

Гистология печени
Внутреннее строение печени взрослого человека подчинено архитектонике кровеносного и желчевыводящего русла.
Основной структурной единицей П. является печеночная долька. Клетки в ней образуют печеночные балки, расположенные по радиусам.
Между балками к центру дольки, где расположена центральная вена, тянутся синусоиды. На периферии дольки из желчных межклеточных капилляров формируются начальные желчные протоки (междольковые). Укрупняясь и сливаясь, они образуют в воротах П. печеночный проток, по которому желчь выходит из печени.
По Элиасу (Н. Elias, 1949), печеночная долька построена из системы печеночных пластинок, сходящихся по направлению к центру дольки и состоящих из одного ряда клеток. Между пластинками расположены лакуны, образующие лабиринт (рис. 5).
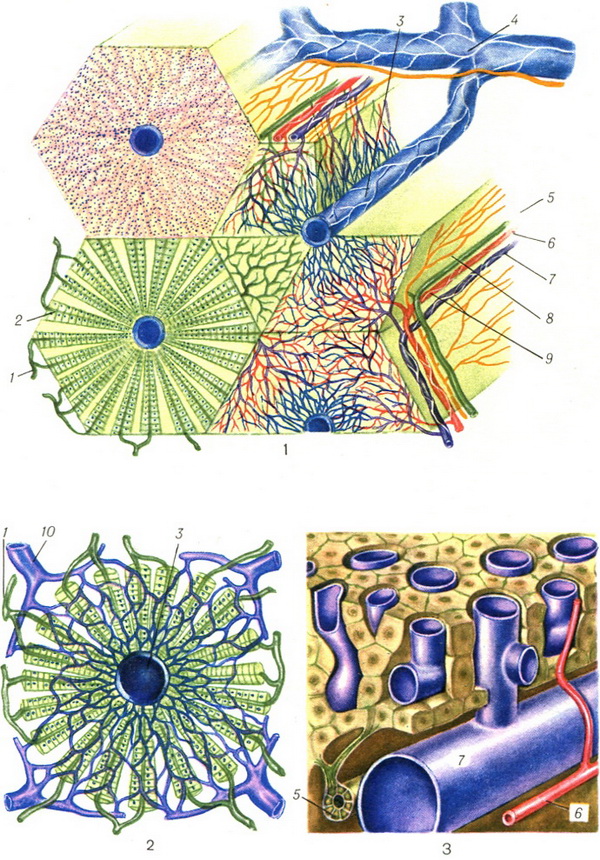 Рис.
1—3. Схемы строения печеночной дольки
(рис. 3 по Чайлду): 1—ductuli biliferi; 2 — желчные
капилляры; 3 — v. centralis; 4 — v. sublobularis; 5 —
ductus interlobularis; б —a. interlobularis; 7 —v.
interlobularis; 8 — междольковые лимфатические
капилляры; 9 — пернваскулярное нервное
сплетение; 10 — приток междольковых вен.
Рис.
1—3. Схемы строения печеночной дольки
(рис. 3 по Чайлду): 1—ductuli biliferi; 2 — желчные
капилляры; 3 — v. centralis; 4 — v. sublobularis; 5 —
ductus interlobularis; б —a. interlobularis; 7 —v.
interlobularis; 8 — междольковые лимфатические
капилляры; 9 — пернваскулярное нервное
сплетение; 10 — приток междольковых вен.
Из долек складываются области и сегменты печени, связанные с ветвями воротной вены и печеночных артерий. Различают передний и задний сегменты в веществе правой доли П., медиальный сегмент, занимающий территорию хвостатой и квадратной долей, и латеральный сегмент, соответствующий левой доле. Каждый из главных сегментов делится на два. П. построена из железистой эпителиальной ткани. Печеночные клетки разделяются желчными капиллярами (рис. 6).
|
|
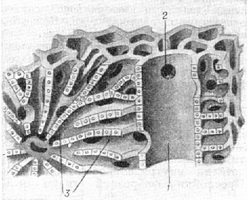 Рис.
5. Микроскопическое строение дольки
печени (по Элиасу); справа — портальное
пространство для приводящей вены (1),
ограниченное lamina limitans; видно отверстие
(2) для афферентной венулы, ведущее в
лабиринт; слева—лабиринт дольки (3),
лакуны которого ограничены печеночными
пластинками (laminae hepaticae); лакуны
конвергированы к центральному
пространству (для центральной вены).
Рис.
5. Микроскопическое строение дольки
печени (по Элиасу); справа — портальное
пространство для приводящей вены (1),
ограниченное lamina limitans; видно отверстие
(2) для афферентной венулы, ведущее в
лабиринт; слева—лабиринт дольки (3),
лакуны которого ограничены печеночными
пластинками (laminae hepaticae); лакуны
конвергированы к центральному
пространству (для центральной вены).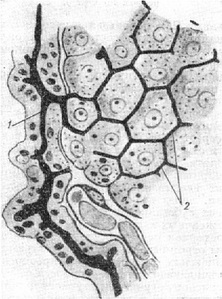 Рис.
6. Внутридольковый желчный прекапилляр
(1), дренирующий желчь из внутридольковых
желчных капилляров (2) (по Элиасу).
Рис.
6. Внутридольковый желчный прекапилляр
(1), дренирующий желчь из внутридольковых
желчных капилляров (2) (по Элиасу).
|
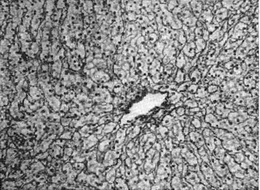 Рис.
7. Решетчатые (аргирофильные) волокна
внутри печеночной дольки (импрегнация
серебром по Футу).
Рис.
7. Решетчатые (аргирофильные) волокна
внутри печеночной дольки (импрегнация
серебром по Футу).Прослойки фиброзной ткани между дольками П. и паравазальные соединительнотканные тракты составляют строму печени. Здесь много коллагеновых волокон, в то время как в строме дольки представлены главным образом аргирофильные ретикулиновые волокна (рис. 7).
Цитохимия и ультраструктура печеночных клеток
Печеночная клетка — гепатоцит — имеет полигональную форму и размер от 12 до 40 мк в диаметре в зависимости от функционального состояния.
В гепатоците выделяют синусоидальный и билиарный полюсы. Через первый происходит всасывание различных веществ из крови, через второй — секреция желчи и других субстанций в просветы межклеточных желчных канальцев.
Абсорбирующая и секреторная поверхности гепатоцита снабжены огромным количеством ультрамикроскопических выростов — микроворсинок, увеличивающих эти поверхности.
Гепатоцит ограничен двухконтурной белково-липидной плазматической мембраной, обладающей высокой ферментативной активностью—фосфатазной на билиарном полюсе и нуклеозидфосфатазной — на синусоидальном. Плазматическая мембрана гепатоцита содержит и фермент транслоказу, катализирующую активный транспорт ионов и молекул в клетку и из нее. Цитоплазма гепатоцита представлена мелкозернистым матриксом с небольшой электронной плотностью и системой мембран, которые составляют одно целое с плазматической и ядерной оболочками. Последняя также двухконтурна, состоит из белков и липидов и окружает шаровидное ядро с 1 — 2 ядрышками. В ядерной оболочке имеются поры диаметром 300—500 А. Некоторые гепатоциты (с возрастом их становится больше) имеют по два ядра. Двуядерные клетки, как правило, полиплоидны. Митозы встречаются редко.
К органеллам гепатоцита относятся эндоплазматическая сеть (гранулярная и агранулярная), митохондрии и аппарат (комплекс) Гольджи.
Гранулярная эндоплазматическая сеть (эргастоплазма) построена из парных параллельных липопротеиновых мембран, ограничивающих ультрамикроскопические канальцы. На наружной поверхности этих мембран располагаются рибосомы — рибонуклеопротеиновые гранулы диаметром 100—150 А. Агранулярная эндоплазматическая сеть построена так же, но рибосом не имеет.
Митохондрии в числе 2000—2500 встречаются в виде нитей, палочек и зерен величиной 0,5—1,5 мк и расположены около ядра и по периферии клетки. Митохондрии гепатоцита содержат огромное количество ферментов и являются энергетическими центрами клетки.
Ультрамикроскопически — митохондрии сложные липопротеиновые мембранные структуры, осуществляющие ферментативные превращения трикарбоновых кислот, сопряжение потока электронов с синтезом АТФ, перенос активных ионов во внутренние пространства митохондрий, а также синтез фосфолипидов и жирных кислот с длинной цепью.
Аппарат Гольджи представлен сетью перекладин разной толщины, которые располагаются в разные фазы секреторного цикла гепатоцита около ядра или вблизи желчных канальцев. Ультрамикроскопически он состоит из агранулярных липопротеиновых мембран, образующих трубочки, пузырьки, мешочки и щели. Аппарат Гольджи богат нуклеозидфосфатазами и другими ферментами. Лизосомы — перибилиарные тельца — пузырьки диаметром 0,4 мк и меньше, ограниченные одноконтурными мембранами, расположены около просветов желчных канальцев. Они содержат гидролазы и особенно богаты кислой фосфатазой. Непостоянные включения (гликоген, жир, пигменты, витамины) по своему составу и количеству варьируют.
Эндогенные пигменты—это гемосидерин, липофусцин, билирубин. Экзогенные пигменты могут присутствовать в цитоплазме гепатоцитов в виде солей различных металлов.
В 1833 г. Кирнан ввёл понятие о дольках печени как основе её архитектоники. Он описал чётко очерченные дольки пирамидальной формы, состоящие из центрально расположенной печёночной вены и периферически расположенных портальных трактов, содержащих жёлчный проток, ветви воротной вены и печёночной артерии. Между этими двумя системами располагаются балки гепатоцитов и содержащие кровь синусоиды.
С помощью стереоскопической реконструкции и сканирующей электронной микроскопии показано, что печень человека состоит из столбиков гепатоцитов, отходящих от центральной вены, в правильном порядке чередующихся с синусоидами (рис. 1-9).
Ткань печени пронизана двумя системами каналов — портальными трактами и печёночными центральными каналами, которые расположены таким образом, что не касаются друг друга; расстояние между ними составляет 0,5 мм (рис. 1-10). Эти системы каналов расположены перпендикулярно друг другу. Синусоиды распределяются неравномерно, обычно проходя перпендикулярно линии, соединяющей центральные вены. Кровь из терминальных ветвей воротной вены попадает в синусоиды; при этом направление кровотока определяется более высоким давлением в воротной вене по сравнению с центральной.
Центральные печёночные каналы содержат истоки печёночной вены. Они окружены пограничной пластинкой печёночных клеток.
Портальные триады (синонимы: портальные тракты, глиссонова капсула) содержат терминальные ветви воротной вены, печёночную артериолу и жёлчный проток с небольшим количеством круглых клеток и соединительной ткани (рис. 1-11). Они окружены пограничной пластинкой печёночных клеток.
Анатомическое деление печени проводят по функциональному принципу. Согласно традиционным представлениям, структурная единица печени состоит из центральной печёночной вены и окружающих её гепатоцитов.
Однако Раппапорт [34] предлагает выделять ряд функциональных ацинусов, в центре каждого из которых лежит портальная триада с терминальными ветвями портальной вены, печёночной артерии и жёлчного протока — зона 1 (рис. 1-12 и 1-13). Ацинусы расположены веерообразно, в основном перпендикулярно по отношению к терминальным печёночным венам соседних ацинусов. Периферические, хуже кровоснабжаемые отделы ацинусов, прилежащие к терминальным печёночным венам (зона 3), наиболее страдают от повреждения (вирусного, токсического или аноксического). В этой зоне локализуются мостовидные некрозы. Области, расположенные ближе к оси, образованной приносящими сосудами и жёлчными протоками, более жизнеспособны, и позднее в них может начаться регенерация печёночных клеток. Вклад каждой из зон ацинуса в регенерацию гепатоцитов зависит от локализации повреждения [30, 34].
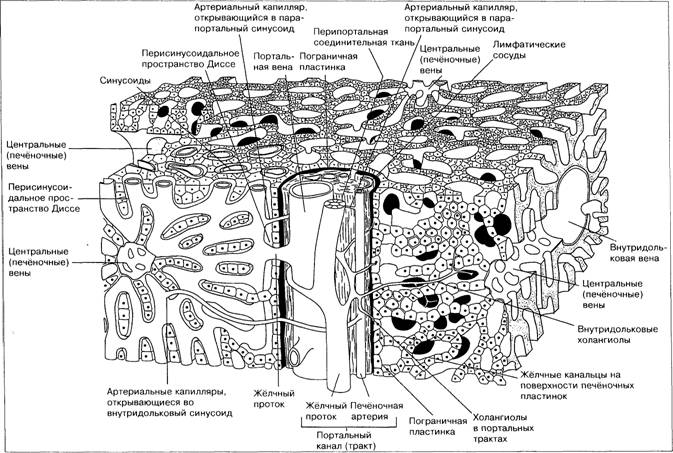
Рис. 1-9. Структура печени человека в норме.
Рис. 1-10. Гистологическое строение печени в норме. Н — терминальная печёночная вена; Р — портальный тракт. Окраска гематоксилином и эозином, х60. См. также цветную иллюстрацию на с. 767.
Рис. 1-11. Портальный тракт в норме. А — печёночная артерия; Ж — жёлчный проток. В — портальная вена. Окраска гематоксилином и эозином. См. также цветную иллюстрацию на с. 767.
Печёночные клетки (гепатоциты) составляют около 60% массы печени. Они имеют полигональную форму и диаметр, равный приблизительно 30 мкм. Это одноядерные, реже многоядерные клетки, которые делятся путём митоза. Продолжительность жизни гепатоцитов у экспериментальных животных составляет около 150 дней. Гепатоцит граничит с синусоидом и пространством Диссе, с жёлчным канальцем и соседними гепатоцитами. Базальной мембраны у гепатоцитов нет.
Синусоиды выстланы эндотелиальными клетками.
К синусоидам относятся фагоцитирующие клетки ретикулоэндотелиальной системы (клетки Купфера),
звёздчатые клетки, также называемые жирозапасающими, клетками Ито или липоцитами.
В каждом миллиграмме нормальной печени человека содержится приблизительно 202•103 клеток, из которых 171•103 являются паренхиматозными и 31•103 — литоральными (синусоидальные, в том числе клетки Купфера).
Пространством Диссе называется тканевое пространство между гепатоцитами и синусоидальными эндотелиальными клетками. В перисинусоидальной соединительной ткани проходят лимфатические сосуды, которые на всём протяжении выстланы эндотелием. Тканевая жидкость просачивается через эндотелий в лимфатические сосуды.
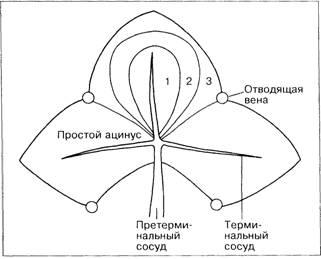
Рис. 1-12. Функциональный ацинус (по Раппапорту). Зона 1 примыкает к входной (портальной) системе. Зона 3 примыкает к выводящей (печёночной) системе.
Ветви печёночной артериолы образуют сплетение вокруг жёлчных протоков и впадают в синусоидальную сеть на различных её уровнях. Они снабжают кровью структуры, расположенные в портальных трактах. Прямых анастомозов между печёночной артерией и воротной веной нет.
Экскреторная система печени начинается с жёлчных канальцев. Они не имеют стенок, а являются просто углублениями на контактирующих поверхностях гепатоцитов , которые покрыты микроворсинками. Плазматическая мембрана пронизана микрофиламентами, образующими поддерживающий цитоскелет. Поверхность канальцев отделена от остальной межклеточной поверхности соединительными комплексами, состоящими из плотных контактов, щелевых контактов и десмосом. Внутридольковая сеть канальцев дренируется в тонкостенные терминальные жёлчные протоки или дуктулы (холангиолы, канальцы Геринга), выстланные кубическим эпителием. Они заканчиваются в более крупных (междольковых) жёлчных протоках, расположенных в портальных трактах. Последние разделяются на мелкие (диаметром менее 100 мкм), средние (±100 мкм) и крупные (более 100 мкм).

Рис. 1-13. Кровоснабжение простого ацинуса печени, зональное расположение клеток и микроциркуляторное периферическое русло. Ацинус занимает примыкающие секторы соседних шестиугольных полей. Зоны 1, 2 и 3 соответственно представляют области, снабжаемые кровью с I, II и III степенью содержания кислорода и питательных веществ. В центре этих зон находятся терминальные ветви приносящих сосудов, жёлчных протоков, лимфатических сосудов и нервов (PS), а сами зоны простираются до треугольных портальных полей, из которых выходят эти ветви. Зона 3 оказывается на периферии микроциркуляторного русла ацинуса, поскольку её клетки так же удалены от афферентных сосудов своего ацинуса, как и от сосудов соседнего ацинуса.
Перивенулярная область образуется наиболее удалёнными от портальной триады частями зоны 3 нескольких прилежащих ацину-сов. При повреждении этих зон повреждённая область приобретает вид морской звезды (затемнённая область вокруг терминальной печёночной венулы, расположенной в её центре — ЦПВ). 1, 2, 3 — зоны микроциркуляции; Г, 2', 3' — зоны соседнего ацинуса .
Поверхность гепатоцитов ровная, за исключением нескольких участков прикрепления (десмосом). Из них в просвет жёлчных канальцев выдаются равномерно расположенные микроворсинки одинаковых размеров. На поверхности, обращённой к синусоиду, располагаются микроворсинки разной длины и диаметра, проникающие в перисинусоидальное тканевое пространство. Наличие микроворсинок свидетельствует об активной секреции или абсорбции (в основном жидкости).
Ядро содержит дезоксирибонуклеопротеин. Печень человека после полового созревания содержит тетраплоидные ядра, а в возрасте 20 лет — также октоплоидные ядра. Считается, что повышенная полиплоидность свидетельствует о предраковом состоянии.
В хроматиновой сети обнаруживаются одно или два ядрышка. Ядро имеет двойной контур и содержит поры, обеспечивающие обмен с окружающей цитоплазмой.
Митохондрии также имеют двойную мембрану, внутренний слой которой образует складки, или кристы. Внутри митохондрий протекает огромное количество процессов, в частности окислительное фосфорилирование, при которых происходит освобождение энергии. В митохондриях содержится много ферментов, в том числе участвующих в цикле лимонной кислоты и бета-окислении жирных кислот. Энергия, высвобождающаяся в этих циклах, затем запасается в виде АДФ. Здесь протекает также синтез гема.
Шероховатая эндоплазматическая сеть (ШЭС) выглядит как ряд пластинок, на которых располагаются рибосомы. При световой микроскопии они окрашиваются базофильно. В них синтезируются специфические белки, особенно альбумин, белки свёртывающей системы крови и ферменты. При этом рибосомы могут сворачиваться в спираль, образуя полисомы. В ШЭС синтезируется Г-6-Фаза. Из свободных жирных кислот синтезируются триглицериды, которые в виде липопротеидных комплексов секретируются путём экзоцитоза. ШЭС может участвовать в глюкогенезе.
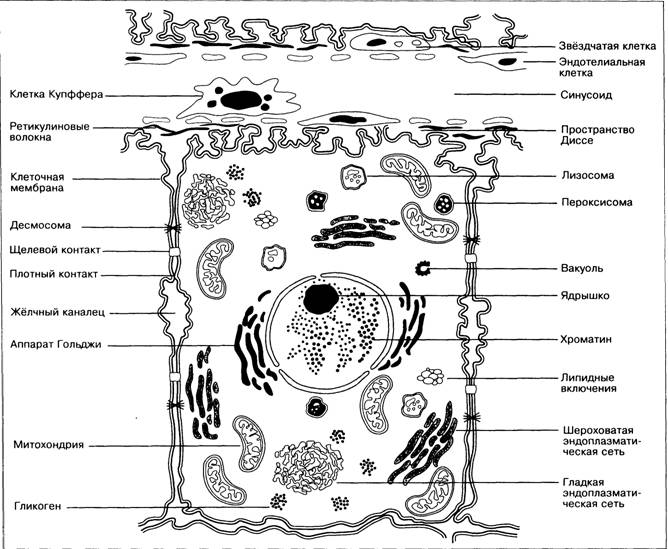
Рис. 1-14. Органеллы гепатоцита.
Гладкая эндоплазматическая сеть (ГЭС) образует тубулы и везикулы. Она содержит микросомы и является местом конъюгации билирубина, детоксикации многих лекарств и других токсичных веществ (система Р450). Здесь синтезируются стероиды, в том числе холестерин и первичные жёлчные кислоты, которые конъюгируют с аминокислотами глицином и таурином. Индукторы ферментов, например фенобарбитал, увеличивают размеры ГЭС.
Пероксисомы располагаются поблизости от ГЭС и гранул гликогена. Их функция неизвестна.
Лизосомы — плотные тельца, примыкающие к жёлчным канальцам. Они содержат гидролитические ферменты, при выделении которых клетка разрушается. Вероятно, они выполняют функцию внутриклеточной очистки от разрушенных органелл, срок жизни которых уже истёк. В них откладываются ферритин, липофусцин, жёлчный пигмент и медь. Внутри них можно наблюдать пиноцитозные вакуоли. Некоторые плотные тельца, расположенные около канальцев, называются микротельцами.
Аппарат Гольджи состоит из системы цистерн и пузырьков, которые также лежат около канальцев. Его можно назвать «складом веществ», предназначенных для экскреции в жёлчь. В целом эта группа органелл — лизосомы, микротельца и аппарат Гольджи — обеспечивает секвестрирование любых веществ, которые были поглощены и должны быть удалены, секретрированы или сохранены для метаболических процессов в цитоплазме. Аппарат Гольджи, лизосомы и канальцы подвергаются особенно выраженным изменениям при холестазе.
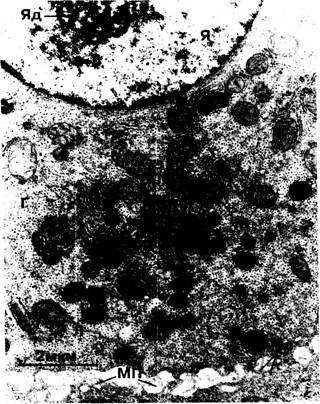
Рис. 1-15. Электронно-микроскопическая картина части нормального гепатоцита. Я — ядро; Яд — ядрышко; М — митохондрии; Ш — шероховатая эндоплазматическая сеть; Г — гранулы гликогена; mb— микроворсинки во внутриклеточном пространстве; Л — лизосомы; МП — межклеточное пространство.
Цитоплазма содержит гранулы гликогена, липиды и тонкие волокна.
Цитоскелет, поддерживающий форму гепатоцита, состоит из микротрубочек, микрофиламентов и промежуточных филаментов [15]. Микротрубочки содержат тубулин и обеспечивают перемещение органелл и везикул, а также секрецию белков плазмы. Микрофиламенты состоят из актина, способны к сокращению и играют важную роль в обеспечении целостности и моторики канальцев, тока жёлчи. Длинные ветвящиеся филаменты, состоящие из цитокератинов, называют промежуточными филаментами [42]. Они соединяют плазматическую мембрану с перинуклеарной областью и обеспечивают стабильность и пространственную организацию гепатоцитов.