
ПРОИЗВОДНЫЕ НЕРВНОГО ГРЕБНЯ
|
|
Чувствительные нейроны спинальных узлов и узлов черепных нервов Нейроны симпатической и парасимпатической нервной системы Нейролеммоциты (шванновские клетки) и глиоциты ганглия Мягкая и паутинная оболочки мозга; Меланоциты Клетки каротидного тельца Парафолликулярные клетки щитовидной железы Клетки диффузной эндокринной системы желудочно-кишечного и дыхательного трактов (APUD-серия) Хромаффиноциты мозгового вещества надпочечников Хрящи, кости, мышцы и соединительные ткани лица (эктомезенхима) Верхнечелюстной и челюстной отростки; Подъязычные дуги и 3-я глоточная дуга; Одонтобласты → дентин зуба Задний эпителий роговицы Расширяющая и суживающая мышцы зрачка Ресничная мышца
|
Embryology is a science about the embryonic development of the organism.
The embryonic development, or embryogenesis, is a period from the moment of fertilization to the birth (in live-bearing animals).
The process of embryogenesis may be divided into several stages:
fertilization is a series of events leading to fusion of male and female gametes with formation of unicellular new organism, the zygote;
cleavage, which consists of a series of rapid mitotic divisions which result in in formation of multicellular embryo, the blastula;
gastrulation is a complex series of morphogenetic movements of cell groups leading to the establishment of the primary germ layers that giving rise to the embryo, composing of three germ layers, the gastrula;
neurulation and notogenesis, the formation of the nerve tube and other axial primordia;
histogenesis and organogenesis, each of the three germ layers gives rise to a number of specific tissues and organs;
systemogenesis, the organ systems formation..
Gametogenesis, the production of specialized male and female germ cells or gametes precedes fertilization.
GAMETOGENESIS. REPRODUCTIVE CELLS
Gametogenesis is the production of specialized germ cells, or gametes. During the gametes’ maturation the number of chromosomes is reduced by half to the haploid number.
Primordial germ cells, lso known as PGCs, precursor germ cells or gonocytes or gonoblasts, in human embryos appear in the wall of the yolk sac at the end of the third week of the prenatal life. These cells are indifferent: they are not divided onto the male and the female cells. They migrate by ameboid movement from the yolk sac toward through tthe developing gonads (the paired gonadal ridges on the medial surface of the mesonephros), divide repeatedly on their migratory route, and differentiate there into spermatogonia or oogonia (in humans, sexual differentiation starts approximately 6 weeks post conception). Germ cells produce gametes and are the only cells that can undergo meiosis as well as mitosis.
SPERMATOGENESIS
Male reproductive cells, the spermatozoa, are produced in seminiferous tubules of the testis beginning with the puberty age and continue throughout adult life. The process of differentiation of the male germ cells is called spermatogenesis and can be divided into 4 phases (Fig.2):
proliferation phase
growth phase
maturation phase
formation, or spermiogenesis
The phase of proliferation. During this phase spermatogonia divide by mitoses, producing successive generations of cells that finally give rise to primary spermatocytes. Spermatogonia consist of two types: type A spermatogonia, and type B spermatogonia (fig.1).
Type A dark spermatogonia are true stem cells, steady to harmful factors action, rarely dividing cell. At sexual maturity, this cell undergoes a series of mitoses, and the newly formed cells can follow one of 2 paths: they can continue, as undifferentiated stem cells (type A pale spermatogonia), or they can differentiate during progressive mitotic cycles to become type B spermatogonia.
Type B spermatogonia are committed cells, they prepare to the first meiotic division, enter the phase of growth and give rise to primary spermatocytes.
The phase of growth. During this phase primary spermatocytes grow (their size 4 times more as spermatogonia), and enter a prolonged prophase of the first meiotic division (lasts about 22 days), followed by rapid completion of meiosis I and formation of secondary spermatocytes;
The phase of maturation. Secondary spermatocytes immediately enter the second meiotic division, to form spermatids, which contain the haploid number of chromosomes;
The phase of formation, or spermiogenesis. During this phase the spermatids go through an elaborate process of cytodifferentiation, producing small, highly motile spermatozoa.
Spermiogenesis is a complex process of differentiation that includes (Fig.3):
formation of the acrosome;
condensation and elongation of the nucleus;
flagellum’ development;
the loss of much of the cytoplasm.
The
acrosome
is a special organelle, originated from the Golgi complex. It is a
membrane-limited vesicle (the acrosomal cap), which cover the
anterior half of the condensing nucleus. The acrosome contains
several hydrolytic enzymes, such as hyaluronidase, penetrase, acrosin
(trypsin-like e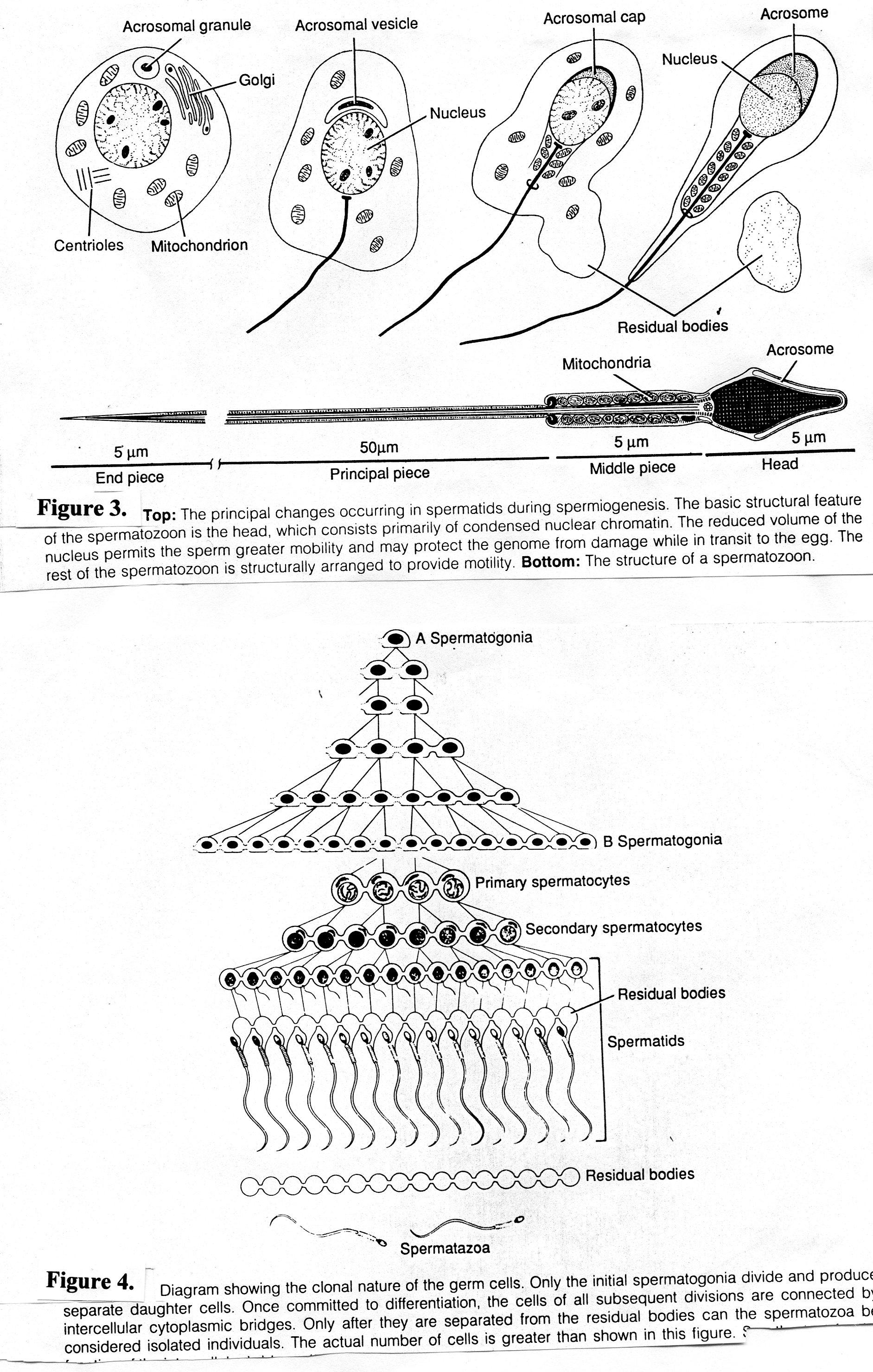 nzyme),
acid phosphatase, and proteases. The acrosome thus serves as a
specialized type of lysosomes. These enzymes are known to dissociate
follicular cells ofthe
corona radiata
and to digest the
zona pellucida,
structures that surround recently ovulated eggs.
nzyme),
acid phosphatase, and proteases. The acrosome thus serves as a
specialized type of lysosomes. These enzymes are known to dissociate
follicular cells ofthe
corona radiata
and to digest the
zona pellucida,
structures that surround recently ovulated eggs.
Fig.3. Top: The principal changes occurring in spermatids during spermiogenesis.
Bottom: The structure of a spermatozoon.
During spermiogenesis the centrioles of the spermatid migrate to a position near the cell surface and opposite the location of the forming acrosome. One of the centrioles (distal) gives rise to microtubules of the axoneme of the flagellum. The axoneme is surrounded by dense skeleton (coarse fibrils, rib-like fibrils and columns). The mitochondria aggregate in a helical manner around the proximal part of the flagellum axoneme, forming a thickened region known as the middle piece, the region where the movements of the spermatozoa are generated.
The reduced volume of the nucleus permits the sperm greater mobility and may protect the genome from the damage while it transit to the egg cell.
Residual cytoplasm is shed and excess cytoplasm is phagocytosed by Sertoli cells. Cytoplasmic bridges break down and the spermatozoa are released into the lumen of the seminiferous tubule.
The time required for a spermatogonium to become a mature spermatozoon is approximately 64 days.
During division of the spermatogonia, the resulting cells do not separate completely but remain attached by cytoplasmic bridges forming non-cellular structure called the syncitium. The intercellular bridges provide communication between every primary and secondary spermatocytes and spermatid derived from a single spermatogonium. By permitting the interchange of information from cell to cell, these bridges play an important role in coordinating the sequence of events in spermatogenesis. When the process of spermatogenesis is completed, the sloughing of the cytoplasm and cytoplasmic bridges as residual bodies leads to a separation of the spermatids.
The mature spermatozoon has a lash-like appearance and consists of a head, a neck, and a tail with middle, principal and terminal pieces (Fig.4). The head, which, for most mammals, has a flattened, oval shape, contains the condensed nucleus and is capped by an acrosome filled with hydrolytic enzymes. The midpiece contains large helical mitochondria that generate the energy. The tail contains microtubules.
OOGENESIS
.The process of oogenesis occurs in the ovary. The oogenesis includes 3 phases (Fig.4):
prolif
growth
maturation
The phase of proliferation. Once primordial germ cells have arrived in the gonad of a genetic female, they differentiate into oogonia. Oogonia divide by mitosis only during embrional life. Beginning in the 3-rd fetal month, some oogonia enter the prophase of the 1st meiotic division and become much larger primary oocytes. In the human, this process is completed by the end of the seventh month of gestation. During this time, many primary oocytes are lost as a result of a degenerative process called atresia. So oocytes are formed during intrauterine life, and their number does not increase after birth.
The phase of growth. This phase lasts a very long time (from 12 up to 50 years) and subdivided onto small and large growth.
Small growth period. At birth all egg cells in the ovary are primary oocytes. Their total number varies from 700,000 to 2 million.
The primary oocytes become surrounded by a layer of flattened follicular cells and become known as primordial follicles. The process of encapsulation of the primary oocyte blocks the first meiotic division in the prophase. Primary oocytes do not finish their 1st meiotic division but remain in the diplotene stage until puberty.
Large growth period. With the onset of puberty a number of primordial follicles begin to grow and mature with each ovarian cycle under influence of follicle-stimulating hormone (FSH). Under normal conditions, only one of these follicles reaches full maturity, the others degenerate and become atretic. Only approximately 400,000 primary oocytes are present by the beginning of puberty, and fewer than 500 will be ovulated in the reproductive lifetime of the individual.
The phase of maturation. The first meiotic division is completed just before ovulation. The chromosomes are equally divided between the daughter cells, but one cell, the secondary oocyte, retains almost all of the cytoplasm. The other becomes the first polar body, a very small cell containing the nucleus and a minimal amount of cytoplasm. Immediately after expulsion of the first polar body the secondary oocyte starts the 2nd meiotic division, which stops in metaphase. The 2nd meiotic (maturation) division will be completed only if the oocyte is fertilized. The secondary oocyte, in turn, gives rise to a mature oocyte plus another polar body. Hence, a primary oocyte develops into one mature oocyte and three polar bodies.
Зрелая яйцеклетка млекопитающих – крупная, в сравнении со сперматозоидом, неподвижная клетка. Особенность строения яйцеклетки – отсутствие центриолей и наличие характерных трофических включений – желточных гранул. По периферии яйцеклетки, под клеточной мембраной, равномерно распределены кортикальные гранулы, содержащие ферменты (гидролазы), необходимые для формирования оболочки оплодотворения после проникновения сперматозоида.
При овуляции яйцеклетка окружена (рис.6):
прозрачной оболочкой (zona pellucida), содержащей густую сеть тонких нитей, состоящих из гликопротеинов (фракции ZP1, ZP2, ZP3);
фолликулярными клетками, образующими лучистый венец
The phase of formation in oogenesis is lacking.
Table 1
|
Comparison of spermatogenesis and oogenesis | ||
|
Similarities |
Differences | |
|
Spermatogenesis |
Oogenesis | |
|
Both processes begin with mitosis (phase of proliferation) Both processes have meiosis |
|
|
Ovary Structure
Oogenesis results in a large non-motile ovum (Fig.5). This cell is spherical and many times larger than a spermatozoon. Ovum has no centrioles, so this cell is not capable to divide itself. The cytoplasm of the ovum is packed with food reserve. Membrane-bound cortical granules accumulate in the cytoplasm, just beneath the plasma membrane. Cortical granules contain enzymes releasing a cortical reaction.
Fig.5. Human egg cell: 1) nucleus; 2) egg cell microvilli; 3) follicular cells; 4) follicular cells processes; 5) cortical granules; 6) yolk inclusions; 7) zona pellucida; 8) receptors.
In
matu re
follicles the oocyte is enveloped by thezona
pellucida,
a thick coat, composed of at least 3 different glycoproteins (ZP1,
ZP2, ZP3). ZP3
is the ligand in the zona pellucida which binds with the acrosome of
the sperm in the acrosome reaction.
re
follicles the oocyte is enveloped by thezona
pellucida,
a thick coat, composed of at least 3 different glycoproteins (ZP1,
ZP2, ZP3). ZP3
is the ligand in the zona pellucida which binds with the acrosome of
the sperm in the acrosome reaction.
The oocyte with the zona pellucida is surrounded by several layers of follicular cells. Long apical processes of follicular cells and microvilli of the oocyte penetrate the zona pellucida and make contact with one another via gap junctions. The follicular cells of the first layer around the ovum, that are, therefore, in close contact with the zona pellucida form the corona radiata, which accompanies the ovum when it leaves the ovary in the process of ovulation. The corona radiata is still present when the spermatozoon fertilizes the ovum; it is retained for some time during the passage of the ovum through the oviduct.
FERTILIZATION
Fertilization (also known as conception), the process by which male and female gametes fuse, occurs normally in the in the ampulla of the uterine tube.
During ovulation the secondary oocyte, together with surrounding the zona pellucida and corona radiata cells, is discharged from the ovary and is carried into the tube by sweeping movements of the fimbriae and by motion of cilia on the epithelial lining.
Human ejaculate contains 3x108 spermatozoa. These cells remain viable in the female reproductive tract for approximately 24 hours. Only 300-500 spermatozoa reach the site of fertilization.
Fertilization is more a chain of events than a single, isolated phenomenon and may be subdivided into 3 stages:
distant interaction;
contact interaction;
penetration of the sperm cell into the oocyte and egg activation;
fusion of male and female pronuclei, and formation of a zygote.
The process of fertilization lasts 12-24hours.
The distant interaction is based on phenomenon of chemotaxis: it was demonstrated that the surface membrane of human sperms possess receptors for specific chemoattractant substances (such as formyl Met-Leu-Phe) which have the ability to induce migration of sperm. Contraction of the musculature of the uterine tube promote this ascent.
Freshly ejaculated sperm are unable to fertilize. Sperm cells must undergo capacitation.
Capacitation occurring in the female reproductive tract, is a complex process, which renders the sperm cell capable for specific interaction with the oocyte. This process takes place under influence of mucous secretion of uterine tube epithelial cells, pH in the female reproductive tract etc. This final step in the maturation of spermatozoa is biochemical event. It associated with removal of adherent seminal plasma glycoproteins, reorganization of plasma membrane lipids and proteins, rendering it more fusigenic. The result is a more fluid membrane with an increased permeability to Ca2+. An influx of Ca2+ produces increased intracellular cAMP levels and thus, an increase in motility. Hyperactivation is also part of capacitation and is the result of the increased Ca2+ levels.
Capacitation lasts approximately 7 hours. Spermatozoa used for in vitro fertilization are artificially capacitated.
The contact interaction includes:
attachment and binding. The sperm binds to the zona pellucida of the egg. A special complementary molecule on the surface of the sperm head then binds to ZP3 glycoproteins in the zona pellucida. Binding of sperm to the zona pellucida is a receptor-ligand interaction with a high degree of species specificity: only sperm of same species may enter.
the acrosomal reaction. This binding triggers the acrosomal reaction: the acrosome vesicle membrane fuses with the sperm plasma membrane at multiple sites, releasing acrosomal enzymes, such as hyaluronidase and acrosin to degrade the zona pellucida and, in that way, facilitate penetration through it. After the acrosome reaction, it is believed that the sperm remains bound to the zona pellucida through exposed ZP2 receptors.
Penetration of spermatozoon and egg activation and the cortical reaction. Upon binding of a sperm cell, the egg rapidly undergoes a number of metabolic and physical changes that collectively are called egg activation. Prominent effects include a rise in the intracellular concentration of calcium, completion of the second meiotic division and the cortical reaction. Once the sperm cells find their way past the zona pellucida (owing to 2 factors: motility and zona-digesting enzymes), the cortical reaction occurs: cortical granules inside the ovum fuse with the plasma membrane of the cell, causing enzymes (including several proteases) inside these granules to be expelled to the zona pellucida. This in turn causes the glycoproteins in the zona pellucida to cross-link with each other, making the whole matrix hard and impermeable to sperm (sperm receptors are destroyed also). So, the zona pellucida transforms into the fertilization membrane (or the zona reaction) around a fluid-filled perivitelline space. This membrane serves as a barrier and prevents completely further penetration of other sperms (block to polyspermy).
Fusion and formation of a zygote
Both the oocyte and the sperm go through transformations, as a reaction to the fusion of cell membranes, preparing for the fusion of their genetic material.
Entry of the sperm triggers completion of the oocyte second meiotic division. This results in a mature ovum. The nucleus of the oocyte is called a female pronucleus in this process, to distinguish it from the nuclei that are the result of fertilization.
The head of engulfed sperm swells, forming a male pronucleus. The sperm’s tail and mitochondria degenerate with the formation of the male pronucleus. This is why all mitochondria in humans are of maternal origin.
This process ultimately leads to the formation of a diploid cell called a zygote. The zygote is the fertilized ovum, and is the first form of a new human being.
The male pronucleus, its centrioles and female pronucleus migrate towards the center of the oocyte, rapidly replicating their DNA as they do so to prepare the new human for its first mitotic (cleavage) division.
The male and female pronuclei don’t fuse, although their genetic material do so. Instead, their membranes dissolve, leaving no barriers between the male and female chromosomes. During this dissolution, a mitotic spindle forms around them to catch the chromosomes before they get lost in the egg cytoplasm. The first cleavage division follows, producing two identical daughter cells.
BIOLOGICAL SIGNIFICANCE of FERTILIZATION
the penetration by the sperm cell reconstitutes the diploid number of chromosomes;
the zygote contains a new combination of chromosomes and genetic material that is different from those of the parents;
the embryo sex is determined at fertilization by kind of the sperm that fertilizes the ovum (X-bearing sperm or Y-bearing sperm);
the penetration by the sperm cell serves as a stimulus for the ovum to complete the second meiotic division and cast off the second polar body;
fertilization initiates a series of rapid mitotic divisions called the zygote cleavage
Comparative Vertebrate Embryology
CLASSIFICATION of EGG CELLS
Morphological characteristics of egg cells of different representatives of the phylum Chordata vary. The principal modifications are associated with varying amounts of yolk within ovum. Yolk decelerates or even prevents division of the ovum cytoplasm. Thus, amounts of yolk as well as regional differences in yolk allocation are important for ways of subsequent development. So, classification of ova is based on amounts of yolk and its allocation within ovum (Tabl.2; Fig.6).
Tabl.2 TYPES of OOCYTES
|
OLIGOLECITHAL |
POLYLECITHAL | ||
|
ISOLECITHAL |
TELOLECITHAL | ||
|
PRIMARY (amphioxus) |
SECONDARY (mammals) |
MESOLECITHAL (Pisces, amphibious) |
PRONOUNCED-TELOLECITHAL (reptiles, birds) |
A) primary isolecithal egg cell; B) mesolecithal egg cell; D) pronounced telolecithal egg cell; E) secondary isolecithal egg cell:
1) Nucleus; 2) yolk (vitelline) granules; 3) follicular cells
Oligolecithal ovum is characterized by small amounts of yolk. In any oligolecithal ovum yolk is equally distributed within cytoplasm. Thus, all oligolecithal ova are isolecithal ova. A primitive vertebrate (such as Amphioxus, lancelet) leading an aquatic existence produces many small, yolk-poor eggs, which form free swimming larvae. Their ova are known as primary isolecithal ova. Through phylogenesis in eutherian mammals yolk has been secondarily reduced to the oligolecithal form, because their development occurs in a uterus. Thus, the human egg cell can be referred to oligolecithal, secondary isolecithal oocyte.
Polylecithal ovum is characterized by large amounts of yolk. Telolecithal ovum is characterized by the polarity of yolk allocation. One of the cell poles - the vegetative pole - is laden heavily with yolk, while another pole – the animal pole – is not so heavily yolked. The nucleus is placed at this animal pole. This polarity in yolk allocation may be pronounced more moderately – in mesolecithal ova, or more sharply – in pronounced-telolecithal ova. Most amphibians have large, moderately heavily yolked (mesolecithal) eggs, which form free swimming larvae (tadpoils). In adaptation to a terrestrial existence, reptiles and birds produce few, large, yolk-rich (pronounced-telolecithal) eggs, with protective membranes and a shell, and have no larval stage.
CLEAVAGE
The zygote undergoes a rapid series of mitotic divisions called cleavage.
Cleavage is a rapid succession of consists of cell mitotic divisions following DNA synthesis but without cell grown: no increase in size, only increase in cell number.. These divisions result in a collection of small adherent cells called blastomeres. They do not grow in size but divide the zygote progressively into smaller cells.
The type of cleavage depends on the type of the egg cell in animal development. The amount and distribution of yolk in each cell determines the rate of cleavage and the relative size of the blastomeres.
Cleavage may be complete (holoblastic) or incomplete depending on extent of cytoplasmic subdivision (total or partial subdivision respectively), and to be equal or unequal depending on the relative sizes of the early blastomeres.
A series of cleavage divisions result in formation of blastula. Type of blastula depends on the cleavage type (Tabl.3, Fig.12).
Tabl.3 The principal types of cleavage and blastula
|
Group |
Type of the egg cell |
Type of the cleavage |
blastula |
Blastula cavity (blastocoele) |
|
Amphioxus, Mammals |
Isolecithal, oligolecithal or microlecithal (little yolk) |
Complete (or holoblastic): initial cleavage planes extend through egg); Equal (in amphioxus), or unequal (in mammals) |
Sphere with single layer wall (celoblastula in Amphioxus; blastocyst in mammals) |
Large, central sphere |
|
Amphibians, fishes |
Mesolecithal (moderate yolk) |
Holoblastic unequal |
Sphere with layered wall (amphiblastula) |
Small sphere |
|
Reptiles Birds |
Telolecithal (heavy yolked) |
Incomplete (or meroblastic): initial cleavages planes limited to animal pole, the region opposite the yolk) |
Cell disk on surface of yolk, bilaminar - diskoblastula |
Flat space between epiblast and hypoblast |
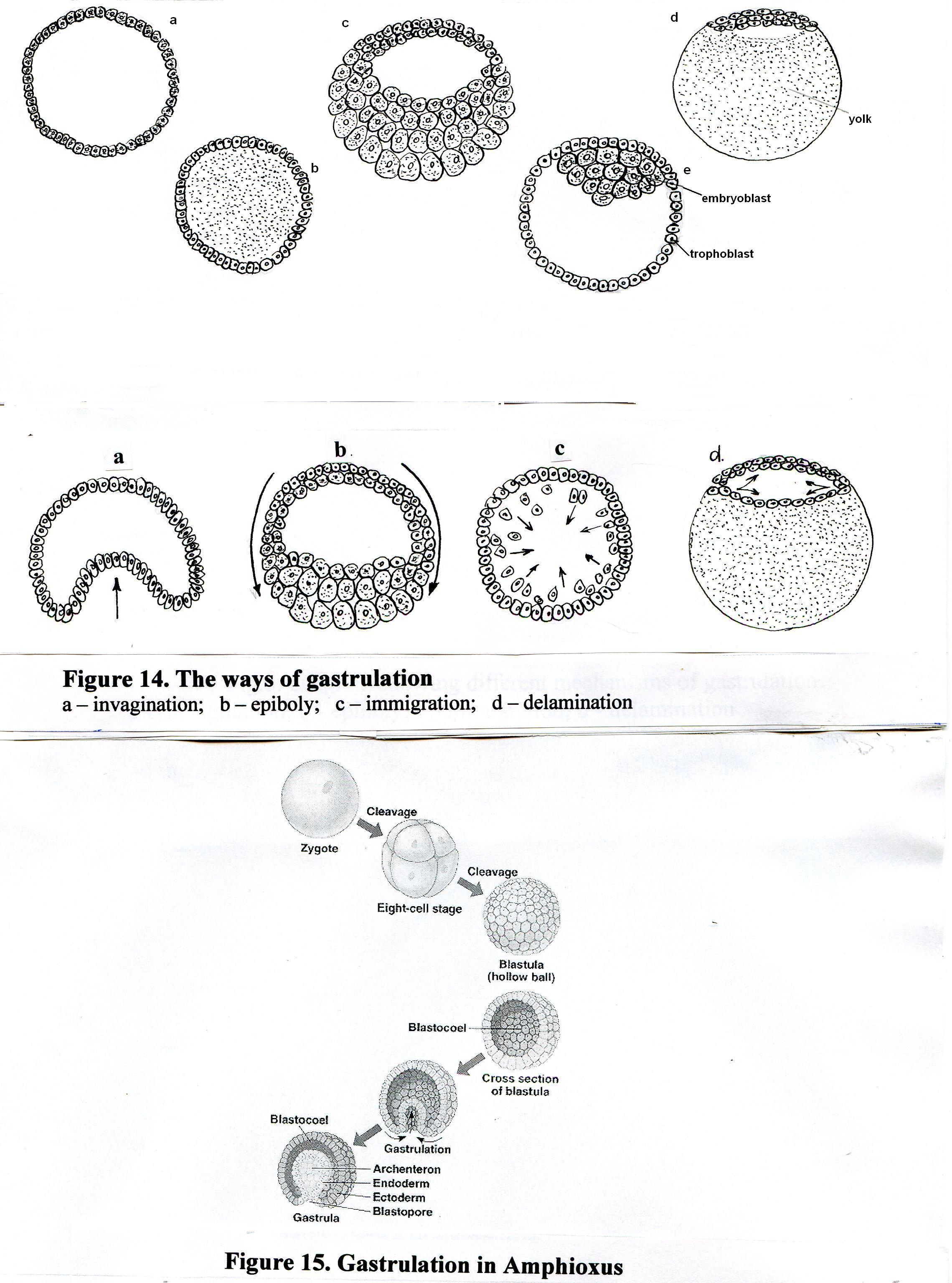
Fig.12. Blastula Types: a) celoblastula; b) periblastula (in Insects); c) amphiblastula;
d) diskoblastula; e) blastocyst.
The egg cells (and hence the zygote as well as blastula) is always asymmetric, having an animal pole and a vegetal pole. The upper hemisphere is the animal pole; the lower hemisphere is the vegetal pole. The vegetal pole draws its name from its inactivity relative to the lively animal pole. In some animal groups the vegetal pole contains large yolky cells that divide very slowly.
Complete and equal cleavage of the oligolecithal egg of Amphioxus results in a simple hollow sphere of cells enclosing a cavity – blastocoele. This sphere is termed a celoblastula.
In result of complete unequal cleavage of the mesolecithal egg of frog it is formed a blastula with an eccentric blastocoele surrounded by small blastomeres at the animal pole and by larger blastomeres at the vegetative pole. This blastula is called amphiblastula.
During incomplete and unequal cleavage of the telolecithal egg of the chick, the yolk does not divide. Cytoplasmic division are limited to the animal pole, and bilaminar disk shaped mass of blastomeres, surrounding the yolk, is produced. A cleft between the laminae is homologous to the blastocoele of lower living forms’ blastula. A subgerminal cavity between blastomeres and main yolk mass is formed shortly thereafter. It is filled with nutritive liquefying yolk. This blastula is called diskoblastula.
Complete unequal cleavage of the secondary isolecithal egg of eutherian mammal results in a compact collection of cells called a morula (20-50 cells). Morula begins to absorb uterine fluid forming a central cavity. The blastocyst, as it now known, consists of a peripheral layer of blastomeres forming the trophoblast, with a mass of cells at one aspect, the polar trophoblast, bulging into the central lumen and known as the inner cell mass, or embryoblast from which the embryo proper arises.
GASTRULATION
The next stage of embryogenesis is a process of transformation of blastula into gastrula, the embryo, consisting of 3 primary germ layers:
ectoderm (externally)
endoderm (internally) and
mesoderm (between them).
Gastrulation is rearrangement of the cells by migration and division. Gastrulation is highly variable in vertebrates so the different types will be described. Four main mechanisms of gastrulation can be distinguished in animals of the phylum Chordata (Fig.17):
invagination;
epiboly;
delamination;
immigration.
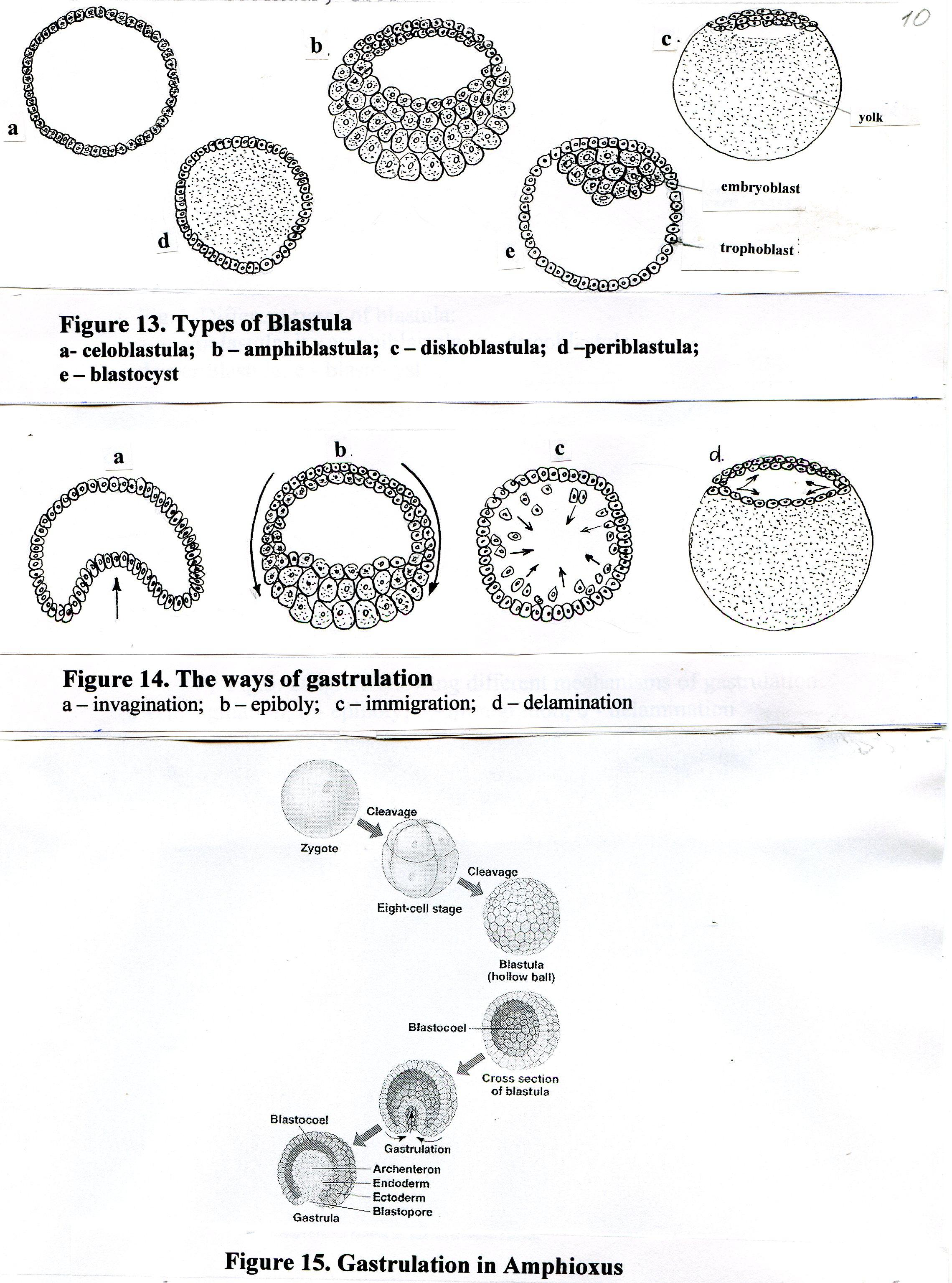
Fig.13. The ways of gastrulation: a – invagination; b – epiboly; c – immigration; d – delamination.
In forms with free swimming larvae (Anamniota) the whole blastula is transformed into the embryo, whereas in terrestrial forms (Amniota) some of the blastula forms extra-embryonic membranes.
In Amphioxus gastrulation proceeds by invagination: part of the blastula wall sinks below the surface so that a double-walled cup – the gastrula – is formed. The central cavity of the cup is a primitive gut – gastrocele, or archenterons, and the opening to the exterior is termed blastopore. Further rearrangement of the gastrula cells gives rise to a third layer within the double wall (Fig.14).
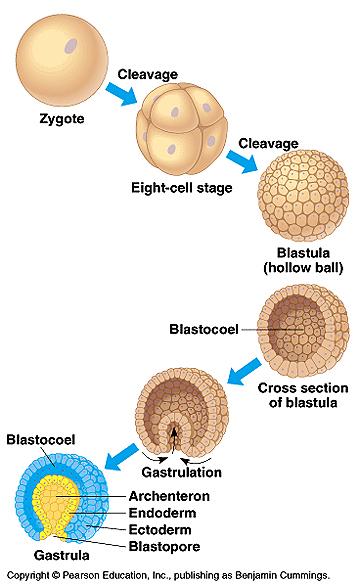 Fig.14.
Gastrulation in Amphioxus.
Fig.14.
Gastrulation in Amphioxus.
In Amphibian gastrulation occurs through combination of invagination and epiboly (Fig.15).
Fig.15. Gastrulation in Frogs
In reptiles, birds and mammals gastrulation proceeds by combination of delamination and immigration.
For practical reasons most of experiments on the mechanisms of vertebrate development have been carried out on amphibia. At the end of cleavage the amphibian blastula consists of a hollow sphere of cells enclosing a cavity – the blastocele. At the animal pole the cells are small and pigmented; at the vegetative pole they are large, pale and laden with yolk. If small regions of the surface of the blastula are stained with a vital dye, their fate in subsequent development may be followed. It is thus possible to say, for instance, that the cells of the animal pole normally give rise to ectoderm. From the results of many such experiments a map indicating presumptive fates may be drawn .
Gastrulation begins when a small indented crease, the dorsal lip of the blastopore, appears on one side of the blastula. The crease is formed by cells changing shape and pushing inward from the surface (invagination). Gastrulation entails spreading on the animal pole cells (the future ectoderm) over the vegetative cells (a process called epiboly). Sheets of cells at junction of vegetal and animal pole begin migrating in along the blastocoele. The site of this invagination is the blastopore (primitive mouth), which forms on the dorsal side of the embryo. .The presumptive (future) endoderm, notochord and mesoderm of the marginal zone move inside at this site through the dorsal lip of the blastopore. (Fig.15). These cell movements result in obliteration of the blastocele and the formation of another cavity – the gastrocele (archenteron) – which will become the gut. The gastrocele communicates with the exterior through the blastopore. The roof of the gastrocele consists of the chorda-mesoderm material, which now lies in contact with the overlying presumptive nerve tube.
The region of the blastopore through which invagination of chorda-mesoderm is occurring, is its dorsal lip. General body mesoderm proliferates from the lateral lips of the blastopore which approach each other and fuse, forming the primitive streak. The original blastopore is thus divided by the primitive streak into dorsal and ventral orifices (foramens).
Expressed in general terms, gastrulation is a complex series of morphogenetic movements of cell groups leading to the establishment of the primary germ layers and bringing the presumptive organ rudiments (notochord, nerve tube, somites, and primitive gut) into their definitive position. Other major changes during gastrulation:
Heavy RNA transcription using embryonic genes;
Cells starts major differentiation processes losing their pluripotentiality.
Gastrulation is regulated by homeobox genes and their proteins – morphogens.
THE EMBRYONIC DEVELOPMENT IN BIRDS
Birds produce few, large, yolk-rich eggs with protective membranes and shells. Thus, the ova in birds are referred to as polylecithal, pronounced telolecithal ova.
Cleavage. During incomplete (meroblastic, partial), diskoidal cleavage of the telolecithal egg the yolk does not divide. Cytoplasmic divisions are limited to the animal pole of the zygote (Fig.16)/ A series of cleavage divisions result in appearance of blastomeres. Blastomeres are arranged in the form of a disk, which lies over the yolk mass. This blastula is termed as a diskoblastula. The roof of the diskoblastula is formed by the blastodisk, and the bottom is the superficial layer of non-dividing yolk, which is termed as periblast.
 Fig.16.
Fig.16.
Gastrulation in birds proceeds by combination of delamination and immigration.
The first phase of gastrulation is delamination (Fig.17); it results in splitting off the internal layer – hypoblast (primitive endoderm), which will give rise to the extra-embryonic endoderm. At first the gut endoderm consists of round or irregular-shaped cells, which are arranged disorderly. Gradually the cells close up and form more or less regular single-layered plate. The rest of the germ disk forms epiblast (primitive ectoderm). Subsequently a cleft arises, which distinctly separates hypoblast from epiblast. The blastocoele is the thin cavity between the epiblast and hypoblast.
Further, cells of the central part of epiblast proliferate intensively, close up tightly and become more highly-prismatic in shape, forming the thickening of the central part of blastodisk. The epiblast contains the cell material of presumptive epidermal ectoderm, notochord, neural tube and mesoderm and endoderm of the embryo body. The thickened part of the blastodisk, consisting of two layers (the external one - epiblast, and the internal one - hypoblast), is termed as the germ disk. The germ disk is just the material, which forms the embryo body. The marginal parts of the blastodisk are extra-embryonic ecto- meso- and endoderm.
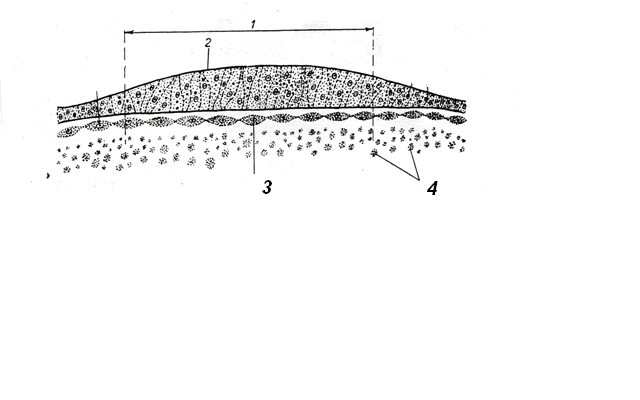
Fig.17. The diskoblastula in the chick (the vertical section).