

Основы альгологии и микологии Охапкин
.pdfся трабекулами или балками. Балки расположены радиально и перпендикулярно поверхности слоевища. Они увеличивают механическую прочность таллома и внутреннюю поверхность, выстланную цитоплазмой, что повышает интенсивность обмена веществ водоросли.
Кроме вегетативного размножения у каулерпы известен половой процесс. В особых плодущих участках ассимиляторов, не отделенных от остального таллома перегородками, образуются гаметы. Они выходят в воду через отверстия в специальных бугорках или сосочках, возникающих на поверхности плодущих участков. У большинства видов отмечена гетерогамия, после слияния гамет диплоидная зигота без периода покоя прорастает в новое растение. Происходит ли перед образованием гамет мейоз, остается неясным. Виды каулерпы распространены в тропических морях, и лишь немногие, в том числе и Caulerpa prolifera, обитают в Средиземном море.
Представители класса Dasicladophyceae – Дазикладофициевые –
при наличии сифональной структуры таллома отличаются радиальной симметрией и волосовидными выростами, которые отчасти могут сбрасываться, оставляя при этом рубцы. Центральная ось слоевища несет мутовчато-расположенные боковые ветви. Виды этого класса встречаются исключительно в морях.
Слоевище водорослей из порядка Dasicladales – Дазикладовые – представляет собой одну гигантскую радиально-симметричную клетку
исостоит из неразветвленной центральной оси, от которой мутовками отходят короткие боковые веточки. Эти ветви могут быть простыми или разветвленными и часто заканчиваются гаметангием. Клеточная стенка преимущественно из маннана. Талломы одноядерные на протяжении вегетационной фазы жизненного цикла. Прикрепляются водоросли к субстрату системой разветвленных ризоидов. Характерно отложение карбоната кальция в клеточной оболочке. Хлоропласты в талломе многочисленные дисковидные или эллипсоидные с пиреноидами или без них. Зерна запасных полисахаридов (фруктан и крахмал) откладываются
ив цитоплазме, что не характерно для других зеленых водорослей. Дазикладовые – типичные обитатели тропических и субтропических морей.
Дазикладус (Dasycladus clavaeformis) (рис. 124) – обитатель Среди-
земного моря, образует густые заросли на небольших глубинах.
231
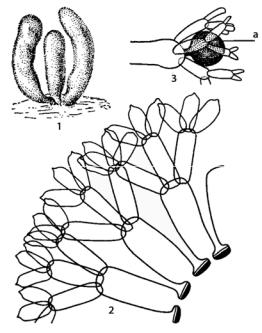
Рис. 124. Dasycladus:
1 – внешний вид слоевища;
2 – часть поперечного среза через слоевище; 3 – вершина веточки с гаметангием (а)
Таллом состоит из центральной оси, имеющей вид узкого пузыря длиной до 5 см, густо покрытого на всем протяжении, кроме самого основания, мутовками из 10–15 боковых веточек. Каждая боковая веточка, в свою очередь, дважды или трижды ветвится. Веточки второго и третьего порядка располагаются также мутовкой на вершине предыдущего сегмента и постепенно утончаются. Конечные веточки не срастаются, оставаясь свободными. Между отдельными сегментами перегородок нет и остается связь в виде узкого протоплазматического тяжа. Стенка центрального пузыря между местами отхождения веточек первого порядка утолщена, слоиста и ее наружные слои пропитаны карбонатом кальция. На верхушках веточек первого порядка образуются гаметангии. Перед обособлением гаметангиев от слоевища в них из соседних веточек перемещается цитоплазма с хлоропластами. В результате этого гаметангии становятся ярко-зелеными, а прилегающие к ним части таллома – бледными. В гаметангиях образуются двужгутиковые изогаметы, которые после выхода в воду попарно копулируют; зигота без периода покоя прорастает в новый таллом.
232
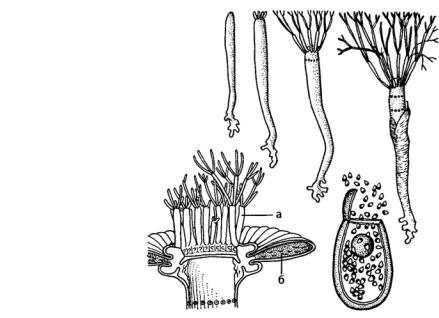
Рис. 125. Разные стадии развития, продольный разрез через зонтик и выход гамет из зрелой
цисты Acetabularia:
а– стерильные побеги;
б– гаметангий
Уацетабулярии (Acetabularia) (рис. 125) вертикальная ось достигает 3–15 см, а у части видов до 20 см длиной и прикрепляется к субстрату с помощью системы разветвленных ризоидов.
На вершине центральной оси формируется одна или несколько мутовок ветвящихся боковых веточек. В целом таллом ацетабулярии напоминает зонтик, особенно на репродуктивной стадии развития. Большая часть клетки занята вакуолью с клеточным соком, в постенном слое цитоплазмы расположены многочисленные хлоропласты. К концу первого года вегетации вертикальная часть слоевища отмирает, зимует лишь многолетняя ризоидная система. На второй год из ризоидной системы снова формируется одна или несколько стерильных вертикальных осей с мутовками боковых веточек. В природе этот процесс может происходить 2–3 года, затем наступает период плодоношения и вместо стерильных боковых осей образуются гаметангии.
Усредиземноморской Acetabularia mediterranea гаметангии мешковидные и срастаются своими боками, образуя подобие шляпки или зонтика. У Acetabularia crenulata, встречающейся в Карибском море, гаме-
233
тангии не срастаются, каждый из них имеет форму банана, и вся совокупность гаметангиев радиально расположена на вершине таллома. Гаметангии до определенного времени сообщаются с полостью центральной оси таллома. В период вегетативного роста диплоидное ядро находится в ризоиде, затем оно после мейоза распадается на большое количество гаплоидных вторичных ядер, которые токами цитоплазмы переносятся в гаметангии. После этого гаметангии отделяются от таллома пробкой, и их многоядерное содержимое распадается на одноядерные цисты. Цисты освобождаются из гаметангиев, а затем после ряда ядерных делений формируются двужгутиковые изогаметы. Зрелые цисты вскрываются крышечкой, через которую изогаметы выплывают в воду. После копуляции гамет зигота развивается в диплоидное растение.
Локализация единственного крупного ядра в ризоиде, его исключительная живучесть, способность к регенерации таллома при его поражении, большие размеры таллома делают ацетабулярию излюбленным объектом различных опытов по изучению роли ядра в формировании таллома, влиянию цитоплазмы на ядро и др. На опытах с ацетабулярией было продемонстрировано наличие ДНК в митохондриях и хлоропластах, влияние АТФ на движение цитоплазмы и др. В культуре можно получить половозрелые растения за полгода.
Таллом водорослей класса Trentepohliophyceae – Трентеполиофи-
циевые – нитчатый, часто гетеротрихальный, т.е. разделяющийся на стелющиеся и восходящие нити, которые могут быть разветвленными. У некоторых видов стелющиеся нити могут срастаться в плоский диск,
как у некоторых представителей Klebsormidiophyceae (Coleochaete, см.
ниже). Цитокинез происходит с образованием фрагмопласта, клеточной пластинки и плазмодесм. У жгутиковых стадий четыре микротрубочковых корешка расположены в клетке крестообразно. Базальные тела жгутиков ориентированы против часовой стрелки, но каждое базальное тело лежит на многослойной структуре (многослойная структура характерна для жгутиков харовых водорослей и высших растений, но у трентеполиофициевых имеет ряд отличий).
Порядок Trentepohliales – Трентеполиевые – как и класс, назван по имени ведущего рода трентеполия – Trentepohlia. Он объединяет главным образом гетеротрихальных представителей, имеющих таллом в виде разветвленных ползучих, приподнимающихся или прямостоячих нитей, разрастающихся на субстрате в порошковидные налеты или войлочные, подушковидные дерновинки. Зооспорангии у трентеполиевых
234
по форме отличаются от вегетативных клеток. Половое размножение происходит путем копуляции двужгутиковых изогамет.
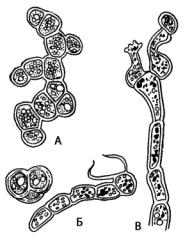
Род трентеполия (Trentepohlia) (рис. 126) относится к аэрофитным водорослям; образует порошковатые скопления оранжевого или бурого цвета на камнях, стволах деревьев, деревянных постройках, а тропические виды – на листьях.
Большинство представителей рода имеет гетеротрихальный таллом, состоящий из стелющихся по коре нитей и отрастающих от них вверх коротких ветвящихся фрагментов. У некоторых видов вертикальная система нитей представлена слабо. Клетки горизонтальной части таллома неправильно шаровидной формы с толстой, часто слоистой оболочкой. Восходящие нити имеют более вытянутые овальные или почти цилиндрические клетки. В клетках содержится много дисковидных или лентовидных хлоропластов без пиреноидов и большое количество липидных капель, окрашенных гематохромом в коричнево-красный цвет. Липидные капли полностью заполняют клетку и маскируют зеленую окраску хлоропластов. В сырое, дождливое и прохладное лето талломы
трентеполии более интенсивно растут, масло расходуется и хлоропласты становятся более заметны. Клетки одноядерные, но у старых количество ядер возрастает. Трентеполия легко размножается вегетативно – фрагментацией нитей на отдельные участки или клетки, которые переносятся ветром или каплями дождя. При попадании на субстрат эти фрагменты дают начало новым талломам.
Бесполое размножение осуществляется четырехжгутиковыми зооспорами, образующимися в крючковидных, овальных или шаровидных зооспорангиях на концах вертикальных нитей. Зооспорангии сидят на особых субспорангиальных клетках или клетках-ножках. При созрева-
235
нии зооспорангии становятся многоядерными, отделяются от таллома, переносятся ветром и при попадании в капельно-жидкую воду прорастают зооспорами. Половое размножение встречается редко. Гаметангии шаровидные или эллипсоидные, сидячие, разносятся ветром в целом виде и в присутствии воды прорастают двужгутиковыми изогаметами. Имеются сведения о наличии у трентеполии гапло-диплобионтного жизненного цикла со спорической редукцией и изоморфной сменой форм развития. Копуляция гамет наблюдается редко, большей частью они партеногенетически прорастают на коре деревьев в новые нити, как и зооспоры.
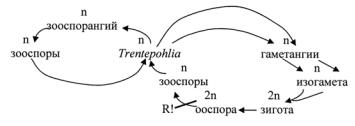
Цикл развития Trentepohlia
Многие виды Trentepohlia являются водорослевыми компонентами лишайников, а также поселяются на коре деревьев, каменистом субстрате, деревянных постройках, где образуют налеты красного, коричневого или желтоватого цвета. Обильно представлена эта водоросль в тропиках, где создает значительные проблемы, повреждая различные сооружения из бетона. ВРоссиивидыэтогородараспространеныповсеместно.
К классу Chlorophyceae – Хлорофициевые или Зеленые водорос-
ли – относятся одноклеточные, колониальные и многоклеточные водоросли с монадным, пальмеллоидным, коккоидным, нитчатым, разнонитчатым, сифональным типами организации таллома. Клеточная стенка у монадных видов состоит из гликопротеинов, у неподвижных представителей – из полисахаридов, в том числе из целлюлозы. При клеточном делении новые поперечные перегородки возникают с участием фикопласта, у некоторых формируется клеточная пластинка с плазмодесмами. Жгутиковые корешки расположены крестообразно, базальные тела ориентированы по типу 1–7 или 12–6. Размножение вегетативное, бесполое (зооспоры, апланоспоры, автоспоры) и половое (хологамия, изогамия, гетерогамия, оогамия). Жизненный цикл гаплобионтный с
236
зиготической редукцией. Виды этого класса обитают преимущественно в пресных водах, незначительная их часть – в солоноватых и морских водах или даже во вневодных местообитаниях.
К порядку Volvocales – Вольвоксовые – относятся монадные, подвижные, одноклеточные, колониальные или ценобиальные водоросли с клетками, построенными, главным образом, по типу одноосевой симметрии, реже клетки являются двусторонне симметричными; в виде исключения встречаются и асимметричные формы. Особи микроскопические, и только колонии вольвокса (Volvox) могут достигать 2 мм в диаметре. Клетки разнообразной формы, покрытые клеточной оболочкой или без нее. Клеточная оболочка обычно многослойная, состоит из гликопротеина, целлюлоза не обнаружена. Часто оболочка инкрустирована солями кальция, железа и марганца и у ряда видов состоит из отдельных фрагментов (например, у факотуса – Phacotus). Жгутиков 2–4 или 8 изоконтных, изоморфных и изодинамичных. Ядро одно, шаровидное или линзовидное с ядрышком, обычно находится спереди или в центре клетки. Хлоропласты по форме могут быть очень разнообразными и по расположению в клетке пристенными или центральными. У части видов при наступлении экстремальных условий (в сильно засоленных водоемах, при развитии на снегу) наблюдается накопление каротиноидов, окрашивающих клетки в оранжевый цвет (Chlamydomonas nivalis, Dunaliella salina). Как у любых монадных водорослей, в клетках вольвоксовых имеются сократительные вакуоли, стигма, а также непульсирующие вакуоли, митохондрии, аппарат Гольджи, эндоплазматическая сеть и различные включения. У большинства колониальных вольвоксовых образуются ценобии, и только у немногих представителей число клеток в колонии может меняться в течение вегетативной фазы.
Размножение бесполое и половое. Безоболочковые формы размножаются продольным делением клетки надвое, а виды с твердой клеточной оболочкой – посредством зооспор. Редко наблюдается фрагментация колоний (у неценобиальных), ценобиальные образуют дочерние колонии внутри материнской особи. Половой процесс, от хологамии до оогамии. Одноклеточные и колониальные вольвоксовые при наступлении неблагоприятных условий могут переходить в пальмеллевидное состояние.
Типичным представителем вольвоксовых является хламидомонас (Chlamydomonas) (рис. 127), многочисленные виды которого (до 500) обитают в лужах, канавах, других мелких водоемах, часто загрязненных органическими веществами.
237
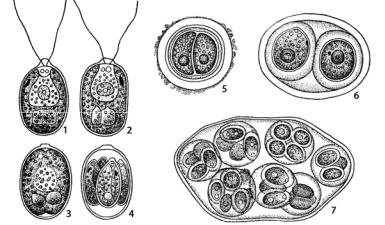
Рис. 127. Chlamydomonas: 1 – внешний вид клетки;
2–4 – процесс образования зооспор; 5–7 – пальмеллевидное состояние и размножение внутри слизи
Клетки хламидомонаса округлые или эллипсоидные с прозрачной оболочкой, на переднем конце с двумя равными жгутиками, с помощью которых клетка активно плавает в толще воды. Ядро одно, хлоропласт у большого количества видов чашевидный с глазком и пиреноидом. В передней части клетки находятся сократительные вакуоли, которые поддерживают постоянство осмотического давления протопласта. Пульсирующих вакуолей две, спереди, реже их много. У части видов на переднем конце имеется вздутие оболочки различной формы (носик), чаще в виде сосочка, по бокам которого выходят жгутики.
Глазок обычно расположен на переднем крае хлоропласта, иногда в середине или в задней его части. Составляющие глазок пигментные глобулы вместе образуют 3–8 рядов. При переходе в пальмеллевидное состояние клетки теряют подвижность, сбрасывают жгутики и оболочка их ослизняется. Втакомсостоянииклеткимогутделиться(рис. 127: 5–7).
При бесполом размножении клетки теряют жгутики, и протопласт делится на 2–4–8 частей, которые внутри материнской особи вырабатывают жгутики, окружаются оболочкой и превращаются в зооспоры, высвобождающиеся при ослизнении ее оболочки. Лизису (ослизнению)
238
стенки материнской клетки способствует особое вещество – автолизин спорангиальных стенок, выделяемое молодыми зооспорами.
Половой процесс у большинства видов хламидомонад изогамный
(рис. 128: А, Б).
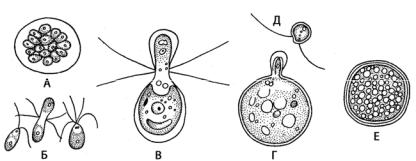
Рис. 128. Половой процеес у Chlamydomonas:
А– образование изогамет; Б – изогамия у Chlamydomonas steinii;
В– гетерогамия у Chlamydomonas braunii; Г – оогамия у Chlamydomonas coccifera; Д – мужская гамета Chlamydomonas coccifera; Е – зигота
Гаметы в числе 32–64 образуются в материнской клетке, как и зооспоры, но меньших размеров. У многих видов наблюдается гетероталлизм, т.е. половой процесс возможен только при слиянии «+» и «–» гамет, обладающих различными биохимическими и физиологическими признаками. Зигота покрывается многослойной оболочкой и переходит в состояние покоя. Большинство видов является обитателями луж с загрязненной водой. Некоторые, развиваясь в массовом количестве, могут вызывать «цветение» воды. В основном виды свободноплавающие и только немногие живут прикрепленно к коловраткам и ракообразным. Chlamydomonas nivalis вызывает красную окраску снега в высокогорьях и в Арктике за счет большого количества каротиноидов.
У некоторых хламидомонад наблюдается гетеро- и оогамия (рис. 128: В–Е). Русский ботаник И.Н. Горожанкин у Chlamydomonas braunii описал гетерогамный половой процесс, при котором более крупные женские гаметы формировались по четыре в материнской клетке, а более мелкие мужские – в числе восьми в другой клетке. У Chlamydomonas coccifera И.Н. Горожанкин установил оогамный половой процесс. У этого вида одни клетки теряют жгутики и превращаются в неподвижные яйцеклетки, а другие клетки образуют большое количество мелких
239

двужгутиковых мужских гамет, которые оплодотворяют яйцеклетку. Позднее оогамия была описана у других одноклеточных вольвоксовых – хлорогониума (Chlorogonium) и картерии (Carteria) (рис. 129: 1, 2).
Цикл развития Chlamydomonas
Рис. 129. Chlorogonium (1), Carteria (2) и Dunaliella (3)
У видов рода картерия (Carteria) (рис. 129: 2) клетка несет по 4 равных жгутика и по строению очень близка к клеткам хламидомонад. У хлорогониума клетки сильно вытянуты в длину, веретеновидные, иногда игловидные. У дуналиеллы (Dunaliella) (рис. 129: 3), встречающейся в сильно засоленных (гипергалинных) водоемах, например, в некоторых водоемах Крыма и Присивашья, клеточной стенки нет, она покрыта только плазмалеммой.
240