

намного превышает физическое испарение, и подавляющая часть влаги поступает от подстилающей поверхности в атмосферу через транспирацию. Так, в экваториальных лесах Малайзии годовая величина транспирации составляет 1350 мм, а испарение с поверхности почвы — всего лишь 25 мм. Только через транспирацию «дождевых» экваториально-тропических лесов в атмосферу поступает 62% влаги, испаряющейся с суши. Если же учесть возврат осадков, перехватываемых кронами деревьев, то в целом биота обеспечивает не менее 70 — 80% внутреннего оборота влаги между атмосферой и остальными блоками наземных геосистем. Растительность прямо или косвенно способствует уменьшению выходного потока влаги путем сокращения поверхностного стока; при наличии мощной подстилки из растительных остатков поверхностный сток может практически прекращаться.
Биогенный оборот веществ
Биогеохимический цикл, или «малый биологический круговорот»,— одно из главных звеньев функционирования геосистем. В основе его — продукционный процесс, т. е. образование органического вещества первичными продуцентами — зелеными растениями, которые извлекают двуокись углерода из атмосферы, зольные элементы и азот — с водными растворами из почвы. Около половины создаваемого при фотосинтезе органического вещества (брутто-продукции) окисляется до СО2 при дыхании и возвращается в атмосферу. Оставшаяся (за вычетом затрат на дыхание) фитомасса называется чистой первичной продукцией. Часть ее поступает в трофическую цепочку — потребляется растительноядными животными (фитофагами); следующий трофический уровень представлен плотоядными животными (зоофагами) .
При переходе от одного трофического уровня к другому отношение биомасс уменьшается на 2 — 3 порядка, т. е. в n: 100— n: 1000 раз. Особенно большой разрыв наблюдается в лесах, где основная масса гетеротрофных организмов приходится на сапрофагов. Согласно Р. Уиттекеру, в злаковниках животные в среднем потребляют 10 — 15% чистой первичной продукции, в лесах — 4 — 7%, в пустынях и тундрах — 2 — 3%. Эффективность роста, т. е. отношение вторичной продукции к потреблению, в сообществах злаковников и пустынь составляет примерно 15%, а у всех остальных сообществ суши — 10%. В результате получается, что вторичная продукция на суше составляет менее 1% от первичной, за исключением степей и саванн (табл. 8) .
Основная часть фитомассы после отмирания разрушается животнымисапрофагами, бактериями, грибами, актиномицетами. В конечном счете мертвые органические остатки минерализуются микроорганизмами (в меньшей степени путем абиотического окисления). Конечные продукты минерализации возвращаются в атмосферу
173
Т а б л и ц а 8. Соотношение первичной и вторичной продуктивности
в разных сообществах (по Р. Уиттекеру)
Сообщества |
Фито- |
Чистая |
Потреб- |
Вторич- |
Биомас- |
|
|
масса, |
ная |
са жи- |
|||
|
первич- |
ление |
||||
|
т/га |
продук- |
вотных, |
|||
|
ная про- |
живот- |
||||
|
|
ция, |
кг/га |
|||
|
|
дукция, |
ными, |
|||
|
|
кг/га x |
|
|||
|
|
т/га год |
т/га год |
|
||
|
|
x год |
|
|||
|
|
|
|
|
||
Влажные тропические леса |
450 |
22 |
1,53 |
152,9 |
194 |
|
Тропические сезонные листопадные |
350 |
16 |
|
|
107 |
|
леса |
0,96 |
96,0 |
||||
|
|
|
||||
Вечнозеленые леса умеренного |
350 |
13 |
|
|
100 |
|
пояса |
0,52 |
52,0 |
||||
|
|
|
||||
Листопадные леса умеренного |
300 |
12 |
|
|
157 |
|
пояса |
0,60 |
60,0 |
||||
|
|
|
||||
Тайга |
200 |
8 |
0,32 |
31,7 |
48 |
|
Саванны |
40 |
9 |
1,33 |
200,0 |
147 |
|
Степи умеренного пояса |
16 |
6 |
0,60 |
88,9 |
67 |
|
Тундра |
6 |
1,4 |
0,04 |
3,8 |
4 |
|
Пустыни и полупустыни с кустар- |
|
|
|
|
|
|
никами |
7 |
0,9 |
0,27 |
3,9 |
4 |
|
Настоящие пустыни; полярные |
|
|
|
|
|
|
зоны |
0,2 |
0,03 |
0,00008 |
0,008 |
0,008 |
|
Болота и марши |
150 |
20 |
1,6 |
160 |
100 |
(CO2 и другие летучие соединения) и в почву (зольные элементы и азот). Процессы созидания и разрушения биомассы не всегда сбалансированы — часть ее (в среднем менее 1%) может выпадать из круговорота на более или менее длительное время и аккумулироваться в почве (в виде гумуса) и в осадочных породах.
Биологический метаболизм характеризуется многочисленными показателями, в том числе относящимися к внутренним обменным процессам в самом биоценозе — между его различными трофическими уровнями, таксономическими группами, зелеными (фотосинтезирующими), нефотосинтезирующими надземными и подземными частями фитоценоза и др. Для географа первостепенный интерес представляют взаимоотношения биоценоза как целого с другими блоками геосистемы, зависимость биогенных потоков и биологической продуктивности от географических факторов, закономерности их проявления на региональном и локальном уровнях, степень замкнутости или открытости биологического круговорота и его роль во внутреннем механизме функционирования ландшафта и его внешних связях.
С этой точки зрения, важнейшие показатели биогенного звена функционирования — запасы фитомассы и величина годичной первичной продукции, а также количество опада и аккумулируемого мертвого органического вещества. Для оценки интенсивности круговорота используются производные показатели: отношение чистой первичной продукции к запасам фитомассы, отношение живой фито-массы к мертвому органическому веществу и др. Для характеристи-
174
ки вклада биоты в функционирование геосистем особенно важны биогеохимические показатели: количество элементов питания, потребляемых для создания первичной биологической продукции (емкость биологического круговорота) и их химический состав, возврат элементов с опадом и закрепление в истинном приросте, накопление в подстилке, потеря на выходе из геосистемы и степень компенсации на входе.
В табл. 9 — 11 приведены основные показатели биологического звена функционирования ландшафтов по разным источникам.
Т а б л и ц а 9. Запасы и продуктивность фитомассы
плакорных сообществ различных зон и подзон
Зоны (нодзоны) |
Фнтомасса, |
Продукция, |
|
т/га |
т/га год |
||
|
|||
Полярные пустыни |
1,6 |
0,2 |
|
Арктическая тундра |
5 |
1 |
|
Субарктическая тундра |
25 |
3 |
|
Лесотундра |
60 |
4 |
|
Северная тайга (темнохвойная) |
125 |
5 |
|
Средняя тайга (темнохвойная) |
250 |
6,5 |
|
Средняя тайга (лиственничная) |
150 |
4 |
|
Южная тайга (темнохвойная) |
300 |
8 |
|
Подтайга восточноевропейская |
300 |
12 |
|
Подтайга западносибирская |
220 |
12 |
|
Широколиственные леса западноевропейские |
380. |
13 |
|
Широколиственные леса восточно- |
|
|
|
европейские |
350 |
12 |
|
Широколиственные леса новозеландские |
400 |
15 |
|
Суббореальные притихоокеанские леса |
|
11 — 16 |
|
из дугласии |
> 1000 (до 2900) |
||
|
|||
Луговые степи европейско-сибирские |
17 |
19 |
|
Типичные суббореальные степи |
10 — 13 |
10 — 13 |
|
Сухие суббореальные степи |
6 |
5 |
|
Пустыни суббореальные (полынно- |
4 |
|
|
солянковые) |
1,2 |
||
|
|||
Пустыни тропические |
1,5 |
0,5 |
|
Влажные субтропические леса |
450 |
24 |
|
Субтропические секвойевые леса |
> 1000 (до 4250) |
До 27 |
|
Саванны типичные |
40 |
12 |
|
Сезонно-влажные саванновые леса |
200 |
16 |
|
Влажные экваториальные леса |
500 |
30 — 40 |
Продуктивность биоты определяется как географическими факторами, так и биологическими особенностями различных видов. Наибольшими запасами фитомассы характеризуется лесная растительность, эдификаторы которой способны накапливать живое вещество в течение многих десятилетий и даже столетий. Максимальные запасы присущи лесам из долго живущей секвойи вечнозеленой. Однако при наличии отдельных специфических видов, составляющих исключения, общая закономерность состоит в том, что у аналогич-
175
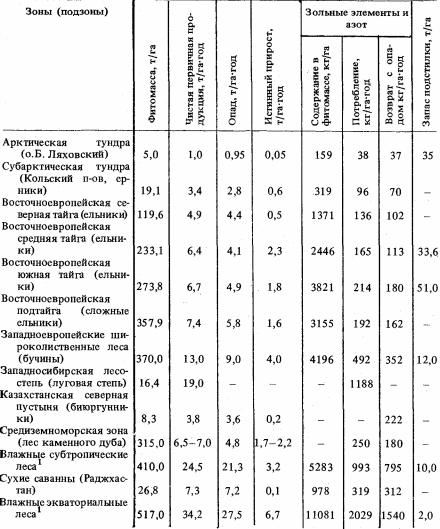
Т а б л и ц а 10. Биогеохимический круговорот в плакорных сообществах
различных ландшафтных зон
1 Осредненные данные, по Л.Е. Родину и Н.И. Базилевич.
ных жизненных форм (древесных, травянистых и др.) запасы биомассы тем больше, чем выше теплообеспеченность и чем ближе к оптимуму соотношение тепла и влаги.
В величине ежегодной биологической продукции региональные
176
Т а б л и ц а 11. Основные показатели структуры и функционирования
некоторых зональных сообществ (по Н. И. Базилевич)
Показатель |
Тундра |
Ельник- |
Дубрава |
Луговая |
Белосак- |
|
|
типичная |
кисличник |
осоково- |
злаково- |
саульник |
|
|
кустар- |
южнота- |
снытьевая |
разно- |
илаковыи |
|
|
ничково- |
ежный |
|
травная |
|
|
|
моховая |
|
|
степь |
|
|
Живое органическое ве- |
|
|
|
|
|
|
щество, т/га |
16,88 |
344,26 |
491,49 |
16,01 |
8,90 |
|
Фитомасса, т/га |
16,27 |
343,48 |
490,62 |
14,88 |
8,73 |
|
В том числе: |
|
5,6 |
1,0 |
21,4 |
8,2 |
|
зеленые части, % |
40,6 |
|||||
|
|
|
|
|||
многолетние надземные |
|
|
|
|
|
|
части, % |
2,2 |
70,2 |
78,8 |
- |
35,1 |
|
подземные части, % |
57,2 |
24,2 |
20,2 |
78,6 |
56,7 |
|
Животные, т/га |
0,012 |
0,08 |
0,17 |
0,20 |
0,02 |
|
В том числе: |
|
|
5,8 |
6,0 |
20,0 |
|
фитофаги, % |
4,0 |
5,0 |
||||
|
|
|
||||
сапрофаги, % |
95,0 |
94,0 |
96,6 |
93,0 |
75,0 |
|
зоофаги, % |
1,0 |
1,0 |
0,6 |
1,0 |
5,0 |
|
Чистая первичная |
|
|
|
|
|
|
продукция, т/ra'год |
3,44 |
16,35 |
16,79 |
20,75 |
2,78 |
|
Отношение первичной |
|
|
|
|
|
|
продукции к живой |
|
|
|
|
|
|
фитомассе |
0,20 |
0,05 |
0,03 |
1,40 |
0,30 |
|
Мертвое органическое |
|
|
|
|
|
|
вещество, т/га |
152,80 |
193,00 |
395,16 |
622,99 |
25,12 |
|
в том числе почвенный |
|
|
|
|
|
|
гумус, % |
46,0 |
41,0 |
70,0 |
96,0 |
99,5 |
|
Емкость биологического |
|
|
|
|
|
|
круговорота, кг/га.год |
115,0 |
470,0 |
600,0 |
1115,0 |
126,0 |
|
Среднее содержание |
|
|
|
1,21 |
0,90 |
|
в продукции N, % |
1,00 |
0,70 |
0,85 |
|||
|
|
|||||
Среднее содержание |
|
|
|
|
|
|
в продукции зольных |
|
|
|
|
|
|
элементов, % |
2,18 |
2,10 |
2,68 |
4,44 |
3,78 |
|
Химизм потребляемых |
N (Ca, K) |
N, Ca, K |
Ca, N, K |
Si, N, K или |
|
|
элементов |
Ca |
Ca, K, N, Mg |
||||
|
|
|
||||
Потребление продукции |
|
|
|
|
|
|
фитофагами, % |
2,0 |
2,0 |
5,0 |
5,6 |
12,0 |
|
Абиотические потоки |
|
|
|
|
|
|
органического вещества, |
|
|
|
|
|
|
кг/га: |
|
|
|
|
|
|
вход |
6,4 |
15,0 |
10,0 |
10,0 |
2,0 |
|
выход |
122,0 |
365,0 |
9,0 |
1,4 |
30,0 |
и локальные географические закономерности, включая зональность, секторность, высотную поясность и внутриландшафтную морфологическую дифференциацию, проявляются более четко. При достаточном количестве влаги продуктивность возрастает от высоких широт к низким в соответствии с ростом энергообеспеченности. В одинаковых термических условиях наибольшая продуктивность наблюдается
177
при оптимальном соотношении тепла и влаги. В сравнимых (плакорных) местоположениях максимальная биологическая продуктивность присуща экваториальным ландшафтам (см. табл. 9), заметно меньше она во влажных (лесных) субтропиках. Среди суббореальных ландшафтов наивысшей продуктивностью выделяются луговые степи, им несколько уступают широколиственные леса. Самая низкая продуктивность присуща ландшафтам с резким дефицитом тепла (полярным) или влаги (пустынным).
Отношение первичной продуктивности к запасам фитомассы наивысшее в травяных сообществах, у которых нет многолетних надземных органов (в луговых степях до 1,4:1), наинизшее — в лесных (0,03 — 0,06: 1) .
Значительная часть ежегодной продукции отмирает и разрушается — попадает в деструкционный цикл, меньшая часть закрепляется в приросте (см. табл. 10). Отмершее органическое вещество, как правило, не полностью минерализуется, аккумулируясь в разном количестве и разных формах в ландшафте. Скорость разрушения органической массы растет с увеличением притока солнечного тепла. При недостатке тепла ежегодный опад не успевает разрушаться и в ландшафте накапливается избыточная мортмасса. В экстрааридных ландшфатах с их высоким энергетическим потенциалом и незначительной продуктивностью интенсивность деструкции намного превышает продуцирование биомассы, и накопление мертвого органического вещества практически отсутствует. Продукционные и деструкционные процессы наиболее сбалансированы в условиях, близких к оптимуму по соотношению тепла и влаги. С увеличением тепло-обеспеченности основная часть органических остатков переходит в почвенный гумус. В луговых черноземных степях его запасы достигают 600 — 1000 т/га, в почвах широколиственных лесов — около 300, тогда как в таежных — около 100, а в тундровых — около 70 т/га. В тундре и тайге в составе мортмассы преобладают неразложившиеся растительные остатки — главным образом подстилка, а также сухостой, валежник, мертвые корни и др. Запасы подстилки в этих зонах достигают 40 — 50 т/га; в широколиственных лесах они сокращаются до 10 — 15, а в экваториальных — до 2 т/га. В степях наземная мортмасса представлена растительной ветошью (4 — 10 т/ra) .
Мертвое органическое вещество и запас биомассы в органах растений служат важным резервом питательных веществ, обеспечивающим устойчивость биоты к колебаниям внешней среды и позволяющим поддерживать биологический метаболизм в условиях интенсивного абиогенного выноса элементов зольного и азотного питания. Лесная растительность, как известно, существует в условиях избыточного увлажнения и интенсивного стока, ведущего к безвозвратной потере элементов питания; лесные почвы относительно бедны этими элементами. В такой обстановке запас элементов-органогенов в живом веществе и в мощной подстилке, прочно удерживающей необходимые элементы питания, обеспечивает определенную авто178
номность (высокую степень замкнутости) биологического круговорота.
В степях, где растительность не способна аккумулировать запасы живой фитомассы, а опад быстро разрушается, резерв минерального питания сосредоточен в почвенном гумусе.
Одним из показателей скорости трансформации органического вещества может служить отношение годичной первичной продукции к запасам мертвых растительных остатков (без почвенного гумуса): в тундре — 0,02, в лесах — 0,15, в луговых степях — 0,9, в пустынях — 25 и более.
С величиной первичной биологической продуктивности непосредственно связана емкость биологического круговорота веществ. Хотя количество вовлекаемого в оборот минерального вещества зависит от биологических особенностей различных видов, размещение этих видов в значительной мере подчинено географическим закономерностям. Так, хвойные деревья ассимилируют меньше зольных элементов и азота, чем лиственные, а последние — меньше, чем травянистая растительность. Растения аридных областей используют для создания органической массы больше химических элементов, чем растения гумидных областей. Наименьшая зольность у мхов (2— 4% от сухого вещества), наибольшая — у галофитов (до 25%). Зольность хвои и листьев деревьев — 3 — 4%, древесины хвойных— 0,4, лиственных —
0,5, злаков — 6 — 10%.
Сочетание всех факторов приводит к тому, что самая низкая емкость биологического круговорота свойственна полярным и тундровым ландшафтам (см. табл. 10, 11), хотя продуктивность у них выше, чем в пустынях. В бореальных и особенно суббореальных лесах в оборот вовлекается значительно больше зольных элементов и азота, но максимальная емкость круговорота среди плакорных суббореальных сообществ присуща луговым степям. На первом месте в зональном ряду по количеству вовлекаемого в круговорот минерального вещества стоят влажные экваториальные леса.
Что касается элементарного химического состава вещества, участвующего в биологическом метаболизме, то основную его часть составляют важнейшие элементы-биогены, главным образом N, К, Са, Si, затем Р, Mg, S, Fe, Al и др. В зависимости от избирательной способности растений к поглощению тех или иных элементов, их количественные соотношения в составе биомассы и ежегодно потребляемого минерального вещества несколько варьируют, обнаруживая также определенные географические закономерности — прежде всего зональность. Так, тундровые сообщества потребляют больше всего азота, затем следуют кальций и калий; в минеральном метаболизме тайги на первом месте также стоит азот, много потребляется кальция, на третьем месте
— калий; в широколиственных лесах — на первом месте кальций, затем азот и калий, в степях основные элементы идут в следующем порядке: кремний, азот, калий, кальций, в пустынях — кальций, калий, азот, магний; в тропических и эквато-
179
риальных лесных ландшафтах особенно активно поглощаются кремний, железо, алюминий.
До сих пор речь шла о минеральном обмене растительности с почвой, но следует вспомнить, что основная часть живого вещества строится из элементов, которые ассимилируются в результате газо-обмена растений с атмосферой. Важную роль играет углеродный обмен, с которым связаны биогенная трансформация солнечной энергии, баланс СО2 в геосистемах и ее дальнейшая миграция, от чего, в частности, зависят характер обменных процессов в поглощающем комплексе почв, химизм вод речного стока и т. д.
Количество ассимилируемого углерода в десятки раз превышает величину потребляемых зольных элементов и азота. Ежегодное потребление СО2 тундровой растительностью составляет около 10 т/га, южнотаежными и широколиственными лесами — около 35, луговыми степями — около 50, влажными экваториальными лесами — около 130 т/га. В большинстве сообществ (особенно в лесных) свыше половины ассимилированного углерода возвращается в атмосферу при дыхании организмов (в основном растений, а также микроорганизмов; доля животных составляет 1 — 3%). Остальная часть содержащегося в ежегодной продукции углерода высвобождается при разложении органических остатков и также возвращается в атмосферу в виде СО2, относительно небольшое количество растворяется в воде, образуя угольную кислоту, которая диссоциируется на ионы НСО3- и Н+. В среднем поглощение и выделение углерода балансируются в годовом цикле, но небольшая его доля (менее 1%) выводится из биологического круговорота — аккумулируется в почве, выносится со стоком.
Связывание СО2 в процессе фотосинтеза сопровождается выделением свободного кислорода. При дыхании и разложении органических остатков кислород потребляется на окисление органического вещества до СО2. Но поскольку часть мертвой органической массы выпадает из деструкционного цикла, поступление кислорода в атмосферу в результате биологического метаболизма превышает его потребление. Согласно Н. И. Базилевич, это превышение составляет в дубравах 6,16 т/га год, в южнотаежных ельниках — 3,7, в бело-саксаульниках — 0,65, в тундре — 0,44 т/га год. В луговой степи синтез и разложение сбалансированы и биологический обмен не влияет на кислородный баланс. Если же исключить накопление биомассы в приросте и учитывать только необратимые потери органического вещества (его внешние выходные потоки — со стоком, ветром), то на первом месте (в пределах территории СССР) по накоплению кислорода будут ельники, затем тундра; вклад дубрав и луговых степей ничтожен.
Все изложенные закономерности основаны на достаточно широких региональных сравнениях по материалам для плакорных местоположений и соответствующих им сообществ. Между тем в характере биологического круговорота и продуцировании биомассы наблюдаются существенные внутриландшафтные различия между пла-
180
корными (автономными, элювиальными) и подчиненными (аккумулятивными, преимущественно гидроморфными) фациями. При недостаточном атмосферном увлажнении и высокой теплообеспеченности перераспределение влаги в ландшафте обусловливает большую контрастность в интенсивности биологического круговорота и продуцировании биомассы по местоположениям. В гидроморфных местоположениях, как правило, наблюдаются наиболее высокие показатели. Так, в гривисто-ложбинном лесостепном ландшафте Барабинской степи плакорные лугово-степные фации плоских вершин грив на обыкновенных черноземах продуцируют ежегодно 19 т/га фитомассы, а травяные болота межгривных депрессий на торфяноболотной слабосолончаковатой почве — свыше 60 т/га. Первые потребляют на создание фитомассы 1761 кг/ra год зольных элементов и азота, а вторые — 3831 (данные Н. И. Базилевич и А. А. Титляновой).
Резко выражена внутриландшафтная контрастность в указанном отношении в пустынях, где участки с ничтожной продуктивностью (такыры, солончаки) сочетаются с травяными тугаями (заросли тростника, рогоза и др.), дающими почти до 100 т/га первичной продукции.
Вусловиях избыточного атмосферного увлажнения и низкой теплообеспеченности дополнительное количество влаги за счет внутриландшафтного перераспределения атмосферных осадков мало влияет на биологическую продуктивность или даже косвенно ведет к ее снижению, поскольку переувлажнение ухудшает термический режим и аэрацию. По этим причинам в тундре заболоченные понижения отличаются наиболее замедленным биогенным оборотом веществ и пониженной продуктивностью.
Сдругой стороны, плакорные повышения сильнее подвержены воздействию ветра, сдуванию снега и выхолаживанию. Самыми благоприятными оказываются трансэлювиальные склоновые местоположения, где и наблюдается наибольшая для тундры биологическая продуктивность.
Втайге и широколиственных лесах внутриландшафтные различия продуктивности и интенсивности биогенного круговорота относительно невелики; в наилучших условиях находятся более теплые и дренированные местоположения, в наихудших — болота, дающие всего 2 — 4 т/га первичной продукции в год.
Степень замкнутости биологического круговорота также различна для автономных и подчиненных фаций. Первые в целом характеризуются высокой замкнутостью круговорота, т. е. незначительным участием внешних потоков органического вещества по сравнению с внутренними. Приток вещества извне (с атмосферными осадками, пылью), как правило, невелик, потеря его (со стоком, ветровым выносом) чаще превышает приток, но в разных зональных типах геосистем соотношения могут существенно дифференцироваться. По расчетам Н. И. Базилевич, в тундре входные потоки органического вещества составляют всего 0,2% от величины ежегодной первичной продукции, а выходные достигают около 4%, так что баланс вещества отрицательный и потеря его составляет 116 кг/га, или 3,4% от ежегодной продукции. В южнотаежных ельниках вынос (365 кг/га)
181
превышает привнос извне (15 кг/га) на 350 кг/га, т. е. составляет примерно 2% от первичной продукции. В дубравах входные и выходные потоки (10 и 9 кг/га соответственно) почти сбалансированы, а в луговой степи первые даже несколько превышают вторые (10 против 1,4 кг/га), так что положительное сальдо баланса составляет около 0,4% от первичной продукции. В белосаксаульниках потери органического вещества, главным образом из-за выдувания, равны 30 кг/га, приход же незначителен (2 кг/га), и в среднем за год теряется около 10% первичной продукции 1.
В подчиненных морфологических частях ландшафта входные и выходные потоки вещества могут быть во много раз больше, чем в плакорных. В гидроморфных фациях входные и выходные потоки обычно нескомпенсированы и возможна повышенная аккумуляция как органического, так и минерального вещества. Показательный пример сравнения плакорной лугово-степной фации на плоской вершине гривы и низинного травяного болота в межгривном понижении приводят Н. И. Базилевич и А. А. Титлянова
(табл. 12).
Т а б л и ц а 12. Круговорот элементов в двух фациях
западносибирской лесостепи (кг/га. год)
Звенья круговорота |
Углерод |
Азот |
Зольные элементы |
||
|
|
|
Всего |
Несоле- |
Соли |
|
|
|
вые |
|
|
|
|
|
|
|
|
Луговая степь на обыкновенных черноземах |
|
|
|||
Потребление на создание фитомассы |
7120 |
260 |
1501 |
- |
- |
Поступление с атмосферными осадками |
+ 37,7 |
+3,6 |
+ 132,1 |
+ 37,2 |
+ 94,9 |
Вынос с поверхностным стоком |
— 163,1 |
— 6,1 |
-216,6 |
-192,7 |
-23,9 |
Вынос с внутрипочвенным боковым стоком |
— 0,01 |
|
-0,1 |
-0,01 |
-0,1 |
Вынос с нисходящими растворами |
— 1,0 |
— 0,1 |
-10,0 |
0 |
-10,0 |
Баланс абиотических потоков |
— 126,4 |
— 2,6 |
-94,6 |
-155,5 |
+60,9 |
Травяное болото на торфяно-болотной слабосолончаковатой почве |
|
||||
Потребление на создание фитомассы |
31 340 |
585 |
3246 |
- |
- |
Поступление извне — всего с |
+451,6 |
+32,5 |
+ 6244,6 |
+615,2 |
+ 5629,4 |
атмосферными осадками |
+ 71,0 |
+7,0 |
+ 246,0 |
+ 70,0 |
+ 176,0 |
поверхностным стоком |
+ 301,0 |
+25,0 |
+827,0 |
+287,0 |
+ 540,0 |
внутрипочвенным боковым стоком |
-+-0,6 |
-+-0,03 |
+6,6 |
+0,2 |
+6,4 |
восходящими растворами |
+47,0 |
+-4,5 |
+ 2849,0 |
+147,0 |
+ 2702,0 |
потоком грунтовых вод |
+-32,0 |
+3,0 |
+ 2316,0 |
+ 111,0 |
+ 2205,0 |
Вынос — всего с |
— 54,9 |
— 2,6 |
-3021,0 |
-73,2 |
-2947,8 |
внутрипочвенным боковым стоком |
— 0,9 |
— 0,05 |
-5,0 |
-0,2 |
-4,8 |
нисходящими растворами |
— 51,0 |
— 1,6 |
-2842,0 |
-54,0 |
-2788,0 |
потоком грунтовых вод |
— 3,0 |
— 1,0 |
- 174,0 |
-19,0 |
-155,0 |
Баланс абиотических потоков |
+396,7 |
+29,9 |
+ 3223,6 |
+542,0 |
+ 2681,6 |
1 См.: Базилевич Н. И., Гребенщиков О. С., Тишков А. А. Географические закономерности структуры и функционирования экосистем. М., 1986. С. 297.
182