

Тема 7. Молекулярные основы эволюции
План
1. Элементарные эволюционирующие физико-химические структуры: белки и
нуклеиновые кислоты.
2. Проблема возникновения новых генов и их дальнейшей эволюции.
3. Основные тенденции в эволюции генов.
4. Теория нейтральности (М. Кимура).
5. «Недарвиновская эволюция».
6. Скорости эволюции генов и белков. Концепция молекулярных часов.
7. Использование митохондриальной ДНК в эволюционных исследованиях.
8. Построение филогенетических древ на основе "молекулярных часов".
Различие в структуре двух белков отражает их дивергенцию. Этот показатель различия между аминокислотами выражается в процентах. Как правило, путем определения дивергенции белков у разных видов обычно судят о сроках их расхождения. Таким способом составляют эволюционные «часы». С их помощью можно определять мутации, которые произошли в структуре данного белка. По результатам этих измерений определяется время появления новых генов. Дивергенция нуклеиновых кислот, естественно, отличается от дивергенции белков, которые синтезируются при их участии. Такое отличие, как было указано выше, обусловлено кодированием каждой аминокислоты тремя нуклеотидами.
Американские ученые Э. Цукеркандль и Л. Полинг, изучая молекулярные основы эволюции органического мира, доказали, что если третичная структура и функции определенного белка особей разных типов или классов остаются неизменными, то скорость эволюции его приблизительно постоянна и одинакова. Когда речь идет об эволюции белка, то подразумевается и эволюция гена. Скорость эволюции белка, как правило, измеряется числом аминокислотных замен в год на данную позицию в белке. Естественно, что эволюция различных белков и генов протекает с различной скоростью. Тем не менее, учитывая постоянство скорости замены аминокислот в составе белка, можно определить время появления того или иного вида, а также момент дивергенции рода, семейств, отряда, класса, типа. Так, построив родословную P-глобина, можно определить время, когда два ныне живущих вида имели общего предка. С помощью таких родословных установлено, что общий предок карпа и человека существовал около 400 млн. лет назад, ехидны и человека — 225 млн, собаки и человека — 70 млн. Результаты изучения эволюционных процессов на молекулярном уровне подтверждают правильность принципа дивергенции, который был выдвинут Ч. Дарвином.
Но мутации, происходящие в уникальных генах, могут привести к очень серьезным последствиям, даже к гибели организма. Очень важно также и то, какой нуклеотид в триплете претерпевает изменения при мутации. Так, например, триплет УУУ кодирует аминокислоту фенилаланин. Если в результате точковой мутации третий нуклеотид этого кодона заменяется аденином или гуанином, то кодоны УУА и УУГ включают в полипептидную цепь не фенилаланин, а лейцин, что приводит к существенному изменению молекулы белка. В результате таких мутаций ДНК человека оказалась гомологичней ДНК макаки на 66%, быка — на 28%, крысы — на 17%, лосося — на 8%, бактерии кишечной палочки— на 2%. Замена нуклеотидов истинного гена, выполняющего определенную функцию, приводит к закреплению данного изменения естественным отбором.
Замещение науклеотидов в «псевдогенах», то есть неактивных генах, не влияет на жизнеспособность организма, так как эти гены молчащие. Поэтому у них замена нуклеотидов происходит в 10 и более раз выше, чем у истинных. Это объясняется тем, что естественный отбор не может оказать на них воздействия.
Исследования японского генетика Кимури показывают, что в сохранении нейтральных мутаций в поколениях имеет важное значение дрейф генов.
В результате мутационной изменчивости происходит эволюция белка. Она может быть вредной или полезной. Полезные мутации благодаря естественному отбору сохраняются в генофонде популяции. Отдельные белки имеют очень устойчивую структуру и изменяются очень мало или почти не изменяются при переходе от организма одного вида в другой. Так, например, число различий по аминокислотному составу двух видов цепей гемоглобина между человеком и представителями других отрядов млекопитающих от 1 до 25, от 1 до 33.
Значение точковых мутаций в эволюции генов и белков. Каждая пара нуклеотидов в составе гена может подвергаться мутации. Мутация, затрагивающая пару нуклеотидов, называется точковой. Следует напомнить, что нуклеотиды, из которых состоит ДНК, по-разному реагируют на воздействие извне. В некоторых нуклеотидных парах под влиянием воздействий среды происходит мутация всего один или два раза. У других число таких мутаций может достигать нескольких сотен. Такие нуклеотидные пары, в которых часто повторяются мутации, называются «горячими точками».
Необходимо знать, что не всегда генная мутация приводит к изменению в строении и функции белка. Мутации, которые вызывают невидимые изменения, названы «молчащими мутациями». Различают два вида молчащих мутаций. При одних происходит простое замещение одного нуклеотида другим без изменения аминокислот в составе белка. Свидетельством правильности этого мнения является тот факт, что 18 из 20 входящих в состав белка аминокислот кодируется более чем одним кодоном (от двух до шести). При других мутациях происходит замена в белковой молекуле одной аминокислоты другой, однако такая замена не отражается на функциях белка.
В гене помимо кодовой части существуют участки (интроны), не принимающие участия в трансляциях. Мутации, возникающие на этих участках гена, если не принимать во внимание их влияние на структурно-регуляторные части, являются нейтральными.
Возникновение новых генов. Новые гены возникают не на пустом месте, а появляются в результате изменения существующих. В первой стадии появления нового гена происходит дупликация исходного и в большинстве случаев новый ген образуется вблизи той части хромосомы, где расположен исходный ген. К примеру, рассмотрим, какие изменения в историческом процессе произошли у гена, обуславливающего синтез белка гемоглобина, участвующего в переносе кислорода у животных. Исходный ген глобина первоначально подвергся дупликации 500 миллионов лет назад у первобытных представителей рыб. Затем в результате возникновения и накопления мутаций данного гена и естественного отбора произошла дивергенция и образовались молекулы глобина с с двумя видами цепей. В последующем гены каждой цепи, располагаясь в не гомологичных хромосомах, образовали два разных «семейства» генов. Одно семейство глобиновых генов образовалось благодаря дупликации исходного гена. У человека общая длина семейства глобиновых генов составляет 45 тысяч пар нуклеотидов и имеет пять активных генов. Из них 8% составляют экзоны, которые участвуют в синтезе глобиновой иРНК, еще 8% их относятся к интронам р-глобиновых генов. Функция остальных 84% ДНК, входящих в p-ген, еще не определена. Человек, горилла и павиан, происходившие от одного предка, имеют сходство в последовательности этих генов, что является результатом естественного отбора.
Кроме уникальных генов в геноме существуют и псевдогены, которые возникают в результате неудачной попытки создать новые копии генов. В семействе р-глобиновых генов человека их два. Такие «испорченные» псевдогены нормально не функционируют
Говоря о молекулярных основах эволюции, прежде всего, следует осветить изменения, которые произошли в строении и функциях нуклеиновых кислот, в частности ДНК, а также белковых молекул, в процессе исторического развития. На современном этапе молекулярной биологии можно анализировать число различий в последовательностях нуклеотидов в ДНК или аминокислот в молекуле белка разных видов и по этому показателю судить о степени их отличия. Поскольку каждая замена аминокислот в молекуле белка связана с изменением одного, двух или трех нуклеотидов в молекуле ДНК, с помощью компьютерных технологий можно вычислить максимальное или минимальное число нуклеотидных замен и на их основе судить о замещении аминокислот в молекуле белка.
Геном каждого растения, животного или гриба состоит из очень сложных и многообразных генетических элементов. В геноме встречаются, во-первых, так называемые уникальные гены, которые повторяются в геноме всего один или несколько раз, во-вторых, гены, повторяемые десятки и сотни раз, в-третьих, нуклеотиды, которые повторяются от десяти до тысячи раз, в – четвертых , образования, состоящие из коротких последовательностей нескольких нуклеотидов, которые могут повторяться миллионы раз (саттелитные последовательности ДНК).
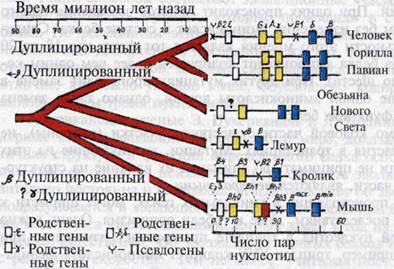
Рис. Эволюция набора глобиновых генов у млекопитающих. Слева — дивергенция
видов, а также многообразие и строение набора глобиновых генов (размеры
набора генов определяются тысячами пар нуклеотидов).
Уникальные гены, которые содержатся в геноме организмов различных типов и классов, сходны, например, блоки человека и бактерии. Многие виды, хотя и имеют идентичные системы генов или повторяющиеся нуклеотиды, отличаются друг от друга числом этих блоков.
В клетках эукариотических организмов ДНК содержится в тысячи раз больше, чем у прокариотов. Однако в пределах крупных таксономических категорий — типов и классов растений, животных, грибов нельзя судить о степени сложности их организации по содержанию ДНК в их клетках.
Митохондриальная ДНК (мтДНК) — ДНК, находящаяся (в отличие от ядерной ДНК) в митохондриях, органоидах эукариотических клеток.
Гены, кодированные мтДНК, относятся к группе плазмагенов, расположенных вне ядра (вне хромосомы). Совокупность этих факторов наследственности, сосредоточенных в цитоплазме клетки, составляет плазмон данного вида организмов (в отличие от генома). Митохондриальная ДНК была открыта Маргит Насс и Сильвен Насс в 1963 году в Стокгольмском университете при помощи электронной микроскопии и, независимо, учёными Эллен Харлсбруннер, Хансом Туппи и Готтфридом Шацем при биохимическом анализе фракций митохондрий дрожжей в Венском университете в 1964 году. Согласно эндосимбиотической теории, митохондриальная ДНК произошла от кольцевых молекул ДНК бактерий, и поэтому имеет иное происхождение, чем ядерный геном. Сейчас преобладает точка зрения, согласно которой митохондрии имеют монофилетическое происхождение, то есть были приобретены предками эукариот лишь однажды.
На основании сходства в последовательностях нуклеотидов ДНК ближайшими родственниками митохондрий среди ныне живущих прокариот считают альфа-протеобактерий (выдвигалась также гипотеза, что к митохондриям близки риккетсии). Сравнительный анализ геномов митохондрий показывает, что в ходе эволюции происходило постепенное перемещение генов предков современных митохондрий в ядро клетки. Необъяснимыми с эволюционной точки зрения остаются некоторые особенности митохондриальной ДНК (например, довольно большое число интронов, нетрадиционное использование триплетов и др.). Ввиду ограниченного размера митохондриального генома бо́льшая часть митохондриальных белков кодируется в ядре. При этом бо́льшая часть митохондриальных тРНК кодируются митохондриальным геномом. У большинства изученных организмов митохондрии содержат только кольцевые молекулы ДНК, у некоторых растений одновременно присутствуют и кольцевые, и линейные молекулы, а у ряда протистов (например, инфузорий) имеются только линейные молекулы. Митохондрии млекопитающих обычно содержат от двух до десяти идентичных копий кольцевых молекул ДНК. У растений каждая митохондрия содержит несколько молекул ДНК разного размера, которые способны к рекомбинации. У протистов из отряда кинетопластид (например, у трипаносом) в особом участке митохондрии (кинетопласте) содержится два типа молекул ДНК — идентичные макси-кольца (20-50 штук) длиной около 21 т.п.о. и мини-кольца (20 000 — 55 000 штук, около 300 разновидностей, средняя длина около 1000 п.о.). Все кольца соединены в единую сеть (катенаны), которая разрушается и восстанавливается при каждом цикле репликации. Макси-кольца гомологичны митохондриальной ДНК других организмов. Каждое мини-кольцо содержит четыре сходных консервативных участка и четыре уникальных гипервариабельных участка. В мини-кольцах закодированы короткие молекулы направляющих РНК (guideRNA), которые осуществляют редактирование РНК, транскрибируемых с генов макси-колец. Митохондриальная ДНК особенно чувствительна к активным формам кислорода, генерируемым дыхательной цепью, в связи с непосредственной их близостью. Хотя митохондриальная ДНК связана с белками, их защитная роль менее выражена, чем в случае ядерной ДНК. Мутации в ДНК митохондрий могут вызывать передаваемые по материнской линии наследственные заболевания. Также имеются данные, указывающие на возможный вклад мутаций митохондриальной ДНК в процесс старения и развитие возрастных патологий. У человека митохондриальная ДНК обычно присутствует в количестве 100—10000 копий на клетку (сперматозоиды и яйцеклетки являются исключением). С множественностью митохондриальных геномов связаны особенности проявления митохондриальных болезней — обычно позднее их начало и очень изменчивые симптомы.
У большинства многоклеточных организмов митохондриальная ДНК наследуется по материнской линии. Яйцеклетка содержит на несколько порядков больше копий митохондриальной ДНК, чем сперматозоид. В сперматозоиде обычно не больше десятка митохондрий (у человека — одна спирально закрученная митохондрия), в небольших яйцеклетках морского ежа — несколько сотен тысяч, а в крупных ооцитах лягушки — десятки миллионов. Кроме того, обычно происходит деградация митохондрий сперматозоида после оплодотворения.
При половом размножении митохондрии, как правило, наследуются исключительно по материнской линии, митохондрии сперматозоида обычно разрушаются после оплодотворения. Кроме того, большая часть митохондрий сперматозоида находятся в основании жгутика, которое при оплодотворении иногда теряется. В 1999 году было обнаружено, что митохондрии сперматозоидов помечены убиквитином (белком-меткой, которая приводит к разрушению отцовских митохондрий в зиготе).
Так как митохондриальная ДНК не является высококонсервативной и имеет высокую скорость мутирования, она является хорошим объектом для изучения филогении (эволюционного родства) живых организмов. Для этого определяют последовательности митохондриальной ДНК у разных видов и сравнивают их при помощи специальных компьютерных программ и получают эволюционное древо для изученных видов. Исследование митохондриальных ДНК собак позволило проследить происхождение собак от диких волков. Исследование митохондриальной ДНК в популяциях человека позволило вычислить «митохондриальную Еву», гипотетическую прародительницу всех живущих в настоящее время людей.
Для некоторых видов показана передача митохондриальной ДНК по мужской линии, например, у мидий. Наследование митохондрий по отцовской линии также описано для некоторых насекомых, например, для дрозофилы, медоносных пчел и цикад.
Существуют также данные о митохондриальном наследовании по мужской линии у млекопитающих. Описаны случаи такого наследования для мышей, при этом митохондрии, полученные от самца, впоследствии отторгаются. Такое явление показано для овец и клонированного крупного рогатого скота. Также описан единственный случай связанный с бесплодием у мужчины.
У млекопитающих каждая молекула мтДНК содержит 15000-17000 пар оснований (у человека 16565 пар нуклеотидов — исследование закончено в 1981 году, по другому источнику 16569 пар) и содержит 37 генов — 13 кодируют белки, 22 — гены тРНК, 2 — рРНК (по одному гену для 12S и 16S рРНК). Другие многоклеточные животные имеют схожий набор митохондриальных генов, хотя некоторые гены могут иногда отсутствовать. Генный состав мтДНК разных видов растений, грибов и особенно протистов различается более значительно.
Особенности митохондриальной ДНК.
Кодирующие последовательности (кодоны) митохондриального генома имеют некоторые отличия от кодирующих последовательностей универсальной ядерной ДНК.
Так, кодон AUA кодирует в митохондриальном геноме метионин (вместо изолейцина в ядерной ДНК), кодоны AGA и AGG — терминаторные кодоны (в ядерной ДНК кодируют аргинин), кодон UGA в митохондриальном геноме кодирует триптофан.Если говорить точнее, то речь идёт не о митохондриальной ДНК, а о мРНК, которая списывается (транскрибируется) с этой ДНК перед началом синтеза белка. Буква U в обозначении кодона обозначает уридин, который при транскрипции гена в РНК заменяет тимин. Количество генов тРНК (22 гена) меньше, чем в ядерном геноме с его 32 генами тРНК. В человеческом митохондриальном геноме информация настолько сконцентрирована, что в последовательностях кодирующих мРНК, как правило, частично удалены нуклеотиды, соответствующие 3'-концевым терминаторным кодонам.
Кроме изучения для построения различных филогенетических теорий, изучение митохондриального генома — основной инструмент при проведении идентификации. Возможность идентификации связана с существующими в митохондриальном геноме человека групповыми и даже индивидуальными различиями.
Вопросы для самоконтроля
1.В чём заключается проблема возникновения новых генов и их дальнейшей эволюции?
2. Назвать и охарактеризовать основные тенденции в эволюции генов.
3.Что гласит теория нейтральности (М. Кимура)?
4. Основные положения назвать «Недарвиновской эволюции».
5. Как оценивают скорости эволюции генов и белков?
6. На чём основана концепция молекулярных часов и её значение?
6. Как и почему использут митохондриальную ДНК в эволюционных исследованиях?
8. Как строят филогенетические древа на основе "молекулярных часов"?