

6 курс / Гастроэнтерология / Российский_журнал_гастроэнтерологии,_гепатологии,_колопроктологии (60)
.pdfСодержание |
|
|
|
|
3, 2010 |
Редакционная |
|
|
|
|
|
В.Т. Ивашкин |
|
|
|
|
|
Ядерные рецепторы и патология печени. Часть 1-я.......................................................... |
|
4 |
|||
Лекции и обзоры |
|
|
|
|
|
Э.Ф. Баринов, О.Н. Сулаева |
|
|
|
|
|
Гастроинтестинальные миофибробласты – роль в регуляции физиологической |
|
||||
активности и репарации желудочно-кишечного тракта...................................................... |
|
9 |
|||
С.Н. Маммаев, А.М. Каримова, Т.Э. Ильясова, А.Ш. Хасаев |
|
|
|||
Цирротическая кардиомиопатия................................................................................... |
|
|
|
19 |
|
Е.А. Федосьина, М.В. Маевская |
|
|
|
|
|
Применение урсодезоксихолевой кислоты при неалкогольном и алкогольном |
|
||||
стеатогепатите............................................................................................................ |
|
|
|
|
29 |
Оригинальные исследования |
|
|
|
|
|
И.В. Козлова, Е.В. Граушкина |
|
|
|
|
|
Клинические особенности и диагностические критерии патологии |
|
||||
эзофагогастродуоденальной зоны у больных желчнокаменной болезнью, |
|
||||
перенесших операцию холецистэктомии |
......................................................................... |
|
37 |
||
И.Ю. Пирогова |
|
|
|
|
|
Скрининговое обследование больных диффузными ........................заболеваниями печени |
46 |
||||
|
|
|
|
RU |
|
Национальная школа гастроэнтерологов, гепатологов |
|
|
|||
М.А. Евсеев, И.М. Клишин |
|
|
|
|
|
|
|
|
- VESTI |
|
|
Эффективность антисекреторной терапии ингибиторами.протонной помпы |
|
||||
при гастродуоденальных язвенных кровотечениях.......................................................... |
|
55 |
|||
Обмен опытом |
|
|
|
|
|
А.А. Шептулин, М.А. Визе-Хрипунова |
|
|
|
|
|
Современные возможности применения ...............препаратов висмута в гастроэнтерологии |
63 |
||||
|
|
.M |
|
|
|
Ю.О. Шульпекова |
|
|
|
|
|
Кисломолочные бактерии: роль в регуляции ..............................кишечной перистальтики |
68 |
||||
|
WWW |
|
|
|
|
А.В. Лапшин, Н.И. Кокина, М.В. Маевская, И.А. Соколина, В.Т. Ивашкин |
|
||||
Саркоидоподобная реакция у пациентки с хроническим гепатитом С |
|
||||
на фоне противовирусной терапии................................................................................ |
|
|
|
74 |
|
Новости колопроктологии |
|
|
|
|
|
С.Н. Наврузов, С.Б. Абдужаппаров, Б.С. Наврузов |
|
|
|||
Низкие и ультранизкие анастомозы в сочетании с различными вариантами |
|
||||
лимфодиссекции при хирургическом лечении ....................................рака прямой кишки |
79 |
||||
А.М. Кузьминов, Ю.Ю. Чубаров, А.А. Тихонов, Ш.Т. Минбаев, В.Ю. Королик |
|
||||
Циркулярная слизисто-подслизистая резекция нижнеампулярного отдела |
|
||||
прямой кишки (степлерная геморроидпексия) при лечении больных |
|
||||
хроническим геморроем............................................................................................... |
|
|
|
|
82 |
Информация |
|
|
|
|
|
Резюме диссертаций: информация из ВАК ..........................................................России |
|
88 |
|||
РЖГГК он-лайн – www.gastro-j.ru |
|

Соntents |
|
|
|
3, 2010 |
Editorial |
|
|
|
|
V.T. Ivashkin |
|
|
|
|
Nuclear receptors and liver disease (the Part 1)........................................................................... |
|
4 |
||
The lectures and reviews |
|
|
|
|
E.F. Barinov, O.N. Sulayeva |
|
|
|
|
Gastrointestinal myofibroblasts: role in regulation of physiological activity |
|
|||
and reparation of gastro-intestinal tract. |
........................................................................... |
|
9 |
|
S.N. Mammayev, A.M. Karimova, T.E. Il’yasova, A.Sh. Khasayev |
|
|
||
Cirrhotic cardiomyopathy.............................................................................................. |
|
|
|
19 |
Ye.A. Fedos’ina, M.V. Mayevskaya |
|
|
|
|
Application of ursodeoxycholic acid at non-alcoholic and alcohol-induced |
|
|||
steatohepatitis.............................................................................................................. |
|
|
|
29 |
Original investigations |
|
|
|
|
I.V. Kozlova, Ye.V. Graushkina |
|
|
|
|
Clinical features and diagnostic criteria of diseases of esophagogastroduodenal zone |
|
|||
in patients with gallstone disease after cholecystectomy...................................................... |
|
37 |
||
I.Yu. Pirogova |
|
|
|
|
Screening investigation of patients with ..............................................diffuse liver diseases |
46 |
|||
National college of gastroenterologist, hepatologist |
|
|
||
M.A. Evseyev, I.M. Klishin |
|
|
|
|
Antisecretory treatment efficacy by proton pump inhibitors at gastroduodenal |
|
|||
|
|
- VESTI |
|
|
ulcer bleedings........................................................ |
............................................... |
|
RU |
55 |
Exchange of experience |
|
|
|
|
A.A. Sheptulin, M.A. Vise-Khripunova |
|
|
|
|
Modern potential of of bismuth in gastroenterology........................................................... |
|
63 |
||
Yu.O. Shulpekova |
|
|
|
|
Fermented-milk bacteria: role in intestinal ..........................................peristalsis regulation |
68 |
|||
|
WWW |
|
|
|
A.V. Lapshin, N.I. Kokina, M.V. Mayevskaya,.IMA. Sokolina, V.T. Ivashkin |
|
|||
Sarcoid-like reaction at the patient with chronic hepatitis C on a background |
|
|||
of antiviral therapy....................................................................................................... |
|
|
|
74 |
News of coloproctology |
|
|
|
|
S.N. Navruzov, S.B. Abduzhapparov, B.S. Navruzov |
|
|
||
Low and ultra-low anastomoses in combination to various variants of lymph |
|
|||
node dissection in rectal cancer....................................................................................... |
|
|
79 |
|
A.M. Kuzminov, Yu.Yu. Chubarov, A.A. Tikhonov, Sh.T. Minbayev, V.Yu. Korolik |
|
|||
Circular mucosal and submucosal resection of low-ampullary region |
|
|||
of the rectum (stapled hemorrhoidopexy) .......................at treatment of chronic hemorrhoids |
82 |
|||
Information |
|
|
|
|
Thesis abstracts: information from the Higher ...................................attestation commission |
88 |
|||
РЖГГК он-лайн – www.gastro-j.ru

Российский журнал Гастроэнтерологии, Гепатологии, Колопроктологии
№ 3 • Том 20 • 2010
Russian Journal of
Gastroenterology,
Hepatology,
Coloproctology
Volume 20 • № 3 • 2010
Учредитель:
Российская
гастроэнтерологическая
ассоциация
Издатель:
ООО «Издательский дом «М-Вести»
Периодичность издания:
1 раз в 2 месяца
Тираж: 3000 экз.
Подписной индекс: 73538
Журнал зарегистрирован Комитетом РФ по печати 15.12.1994 г.
(Регистрационный № 013128)
Информация о журнале
находится в Интернете на сайтах
www.gastro-j.ru, www.m-vesti.ru
Адрес:
119146, г. Москва, а/я 31, «ГАСТРО», Российский журнал гастроэнтерологии,
гепатологии, колопроктологии
Эл. почта: editorial@gastro-j.ru
Журнал входит в Перечень ведущих научных журналов и изданий ВАК Министерства образования и науки России, в которых должны быть опубликованы основные научные результаты диссертаций на соискание ученой степени доктора наук
Перепечатка материалов только с разрешения главного редактора и издателя
Ответственность за достоверность рекламных публикаций несут рекламодатели
Российский журнал гастроэнтерологии, гепатологии, колопроктологии
Состав редакционной коллегии и редакционного совета журнала
Главный редактор
В.Т.Ивашкин
Исполнительный директор проекта
Г.Г.Пискунов
Ответственный секретарь
Т.Л.Лапина
(Эл. почта: editorial@gastro-j.ru)
Редакционная коллегия |
|
|
||
Е.К.Баранская |
|
|
|
|
А.О.Буеверов |
|
|
|
|
С.А.Булгаков |
|
|
|
|
П.С.Ветшев |
|
|
|
|
Г.И.Воробьев |
|
|
|
|
О.М. Драпкина |
|
|
|
|
А.В.Калинин |
|
|
|
|
(зам. главного редактора) |
|
|
||
З.А.Лемешко |
|
|
|
|
А.Ф.Логинов |
|
|
|
|
И.В.Маев |
|
|
|
|
М.В.Маевская |
|
|
|
|
(зам. главного редактора) |
|
RU |
||
С.И.Рапопорт |
|
|
|
|
И.Г. Никитин |
|
|
|
|
А.В.Охлобыстин |
|
|
. |
|
Ю.М.Панцырев |
|
|
||
-VESTI |
|
|||
Ю.В.Тельных |
|
|||
|
|
|
|
|
А.С.Трухманов |
|
|
|
|
П.В. Царьков |
|
|
|
|
С.А.Чернякевич |
|
|
|
|
А.А.Шептулин |
|
|
|
|
.M |
|
|
|
|
(зам. главного редактора) |
|
|
||
О.С. Шифрин |
|
|
|
|
WWW |
|
|
|
|
Редакционный совет |
|
|
|
|
С.А.Алексеенко |
|
|
Хабаровск |
|
О.Я.Бабак |
|
|
Харьков |
|
Э.И.Белобородова |
|
|
Томск |
|
Э.Г.Григорян |
|
|
Ереван |
|
А.Р.Златкина |
|
|
Москва |
|
Г.Ф.Коротько |
|
|
Краснодар |
|
С.А.Курилович |
|
Новосибирск |
||
В.А.Максимов |
|
|
Москва |
|
С.Н.Маммаев |
|
|
Махачкала |
|
Ю.Х.Мараховский |
|
|
Минск |
|
Г.А.Минасян |
|
|
Ереван |
|
О.Н.Минушкин |
|
|
Москва |
|
И.А.Морозов |
|
|
Москва |
|
Ю.Г.Мухина |
|
|
Москва |
|
А.И.Пальцев |
|
Новосибирск |
||
Л.К.Пархоменко |
|
|
Харьков |
|
В.Д.Пасечников |
|
|
Ставрополь |
|
С.Д.Подымова |
|
|
Москва |
|
Г.В.Римарчук |
|
|
Москва |
|
В.И.Симоненков |
|
Санкт-Петербург |
||
А.В.Ткачев |
|
Ростов-на-Дону |
||
Е.Д.Федоров |
|
|
Москва |
|
И.Л.Халиф |
|
|
Москва |
|
Г.В.Цодиков |
|
|
Москва |
|
А.В.Шапошников |
|
Ростов-на-Дону |
||
Editor-in-chief
V.T.Ivashkin
Production Manager
G.G.Piskunov
Editorial Manager
T.L.Lapina
(E-mail: editorial@gastro-j.ru)
Editorial board
Ye.K.Baranskaya
A.O.Bueverov
S.A.Bulgakov
P.S.Vetshev
G.I.Vorobiev
O.M. Drapkina A.V.Kalinin
(deputy editor-in-chief) Z.A.Lemeshko A.F.Loginov I.V.Mayev M.V.Mayevskaya (deputy editor-in-chief) I.G. Nikitin A.V.Okhlobystin Yu.M.Pantsyrev S.I.Rapoport Yu.V.Tel’nykh A.S.Trukhmanov
P.V. Tzar’kov S.A.Chernyakevich A.A.Sheptulin
(deputy editor-in-chief) O.S. Shifrin
Editorial council |
|
S.А.Alexeyenko |
Khabarovsk |
O.Ya.Babak |
Kharkov |
E.I.Byeloborodova |
Tomsk |
E.G.Grigoryan |
Yerevan |
A.R.Zlatkina |
Moscow |
G.F.Korot’ko |
Krasnodar |
S.A.Kurilovich |
Novosibirsk |
V.A.Maximov |
Moscow |
S.N.Mammaev |
Machachkala |
Yu.Kh.Marakhovsky |
Minsk |
G.A.Minasyan |
Yerevan |
O.N.Minushkin |
Moscow |
I.A.Morozov |
Moscow |
Yu.G.Mukhina |
Moscow |
A.I.Pal'tsev |
Novosibirsk |
L.K.Parkhomenko |
Kharkov |
V.D.Pasyechnikov |
Stavropol |
S.D.Podymova |
Moscow |
G.V.Rimarchuk |
Moscow |
V.I.Simonenkov |
Saint-Petersburg |
A.V.Tkachev |
Rostov-on-Don |
Ye.D.Fedorov |
Moscow |
I.L.Khalif |
Moscow |
G.V.Tsodikov |
Moscow |
A.V.Shaposhnikov |
Rostov-on-Don |

Редакционная |
3, 2010 |
УДК 616.36-092
Ядерные рецепторы и патология печени
Часть 1-я
В.Т. Ивашкин
(Московская медицинская академия им. И.М. Сеченова)
Nuclear receptors and liver disease
(the Part 1)
V.T. Ivashkin
дерные рецепторы (ЯР) выступают участ |
основой создания новых эффективных средств |
|
никами всех важнейших функций печени. |
для лечения заболеваний печени. |
|
-VESTI |
|
|
ЯПо существу печень представляет собой |
Терапевтическими.RU |
мишенями могут стать вирус- |
гигантскую железу, которая производит или при- |
ные гепатиты, воспаление, фиброз, метаболический |
|
влекает лиганды этих рецепторов, включая жел- |
синдром, стеатоз печени, холестатические синдро- |
|
чные кислоты (ЖК), жирные кислоты, окислен- |
мы, холангиопатии, желчнокаменная болезнь, рак |
|
ные производные холестерина, ксенобиотические |
(рис. 1). В каждом из этих клинических сценариев |
|
индукторы метаболизма лекарств и ретиноиды. |
ЯР и КоР участвуют или как патогенетические, |
|
.M |
или как адаптационные составляющие. Достаточно |
|
Ядерные рецепторы в ответ на меняющиеся уров- |
||
ни их лигандов реагируют стандартными ком- |
сказать, что в метаболизме ЖК участвуют, по |
|
пенсаторными изменениями экспрессии генов, |
крайней мере, 5 ЯР (FXR, PXR, CAR, LXR, |
|
направленными на поддержание гомеостаза. |
VDR), каждый из которых регулируется разными |
|
Наиболее простыми и элегантными примерами |
желчными кислотами. Через гепатоциты прохо- |
|
этой функции выступают способность жирных |
дит большое число разнообразных эндобиотиков, |
|
кислот активировать пероксисомный пролифера- |
ксенобиотиков и липидов, которые одновременно |
|
тор-активируемый рецептор (PPAR)WWWи индуци- |
служат в качестве модуляторов ЯР и КоР. Эти же |
|
ровать их собственное окисление, а также актива- |
самые молекулы метаболизируются посредством |
|
ция токсическими ксенобиотиками прегнанового |
продуктов генов, регулируемых ЯР/КоР. |
|
Х рецептора (PXR) или конститутивного анд- |
Клинические наблюдения указывают на то, |
|
ростанпрегнанового рецептора (CAR) и индук- |
что большая группа пациентов достаточно успеш- |
|
ция их собственной детоксикации. |
но адаптируется к своему болезненному состо- |
|
Многие исследователи полагают, что мы при- |
янию, тогда как у других этого не происходит. |
|
ближаемся к окончанию интереснейшего периода |
Подобный факт свидетельствует о наличии у |
|
открытий новых ядерных рецепторных лигандов и |
человека механизмов адаптации преимущественно |
|
определения их первичных функций, однако наше |
в виде нутриционных и генетических составля- |
|
понимание этих функций, их взаимодействий и |
ющих. С другой стороны, экспериментальные |
|
влияний на физиологию и патологию человека |
исследования прямо указывают на существенную |
|
находится лишь на стадии начального формиро- |
роль определенных ЯР и КоР в патогенезе болез- |
|
вания. Современная информация о центральной |
ней и на возможность модификации патогенеза |
|
роли ядерных рецепторов и корегуляторов (КоР) |
заболеваний печени |
конкретными регуляторами |
физиологических функций в печени может стать |
ЯР/КоР. Знание механизмов действия модулято- |
|
Ивашкин Владимир Трофимович – академик РАМН, доктор медицинских наук, профессор, директор клиники пропедевтики внутренних болезней, гастроэнтерологии и гепатологии им. В.Х. Василенко, заведующий кафедрой пропедевтики внутренних болезней ГОУ ВПО ММА им. И.М. Сеченова Росздрава; контактная информация для переписки: v.ivashkin@gastro-j.ru; 119991, г. Москва, ул. Погодинская, д. 1, стр. 1, Клиника пропедевтики внутренних болезней, гастроэнтерологии и гепатологии им. В.Х. Василенко ГОУ ВПО ММА им. И.М. Сеченова Росздрава
|
РЖГГК он-лайн – www.gastro-j.ru |
|

3, 2010 |
|
|
|
Редакционная |
||
|
|
|
|
|
|
|
|
мые de novo из углеводов. Нарушение окисления |
|||||
Селективные модуляторы ER |
||||||
|
|
|
жирных кислот в митохондриях и пероксисо- |
|||
|
|
|
мах также может быть причиной их накопле- |
|||
|
|
|
ния в печени. Аккумуляция жира может быть, |
|||
|
|
|
кроме того, результатом нарушения секреции |
|||
|
|
|
частиц липопротеинов очень низкой плотнос- |
|||
|
|
|
ти (ЛПОНП). Секреция триглицеридов печени |
|||
|
|
|
требует сбалансированного соотношения фосфо- |
|||
|
|
|
липидов и апопротеинов (например, апоВ), и неа- |
|||
|
|
|
декватное поступление данных элементов может |
|||
|
|
|
нарушить секрецию триглицеридов. Каждый из |
|||
|
|
|
этих процессов очень тонко регулируется на |
|||
|
|
|
различных уровнях и включает активацию специ- |
|||
|
|
|
фических ЯР, таких как PPAR, LXR или SPEBP |
|||
|
|
|
(A. Vidal-Puig, 2009). |
|
||
|
|
|
В последнее время исследователей привлека- |
|||
|
|
|
||||
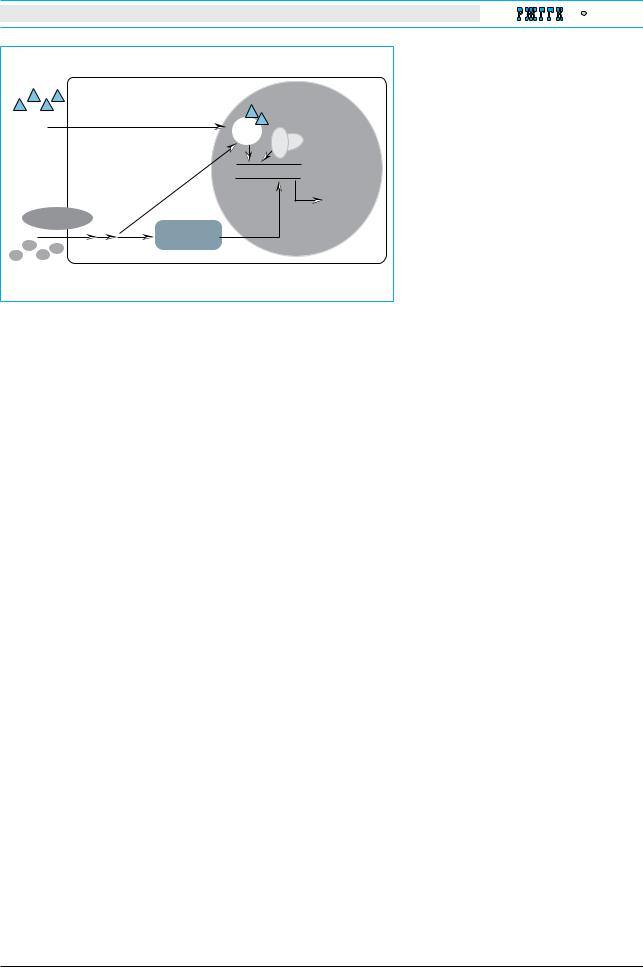
Рис. 1. Роль модуляторов ядерных рецепторов (ЯР) |
ет концепция ограничения или лимита объема |
|||||
и корегуляторов (КоР) могут выполнять разнооб- |
увеличения жировой ткани как фактора, сопря- |
|||||
разные эндобиотики, ксенобиотики, липиды. Эти |
гающего ожирение с метаболическим |
синдро- |
||||
молекулы метаболизируются посредством продук- |
||||||
мом и НАЖБП. Совершенно очевидно, что для |
||||||
ции регулируемых генов. Экспериментальные иссле- |
||||||
дования прямо указывают на существенную роль |
развития ожирения необходим положительный |
|||||
определенных ЯР и КоР в патогенезе болезней и на |
баланс энергии, возникающий при несбаланси- |
|||||
возможность модификации патогенеза заболеваний |
рованном избыточном потреблении пищи и недо- |
|||||
печени определенными регуляторами ЯР/КоР. |
статочном |
расходе энергии. Однако с меньшей |
||||
SERM – селективные модуляторы эстрогеновых |
||||||
долей вероятности можно предположить, что |
||||||
рецепторов; HDAC – деацетилазы гистонов; |
||||||
ER – эндоплазматический ретикулум |
прекращение.RUувеличения жировой массы на фоне |
|||||
|
|
|
сохраняющегося положительного энергетическо- |
|||
|
|
|
го дисбаланса становится основным фактором в |
|||
ров центральных ЯР/КоР-зависимых путей поз- |
развитии НАЖБП. В рамках еще одной концеп- |
|||||
воляет более предметно анализировать такие клю |
ции рассматривается значение отличий наборов |
|||||
чевые проблемы клиники, как прогрессирование |
ЯР, экспрессируемых в печени здоровых лиц |
|||||
|
|
-VESTI |
|
|
||
хронических вирусных гепатитов в направлении |
и пациентов с НАЖБП. В частности, разви- |
|||||
фиброза/цирроза или стеатоза в направлении |
тию НАЖБП может способствовать избыточная |
|||||
|
|
.M |
индукция липидами в печени генетических про- |
|||
стеатогепатита, формирование холестаза и цирро- |
||||||
за при холестатических синдромах и холангиопа- |
грамм первичной жировой ткани. Такая возмож- |
|||||
тиях и многие другие (рис. 2). |
|
ность показана на примере изоформы ядерного |
||||
|
|
|
рецептора PPARg2, которая в физиологических |
|||
Метаболический синдром |
условиях экспрессируется только в белой жиро- |
|||||
вой ткани, но при положительном энергетическом |
||||||
|
|
WWW |
||||
Неалкогольная жировая болезнь печени |
балансе индуцируется эктопически в печени, что |
|||||
(НАЖБП) представлена в человеческой популя- |
способствует аккумуляции жира. |
|
||||
ции очень широко и рассматривается в настоящее |
Привлекает внимание концепция, учитываю- |
|||||
время как индикатор метаболического синдро- |
щая значение специфических типов липидов в |
|||||
ма. Патогенетические механизмы неалкогольного |
развитии метаболических отклонений в печени, в |
|||||
стеатогепатита (НАСГ) полностью не раскры- |
частности резистентности к инсулину. Например, |
|||||
ты. «Гипотеза двух ударов» («two-hit hypothesis») |
увеличение |
содержания триглицеридов |
может |
|||
предполагает начальную фазу прогрессирующего |
отражать повышение липидной нагрузки и, сле- |
|||||
отложения жира в печени, за которой следует |
довательно, рассматриваться как положительный |
|||||
вторая фаза, характеризующаяся интенсивным |
адаптационный процесс, повышающий буферную |
|||||
оксидативным стрессом. Обе фазы в значительной |
емкость печени в отношении накопления энергии |
|||||
степени перекрывают друг друга, что существен- |
в менее токсической форме. Дополнительный |
|||||
но затрудняет дифференциацию специфических |
вклад в понимание сущности НАЖБП может |
|||||
первичных реакций от неспецифических компен- |
быть получен при изучении роли транскрипци- |
|||||
саторных механизмов. Начальный удар («initial |
онных факторов LXR, SREPS и PPAR, контро- |
|||||
hit») включает аккумуляцию в печени свободных |
лирующих баланс между липогенезом de novo в |
|||||
жирных кислот (СЖК) из различных источни- |
печени и окислением жирных кислот, регулируе- |
|||||
ков: СЖК жировой ткани, вышедшие в плазму |
мым малонил КоА. |
|
||||
крови; СЖК гидролизированных хиломикронов, |
За последние 5 лет в значительной мере пере- |
|||||
поступающих из кишечника; СЖК, синтезируе- |
осмыслена роль желчных кислот, и стало очевид- |
|||||
РЖГГК он-лайн – www.gastro-j.ru |
|
|
Редакционная |
|
|
|
|
|
|
|
|
|
|
|
3, 2010 |
||
|
Регуляция транскрипции генов |
|
|
|
ют |
важные функции и |
|
за |
рамками |
|||||
|
|
|
|
кишечно-печеночной оси и действуют |
||||||||||
Липофильные |
|
|
|
|
||||||||||
|
|
|
|
как системные метаболические интег- |
||||||||||
гормоны |
|
|
|
|
||||||||||
|
Ядерные |
|
|
раторы |
|
(K. Schoonjans, |
J. Auwerx, |
|||||||
|
рецепторы |
|
2009). |
В энтероэндокринных |
клетках |
|||||||||
|
|
|
|
|
||||||||||
|
|
|
Кофакторы |
|
TGR5 |
ассоциирован |
с |
опосредуемой |
||||||
|
|
|
|
|
желчными кислотами секрецией глю- |
|||||||||
|
|
Метаболический |
|
кагон-подобного пептида 1 (GLP-1; |
||||||||||
|
Гены |
|
glucagon-like peptide). |
GLP-1 пред- |
||||||||||
|
|
|
контроль |
|
ставляет собой инкретин со значитель- |
|||||||||
|
|
|
|
|
||||||||||
|
Сигнальный |
|
TXN |
|
ной |
антидиабетической |
|
активностью |
||||||
Рецептор |
каскад |
|
|
вследствие его способности повышать |
||||||||||
|
|
|
||||||||||||
|
TXN |
|
|
|
функцию |
панкреатических |
В-кле- |
|||||||
|
факторы |
|
|
|
||||||||||
|
|
|
|
ток. Эти данные указывают на роль |
||||||||||
|
|
|
|
|
||||||||||
Пептидные |
|
|
|
|
TGR5 не только в поддержании энер- |
|||||||||
гормоны |
|
|
|
|
гетического баланса, но и в контроле |
|||||||||
|
|
|
|
|
гомеостаза глюкозы. Если подытожить |
|||||||||
Рис. 2. В зависимости от строения гормона существуют два типа |
|
вышесказанное, то локус TGR5 может |
||||||||||||
|
рассматриваться как |
важный регион |
||||||||||||
взаимодействия. Если молекула гормона липофильна (например, |
|
|||||||||||||
стероидные гормоны), то она может проникать через липидный |
|
генома |
в |
поддержании |
|
нормального |
||||||||
слой наружной мембраны клеток-мишеней. Если молекула имеет |
|
функционирования |
|
панкреатических |
||||||||||
большие размеры или является полярной, то ее проникновение |
|
В-клеток и как привлекательная потен- |
||||||||||||
внутрь клетки невозможно. Поэтому для липофильных гормонов |
|
циальная мишень для лечения сахар- |
||||||||||||
рецепторы находятся внутри клеток-мишеней, а для гидрофильных |
||||||||||||||
ного диабета 2-го типа. |
|
|
|
|||||||||||
(в том числе для пептидных гормонов) – в наружной мембране. |
|
|
|
|
||||||||||
Для получения клеточного ответа на гормональный сигнал в слу- |
|
SHP |
(NROB2), |
|
представляющий |
|||||||||
чае гидрофильных молекул действует внутриклеточный механизм |
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
.RUсуперсемейство ядерных рецепторов, |
||||||||||
передачи сигнала. Связывание гормона с рецептором инициирует |
участвует в биологической регуляции |
|||||||||||||
каскад реакций, приводящих в конечном итоге к изменению скоро- |
||||||||||||||
нескольких важных функций печени. |
||||||||||||||
сти транскрипции регулируемого гена |
|
|
|
Функциональный |
или |
|
структурный |
|||||||
|
|
|
|
|
|
|||||||||
|
|
|
|
|
дефицит SHP приводит к уменьшению |
|||||||||
ным, что помимо их доказанного участия в абсор |
содержания липидов в печени и тормозит раз- |
|||||||||||||
|
|
-VESTI |
|
|
|
|
|
|
|
|
|
|||
бции пищевых жиров и поддержании гомеостаза |
витие стеатоза, вызываемого недостаточностью |
|||||||||||||
холестерина они выполняют функцию биологичес- |
лептина (L. Wang, 2009). SHP регулирует липид- |
|||||||||||||
|
.M |
ный гомеостаз печени путем контроля баланса |
||||||||||||
ких сигнальных молекул. Первые предположения |
||||||||||||||
о такой роли появились, когда был выявлен факт |
таких процессов, как экспорт, импорт, синтез и |
|||||||||||||
контроля желчными кислотами собственного син- |
окисление липидов. С другой стороны, продол- |
|||||||||||||
теза в печени посредством обратного ингибирова- |
жительное выключение функции SHP ускоряет |
|||||||||||||
ния ядерного фарнезоидного Х рецептора (FXR) |
прогрессирование гепатомы в эксперименте, что |
|||||||||||||
и ядерного короткого гетеродимерногоWWWпарт- |
сопровождается ускорением пролиферации гепа- |
|||||||||||||
нера (SHP; |
short heterodimer partner) – К. Lu |
тоцитов и угнетением их апоптоза. SHP рассмат- |
||||||||||||
и соавт., 2000. Далее было продемонстрировано, |
ривают в качестве репрессора транскрипции регу- |
|||||||||||||
что ЖК угнетают в печени продукцию тригли- |
лятора клеточного цикла циклина D1 и активато- |
|||||||||||||
церидов и ЛПОНП посредством активации сиг- |
ра апоптоза посредством прямого взаимодействия |
|||||||||||||
нальных путей тех же самых ядерных рецепторов. |
с митохондриями. В целом SHP может занимать |
|||||||||||||
Сигнальная роль желчных кислот помимо их вли- |
важное место в регуляции клеточного роста и |
|||||||||||||
яния в рамках кишечно-печеночной оси проявля- |
апоптоза и, следовательно, в определении ини- |
|||||||||||||
ется и тем, что они на системной основе повышают |
циации развития гепатоцеллюлярной карциномы. |
|||||||||||||
расход энергии посредством повышения содер- |
Не исключено, что SHP и другие ЯР окажутся |
|||||||||||||
жания трийодтиронина в бурой жировой ткани и |
ключом к пониманию ожирения как фактора |
|||||||||||||
скелетных мышцах. Тем самым они препятствуют |
риска различных форм рака органов желудочно- |
|||||||||||||
развитию ожирения и тканевой резистентности |
кишечного тракта и печени. |
|
|
|
|
|
||||||||
к инсулину (М. Watanabe и соавт., 2006). Этот |
|
|
|
|
|
|
|
|
|
|
|
|||
эффект опосредуется ядерным рецептором TGR5, |
Метаболизм, воспаление и фиброз |
|||||||||||||
сопряженным с G-белком, который, активируясь |
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
||||
под влиянием желчных кислот, стимулирует про- |
Печень играет ведущую роль в поддержании |
|||||||||||||
дукцию циклического АМФ (цАМР) и активность |
энергетического гомеостаза, особенно в ситуациях |
|||||||||||||
энзима дейодиназы 2-го типа. |
|
с резко меняющейся доступностью нутриентов. |
||||||||||||
Приведенные наблюдения составляют прямые |
Она синтезирует гликоген и жирные кислоты в |
|||||||||||||
доказательства, что желчные кислоты выполня- |
периоды нутритивной обеспеченности и произво- |
|||||||||||||
|
|
|
|
|
|
|
РЖГГК он-лайн – www.gastro-j.ru |
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
||

3, |
2010 |
|
|
|
|
|
Редакционная |
|
|
|
|
|
|
|
|
дит глюкозу и кетоновые тела в межпищевари- |
прессии маркёров активации и восстановлением |
||||||
тельные периоды и при голодании. |
|
способности аккумулировать ретиниловые эфиры |
|||||
Факторы роста фибробластов, FGF15 и FGF21, |
(M. Pinzani, 2009). |
|
|||||
характеризуются атипизмом в том отношении, что |
На экспериментальных моделях показано, что |
||||||
они функционируют как эндокринные гормоны и |
введение синтетических лигандов PPARγ, таких |
||||||
регулируют активность печени в зависимости от |
как тиазолидиндионы, приводит к угнетению |
||||||
нутритивного статуса. Образование FGF21 инду- |
фиброгенной активности стеллатных клеток, но |
||||||
цируется в печени в межпищеварительный период |
только в том случае, если лиганд вводился одно- |
||||||
пероксисомным |
пролифератор-активируемым |
временно с причинным фактором повреждения |
|||||
рецептором α (PPARα). В свою очередь, FGF21 |
печени. Наоборот, эффекты влияния синтети- |
||||||
стимулирует в печени окисление жирных кислот, |
ческих лигандов PPARγ на сформировавшийся |
||||||
кетогенез и глюконеогенез. При блокаде продук- |
фиброз оказались очень скромными или отсутс- |
||||||
ции FGF21 существенно уменьшается поток суб- |
твовали вовсе. Данный факт подтверждает нали- |
||||||
стратов по этим метаболическим путям. FGF21 |
чие низкой остаточной экспрессии указанного |
||||||
угнетает рост клеток и тканей, подавляет жен- |
ядерного рецептора в активированных фиброген- |
||||||
скую репродуктивную функцию и способствует |
ных клетках и подчеркивает возможность огра- |
||||||
развитию торпора (органическая и психическая |
ниченного применения этого класса препаратов в |
||||||
анергия) на фоне кратковременной гипотермии. |
качестве антифибротических средств в клиничес- |
||||||
В целом регуляторно-метаболический путь с |
кой практике. |
|
|
||||
участием PPARα – FGF21 играет важную роль |
Еще один ЯР – FXR – в последнее время при- |
||||||
в инициации и координации системного ответа |
влекает пристальное внимание исследователей. |
||||||
на голодание. Вместе с тем родственный гормон, |
FXR имеет прямое отношение к секреции желчи, |
||||||
FGF15, выступает как важный постпрандиальный |
метаболизму липидов и углеводов. В эксперимен- |
||||||
гормон. Синтез FGF15 индуцируется в тонкой |
те показано, что стимуляция стеллатных клеток |
||||||
кишке ЯР желчных кислот FXR. Далее FGF15 |
синтетическими агонистами FXR сопровождает- |
||||||
|
|
|
-VESTI |
|
|
||
поступает в системную циркуляцию и действует |
ся рядом.RUантифиброгенных реакций, включая |
||||||
как фактор, угнетающий синтез ЖК и стимули- |
восстановление экспрессии и функции PPARγ. |
||||||
рующий наполнение желчного пузыря желчью. |
В отличие от PPARγ экспрессия FXR значительно |
||||||
Одновременно FGF15 ингибирует в печени окис- |
возрастает в период активации стеллатных клеток, |
||||||
ление желчных кислот и глюконеогенез. В целом |
что позволяет рассматривать путь регуляции FXR |
||||||
можно считать, что FGF21 и FGF15 реципрокно |
как |
потенциальную мишень предупреждения и |
|||||
|
|
|
.M |
лечения фиброза печени. Подтверждением этому |
|||
регулируют липидный и углеводный метаболизм |
|||||||
в печени в ответ на изменения нутритивного ста- |
служит то, что в человеческих активированных |
||||||
туса (S. Kliewer, D. Mandelsdorf, 2009). |
|
стеллатных клетках экспрессия FXR очень низка |
|||||
Биология ЯР стеллатных клеток – ключевых |
или полностью заблокирована. |
||||||
эффекторных клеток в фиброгенном ответе пече- |
Печень постоянно или эпизодически подвер- |
||||||
ни на хроническое повреждение представляет |
жена воздействию химических агентов, таких |
||||||
собой важную область исследований. Обращает |
как |
ацетаминофен (парацетамол) и алкоголь. |
|||||
на себя внимание варьирующаяWWWэкспрессия ЯР. |
В экспериментах на «нокаутных» животных |
||||||
Последние могут экспрессироваться как в стел- |
показано, что цитохром Р450 и некоторые ЯР |
||||||
латных клетках в здоровой печени, так и в |
определяют масштаб и глубину повреждений |
||||||
активированных миофибробласт-подобных стел- |
печени после воздействия различных химических |
||||||
латных клетках в случае активного фиброгенеза |
веществ. Например, выключение гена цитохро- |
||||||
в поврежденном органе. Прототипом такой вариа |
ма |
Р4502е1 повышает резистентность печени |
|||||
бельности служит ядерный рецептор PPARγ, |
к токсическим |
эффектам |
четыреххлористого |
||||
экспрессия которого последовательно уменьша- |
углерода (CCl4), ацетаминофена и других низко- |
||||||
ется в процессе активации стеллатных клеток. |
молекулярных токсических соединений, многие |
||||||
Угнетение экспрессии PPARγ ассоциировано с |
из которых обнаруживаются в индустриальных |
||||||
нарастанием профиброгенного действия активи- |
стоках. CYP2Е1 служит |
ключевым фактором |
|||||
рованных стеллатных клеток в связи с повышен- |
в метаболической трансформации этих компо- |
||||||
ной активностью транскрипционных факторов, |
нентов в активные электрофильные соединения, |
||||||
например АР-1. Данный фактор регулирует клю- |
которые ковалентно связываются с клеточными |
||||||
чевые профиброгенные гены – проколлаген 1 и |
макромолекулами. Выключение экспрессии ЯР |
||||||
TIMP-1. Более того, угнетение активности PPARγ |
PPARb повышает чувствительность печени к воз- |
||||||
коррелирует с |
переходом стеллатных клеток |
действиям CCl4 |
и азоксиметана, а выключение |
||||
из «адипогенного» в «фиброгенный» фенотип. |
PPARα делает животных более восприимчивыми |
||||||
В соответствии с изложенным индуцируемая de |
к хроническому алкоголь-индуцированному пов- |
||||||
novo экспрессия PPARγ в активированных стел- |
реждению печени и ускоряет развитие цирроза. |
||||||
латных клетках сопровождается угнетением экс- |
Активация ядерного рецептора PXR защищает |
||||||
РЖГГК он-лайн – www.gastro-j.ru |
|
|


3, 2010 |
Лекции и обзоры |
|
|
УДК 612.35.08
Гастроинтестинальные миофибробласты – роль в регуляции физиологической активности и репарации желудочно-кишечного тракта
Э.Ф. Баринов, О.Н. Сулаева
(Донецкий национальный медицинский университет им. М. Горького)
Gastrointestinal myofibroblasts: role in regulation of physiological activity |
|
and reparation of gastro-intestinal tract |
RU |
E.F.Barinov, O.N.Sulayeva |
. |
|
|
Цель обзора. Проанализировать органоспеци- |
The aim of review. To analyze organ-specific |
фические особенности гастроинтестинальных мио- |
features of gastrointestinal myofibroblasts, functional |
фибробластов, функциональные свойства, роль в |
properties, role in embryogenesis and healing of ulcers |
эмбриогенезе и заживлении язвенных дефектов-VESTIof gastro-intestinal tract (GIT). |
|
стенки желудочно-кишечного тракта (ЖКТ). |
Original positions. Morphology, markers, origin |
Основные положения. Обсуждается морфоло.M- |
and stimulators of development of myofibroblasts, their |
гия, маркёры, источники и стимуляторы формирова- |
involvementinformationofmicronicheforepithelialstem |
ния миофибробластов, их участие в формировании |
cells, kinetics of tegmental epithelium, angiogenesis, |
микрониши для эпителиальных стволовых клеток, |
control of vascular permeability is discussed. These |
кинетике покровного эпителия, ангиогенезе, конт- |
functions are related to wide spectrum of myofibroblast |
роле сосудистой проницаемости. Эти функции свя- |
secretory products, producing molecules of basal |
заны с широким спектром секреторныхWWWпродуктов |
membrane, components of matrix and growth factors. |
миофибробластов, образующих молекулы базаль- |
Intestinal myofibroblasts are pacemakers for smooth |
ной мембраны, компоненты матрикса и факторы |
myocytes of GIT, they modulate neurotransmission, |
роста. Интестинальные миофиброласты являются |
participate in modulation of immune response, playing |
пейсмеккерами для гладких миоцитов ЖКТ, моду- |
a critical role in reparation of erosions and ulcers of |
лируют нейротрансмиссию, участвуют в модуляции |
GIT. |
иммунного ответа, играя критическую роль в репа- |
Key words: myofibroblasts, gastro-intestinal tract, |
рации эрозий и язв ЖКТ. |
ulcers, erosions, secretary products of myofibroblasts. |
Ключевые слова: миофибробласты, желудоч- |
|
но-кишечный тракт, язвы, эрозии, секреторные про- |
|
дукты миофибробластов. |
|
Баринов Эдуард Федорович – доктор медицинских наук, профессор, академик АН ВШ Украины, зав. кафедрой гистологии, цитологии и эмбриологии Донецкого национального медицинского университета. Контактная информация для переписки: barinoff@dsmu.edu.ua; Украина, 83003, г. Донецк, пр. Ильича, 16, Донецкий НМУ
Сулаева Оксана Николаевна – кандидат медицинских наук, доцент кафедры гистологии, цитологии и эмбриологии Донецкого национального медицинского университета
РЖГГК он-лайн – www.gastro-j.ru |
|

Лекции и обзоры |
|
|
|
|
3, 2010 |
|
|
|
|||||
аживление дефектов стенки желудочно- |
ми используют антитела против c-kit, экспрессию |
|||||
кишечного тракта (ЖКТ) – эрозий, язв |
которого связывают с высокой чувствительностью |
|||||
З– представляет собой сложный процесс, |
данной популяции клеток к фактору стволовых |
|||||
направленный на ликвидацию повреждения. Его |
клеток – СК (SCF) [34]. Для идентифика- |
|||||
хронология и конечный исход зависят от эффек- |
ции миофибробластов используются также менее |
|||||
тивности реализации последовательных фаз, |
известные, но специфические молекулы клеточ- |
|||||
включая альтерацию и сосудистую реакцию, |
ной адгезии – ОВ-кадгерин и Thy-1 [26]. |
|||||
воспаление, ангиогенез, пролиферацию и диф- |
Морфологически миофибробласты представ- |
|||||
ференцировку клеток с закрытием дефекта гра- |
ляют собой звездчатые клетки с активным ядром. |
|||||
нуляционной тканью, контракцию краев дефекта |
В цитоплазме развиты гранулярная эндоплазма- |
|||||
и эпителиальную реституцию, завершающуюся |
тическая сеть и комплекс Гольджи. Уникальной |
|||||
ремоделированием внеклеточного матрикса с при- |
особенностью является степень развития и орга- |
|||||
обретением дефинитивного паттерна межткане- |
низация цитоскелета, представленного пучками |
|||||
вых отношений [3, 10, 30]. При этом репаратив- |
параллельно |
расположенных микрофиламентов, |
||||
ные процессы, с одной стороны, жестко связаны |
получивших название стрессорных волокон. В их |
|||||
с характером течения воспаления, а с другой, |
составе помимо цитоплазматического β-актина |
|||||
детерминируют сроки восстановления структур- |
присутствует также α-SМА, количество которо- |
|||||
ного гомеостаза и барьерных свойств слизистой |
го прямо пропорционально локальному уровню |
|||||
оболочки (СО) [5, 34]. |
|
трансформирующего фактора роста (TGFβ) |
||||
Несмотря на то, что репаративную регенера- |
[11]. Подобно микрофиламентам гладких миоци- |
|||||
цию при ульцерогенезе считают стереотипным |
тов, стрессорные волокна фиксируются к плот- |
|||||
процессом, описанным в многочисленных отечес- |
ным тельцам цитоплазмы и плазмолеммы [21, |
|||||
твенных и зарубежных публикациях [2, 7, 10], |
35]. |
|
|
|
||
на сегодняшний день, по-прежнему, ряд вопросов |
Характерно, что миофибробласт работает не |
|||||
остается открытым. Например, неясно, наруше- |
как отдельная независимая единица, а как часть |
|||||
ние работы каких клеток играет ведущую роль в |
системы., RUсопряженная с другими клетками и |
|||||
детерминации неэффективной репарации тканей, |
элементами матрикса. Это связано с особеннос- |
|||||
заведомо |
обладающих высоким регенераторным |
тями контактов миофибробластов между собой |
||||
потенциалом. Среди прочих причин ключевым |
и с внеклеточным матриксом (ВКМ). Так, |
|||||
фактором |
дизрегенерации считается |
нарушение |
отростки соседних клеток связаны между собой |
|||
процесса |
формирования грануляционной ткани |
с помощью щелевидных соединений, что обес- |
||||
|
|
-VESTI |
|
|
||
[11]. Ее развитие характеризуется не только |
печивает формирование единой сети [20, 21]. |
|||||
образованием новых клеточных коопераций, но и |
Кроме |
того, |
миофибробласты активно регис- |
|||
|
|
.M |
трируют сигналы из внеклеточной среды пос- |
|||
появлением уникальной клеточной линии – мио- |
||||||
фибробластов, считающихся главными эффек- |
редством специализированных и уникальных по |
|||||
торными клетками грануляций и стимуляторами |
своей организации контактов – фибронексусов. |
|||||
эпителиальной реституции [9, 15, 32]. |
|
В условиях in vitro аналогами этих контактов |
||||
Морфологически миофибробласты были иден- |
являются крупные суперзрелые зоны фокальной |
|||||
тифицированы более 100 лет WWWназад. Но лишь |
адгезии (supermature focal adhesions). На молеку- |
|||||
недавно установлена роль этого семейства клеток |
лярном уровне эти участки соответствуют зонам |
|||||
в паракринной регуляции фундаментальных био- |
экспрессии ОВ-кадгерина (известного также как |
|||||
логических процессов [10, 20]. До сих пор мало |
кадгерин-11) – трансмембранного рецептора адге- |
|||||
известно |
об органоспецифических особенностях |
зии, связанного с цитоплазматической стороны с |
||||
биологии |
гастроинтестинальных миофиброблас- |
пучками микрофиламентов [26]. Протяженность |
||||
тов, возможностях регуляции количества и актив- |
таких зон адгезии зависит от ригидности ВКМ |
|||||
ности этой клеточной линии при заживлении |
и, в свою очередь, лимитирует силу натяжения, |
|||||
язвенных дефектов СО, что и определило цель |
генерируемую стрессорными волокнами, фик- |
|||||
настоящего обзора. |
|
сированными к плотным тельцам [31]. |
||||
|
|
|
Изменение давления и химического состава |
|||
Биология миофибробластов |
|
ВКМ |
(например, при ферментной деградации |
|||
|
плазминогеном) ведет к активации механо-сенси- |
|||||
|
|
|
||||
К семейству миофибробластов традиционно |
тивных ионных каналов [21]. Это сопровождается |
|||||
относят разновидность стромальных клеток мезен- |
входом Са2+, что вызывает деполяризацию плаз- |
|||||
химного происхождения, классическим маркёром |
молеммы миофибробластов, сокращение и пере- |
|||||
которых |
является α-актин гладких |
миоцитов |
дачу сигналов по сети миофибробластов. Кроме |
|||
(α-SМА) [20]. Кроме того, миофибробласты экс- |
того Са2+ является универсальным мессенджером, |
|||||
прессируют мышечный миозин, что определяет |
активирующим такие трансдукторы, как проте- |
|||||
контрактильные свойства этих клеток. Для диф- |
инкиназа С и р38-МАРкиназа [25]. Последняя |
|||||
ференциальной диагностики с гладкими миоцита- |
активируется |
также при стимуляции интегрин- |
||||
10 |
РЖГГК он-лайн – www.gastro-j.ru |