

Смертельні токсодози залежно від шляху надходження ор
|
|
Шлях надходження | ||
|
ОР |
інгаляційний, г х хв/м3 |
через шкіру, мг/кг |
через ШКТ‚ мг/кг |
|
Зарин Зоман V-гази |
0,1 0,03 0,004 |
7-9 1-1,5 0,05 |
1,5 0,5 0,009 |
2.2. Механізм дії і патогенез інтоксикації
У механiзмi дiї ФОР розрiзняють антихолiнестеразну i неантихолiнестеразну теорiю токсичної дiї.
Основною вважають антихолiнестеразну дiю, суть якої полягає у пригнiченнi ферменту ацетилхолiнестерази (АХЕ), який мiститься в тканинах нервової системи, еритроцитах, скелетних м’язах, що приводить до припинення або значного зменшення гiдролiзу медiатору ― ацетилхолiну (АХ), накопичення якого в синапсах супроводжується наступним збудженням i перезбудженням холiнергiчної нервової системи.
Холiнестераза (ХЕ) є простим бiлком-протеїном, який складається лише з амiнокислот і вмiщує у собі вiд 30 до 50 активних центрiв. Кожен активний центр має 2 активнi дiлянки: анiонну (-), яка зв’язується з катiонною (+) групою ацетилхолiну, i естеразну, утворену гiдроксилом амiнокислоти серину та iмiдазольним кiльцем гістидану (акцептором i донором протона), яка зв’язується зi складноефiрною групою ацетилхолiну з послiдовним розривом в АХ ефiрного зв’язку з утворенням холiну i оцтової кислоти.
Гiдролiз АХ триває близько 0,002 сек. Так, на поверхнi кiнцевої пластинки м’язу мiститься така кiлькiсть АХЕ, яка здатна розщепити 1 млн молекул АХ за 1 м/сек, що в 1000 раз в перевищує число молекул АХ, необхiдне для деполяризацiї кiнцевої пластинки (Nachmansahn, 1940). Хiмiчна реакцiя тече таким чином:
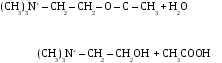
Ацетилхолiн є медiатором (передавачем) нервових iмпульсiв у холiнергiчнiй нервовiй системi (нейронах). Синтез АХ здiйснюється в пресинаптичному кiнцi шляхом переносу ацетильноі групи ацетiл-КоА на холiн за допомогою ферменту холiнацетилази з участю АТФ.

В станi спокою ацетилхолiн міститься в синаптичних пухирцях у зв’язаному станi з бiлком, кожний пухир дiаметром близько 80 нм мiстить до 40 тис. молекул ацетилхолiну (один квант медiатору). Пiд впливом iмпульсу видiляється приблизно 100-200 квантiв медiатору ― кiлькiсть, достатня для iнiцiювання потенцiалу дiї (ПД).
При проходженнi iмпульсу пiд впливом току iонiв Са (для викиду вмiсту одного пухирця (кванта) потрiбно приблизно чотири iони кальцiю), якi проникають iз синаптичної щiлини через пресинаптичну мембрану в клiтину.
Вiдбувається вивільнення окремими порцiями iз пухирців АХ‚ що виходить у синаптичну щiлину, вступає у взаємодiю з холiнорецептором постсинаптичної мембрани. При цьому вiдбувається її деполяризацiя внаслiдок перерозподiлу iонiв натрiю всередині клiтини, а калію назовнi, знижуючи її (мембранний) потенцiал і утворюючи таким чином потенцiал дiї (спайка, яка становить приблизько +30 мВ).
Тобто iмпульс передається на інший нейрон або ефекторний орган i забезпечує виконання певних функцiй органа (в нервовiй клiтинi ПД розповсюджується по аксону на інший нейрон, в м’язевiй або секреторнiй клiтинi ПД викликає визволення внутрiшньоклiтинно зв’язаних iонiв Са2+ i, вiдповiдно, скорочення чи секрецiю. Тривалiсть спайки не перевищує 1 мсек.
Пiсля проведення iмпульсу в клiтинi вiдновлюється стан спокою. В цей перiод iони натрiю, якi проникли в нейрон при збудженнi, замiнюються iонами калiю. Цей процес проходить проти градiєнта концентрацiї, так як iонiв натрiю у середовищi, яке оточує нейрони, набагато бiльше‚ нiж в клiтинi пiсля моменту її збудження.
Перехiд iонiв натрiю проти градiєнта концентрацiї здiйснюється за допомогою натрiєвого насоса з витратою енергiї АТФ. Таким чином, вiдновлюється вихiдна концентрацiя катiонiв калiю i натрiю всерединi клiтини (аксона), i мембрана набуває потенцiалу спокою‚ який становить приблизно від мВ – 50 до 90мВ.
Холiнорецептори ― це бiлково-лiпiднi комплекси. Залежно вiд чутливостi до тiєї чи iншої групи хiмiчних сполук, вони подiляються на мускариночутливi (М-холiнорецептори) i нiкотиночутливi (Н-холiнорецептори).
М-холiнорецептори розташованi на постсинаптичнiй мембранi у синапсах усiх постганглiонарних парасимпатичних нервiв, симпатичних нервiв, якi інервують потовi залози, частково шлунок i матку, а також у нейронейрональних синапсах головного мозку, у клiтинах Реншоу i блокуються холiнолiтиками. Н-холiнорецептори містяться на постсинаптичнiй мембранi скелетних м’язiв, ганглiях, нейронах спинного мозку, в каротидному клубочку, хромофiнних клiтинах мозкової речовини надниркових залоз i блокуються ганглiоблокаторами, кураре, а в ЦНС ― пентафеном.
Мускариновi i нiкотиновi рецептори працюють у рiзночасових режимах. Одиничний нервовий iмпульс в мускариновому рецепторi триває близько 500 мсек, а в нiкотиновому ― 0,2 мсек i навiть у повiльних ганглiонарних синапсах - лише 60 мсек.
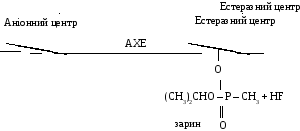
Фосфороорганiчнi сполуки iмiтують реакцiйну здатнiсть ацетилхолiну. Так, зарин, зоман iмiтують лише ефiрну частину молекули медiатору. В молекулах цих ОР мiститься группа (Р=О), яка поляризована, як i карбонiльна група (С=О) ацетилхолiну. Взаємодiя зарину i зоману з естеразною дiлянкою ферменту призводить до фосфорилювання серинового гiдроксилу АХЕ, внаслiдок чого фосфорильована АХЕ не здатна гiдролiзувати АХ.
Взаємодія зарину з ацетилхолінестеразою:
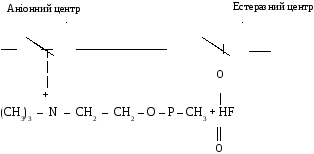
Бiльш сильна антихолiнестеразна дiя є у фосфорилтiохолiнiв типу V - газiв, якi завдяки присутностi амiногрупи здатнi з’єднуватися як з естеразним гiдроксилом серину, так i з анiонною дiлянкою ферменту.
Взаємодія метилфторфосфорилхоліну з ацетилхолінестеразою:
Тривалiсть дiї ФОР, як i iнших антихолiнестеразних речовин, визначається швидкiстю вiдновлення активностi АХЕ, швидкiстю розпаду комплексу IФ (iнгiбiтор - фермент). Поступова втрата фермент - iнгiбiторним комплексом здатностi до дисоцiацiї визначається як “старiння” АХЕ. Час повного “старiння” фосфорильованої АХЕ коливається вiд декiлькох хвилин до декiлькох дiб. Особливо швидко проходить “старiння” зв’язку у зоману. У першi хвилини i години пiсля отруєння можливе вiдщеплення ОР вiд ферменту за допомогою реактиваторiв АХЕ, але пізніше внаслiдок “старiння” фосфорильованого фермента вiдщеплення ОР стає неможливим. Оборотнi iнгiбiтори АХЕ (прозерин, галантамiн) зв’язуються з АХЕ немiцно‚ i активнiсть ії швидко вiдновлюється.
Існує пряма залежність мiж ступенем пригнiчення холiнестерази i токсичнiстю, таким чином, чим сильнiша антихолiнестеразна дiя речовини, тим бiльша її токсичнiсть. Так, при легкому ступенi ураження активнiсть АХЕ пригнiчена на 30-50 %, при середньому ― на 50-70 %, при важкому ― бiльше ніж на 70 %.
Визначення ступеню пригнiчення АХЕ використовується в лабораторнiй практицi для дiагностики отруєння ФОР, а також при iндикацiї для визначення наявностi отрути у водi, харчових продуктах та iнших середовищах.