

Ротация печени плода. Развитие поджелудочной железы эмбриона
Уже на ранней стадии развития от каудальной части печеночного узла отделяется закладка желчного пузыря, которая вначале выглядит как плотное цилиндрическое образование, а впоследствии в нем образуется просвет. Желчь в нем начинает накапливаться приблизительно с конца третьего месяца.
При ротации кишечной петли из области вентральной брыжейки, лежащей между двенадцатиперстной кишкой и закладкой печени, образуется печеночно-желудочная и печеночно-двенадцатиперстная части малого сальника (pars hepatogastrica и pars hepatoduodenalis omenti minoris). Из вентрального отдела вентральной брыжейки, расположенного между печенью и передней стенкой тела, возникает серповидная связка печени (mesohepaticum ventrale, ligamentum falciforme hepatis).
Большая часть первоначального стебелька, связывающего печеночный узел со стенкой двенадцатиперстной кишки, удлиняется и замывается в закладке желчного протока (ductus choledochus) и в закладке печеночного протока (ductus hepaticus). Закладка желчного пузыря связывается с данной трубочкой посредством короткой ножки, которая, удлиняясь, превращается в пузырный проток. Печеночный проток возникает из той части закладки для желчного протока, которая располагается между устьем пузырного протока и печенью.
Первоначально все эти выводные протоки в основной своей массе закладываются в виде плотных клеточных тяжей, в которых со временем (приблизительно во втором месяце) образуются просветы.
Междольковые желчные протоки (ductus interlobulares) закладываются в качестве первичных щелей (каналов) в эпителиальных балочках печеночной закладки (приблизительно в конце первого месяца), а позже вторично соединяются с системой крупных желчных протоков.
Интерстициальная соединительная ткань печени образуется из мезенхимы, в которую врастает закладка печени в процессе своего развития. Из веточек пупочно-брыжеечных вен между печеночными балками образуется система синусоидных капилляров. В течение первого месяца в мезенхиме происходит образование кровяных островков и первичных клеток крови (эмбриональный миэлопоэз), которое, однако, прекращается в более поздние месяцы. Из поверхности вентральной брыжейки двенадцатиперстной кишки образуется перитонеальное (брюшинное) покрытие печени, причем печень постепенно обособляется от поперечной перегородки, которая дает начало вентральным участкам диафрагмы.
Развитие поджелудочной железы эмбриона
Из энтодермальной стенки двенадцатиперстной кишки в конце первого месяца начинают развиваться два следующих эпителиальных узла, которые являются закладками поджелудочной железы. Один из них выпячивается на дорсальной стенке двенадцатиперстной кишки непосредственно под пилорической (привратниковой) частью и представляет собой дорсальную закладку поджелудочной железы (pancreas dorsale). Вторая закладка возникает на вентральной двенадцатиперстной поверхности в хвостовом узле между стенкой кишки и зачатком печени и, по всей вероятности, закладывается в виде парного бугорка (pancreas ventrale).
Дорсальный зачаток поджелудочной железы растет сравнительно быстрее и в виде плотных эпителиальных балок и узлов проникает в мезенхиму дорсальной брыжейки желудка и брыжейки двенадцатиперстной кишки. Приблизительно на шестой неделе закладка уже представляет собой вытянутое плотное образование, посередине которого проходит центральный проток и который своей эпителиальной ножкой соединяется с дорсальной стенкой двенадцатиперстной кишки. Вентральная поджелудочная железа сравнительно меньше и посредством своей соединяющей ножки (выводного протока) связана с зачатком желчного протока, который выходит из печени и удлиняется.
В результате неравномерного роста стенки двенадцатиперстной кишки и под влиянием быстрого роста печеночной закладки вентральная поджелудочная железа вместе с желчным протоком начинает смещаться по правой поверхности двенадцатиперстной кишки на дорсальную сторону кишечника, располагаясь затем под проксимальным отделом первоначальной дорсальной закладки поджелудочной железы. На седьмой неделе развития обе закладки поджелудочной железы прочно срастаются между собой. При этом из дорсальной поджелудочной железы образуются ее тело и хвост (corpus и cauda pancreatis), а из вентрального зачатка возникает самая большая ее часть — головка поджелудочной железы (caput pancreatis). Соединение этих частей происходит настолько прочно, что на поджелудочной железе взрослого человека уже невозможно распознать обе первоначальные закладки.
Секреция
Секреция выработка и выделение железистыми клетками секретов. По существу, в каждой клетке организма в ходе её жизнедеятельности образуются некоторые продукты метаболизма, выделяемые либо во внешнюю среду, либо во внутреннюю. Если секреторная функция становится основной для выполняющих её специализированных, т. н. железистых, клеток, то её называют С. Различают С. внешнюю, или экзокринную, когда продукты, вырабатываемые железой (См. Железы), выделяются из организма во внешнюю среду (секрет сначала поступает в проток железы, а затем выводится на поверхность тела или в полые органы), и внутреннюю (эндокринную) С., или инкрецию, — выделеление синтезируемых веществ в кровь или лимфу.
В секреторном цикле всякой железы различают фазу продукции (биосинтеза) секрета и фазу его выделения. Иногда термином «С.» обозначают только эту последнюю фазу цикла. В некоторых железах обе фазы протекают одновременно, в других же — разделены во времени, т. к. регулируются разными механизмами, особыми для каждой из них. Процесс С. — внутриклеточный конвейер, по которому синтезируемый продукт, постепенно созревая, последовательно передаётся в клетке от одного органоида к другому. Исходные продукты (аминокислоты, моносахариды, жирные кислоты, соли) поглощаются железистой клеткой из крови и тканевой жидкости (рис. 1, I). Биосинтез секрета (особенно белковых продуктов) начинается в эндоплазматической сети, где аминокислоты, адсорбирующиеся на клеточных мембранах, соединяются между собой в последовательности, определяемой РНК рибосом. Синтезируемый начальный продукт накапливается в щелях и лакунах эндоплазматической сети (рис. 1, II), откуда перемещается в область пластинчатого комплекса, или Гольджи комплекса (рис. 1, III, IV), где заканчивается созревание секрета. В некоторых железистых клетках в зоне комплекса Гольджи происходит соединение синтезируемого белка с углеводами и секрет превращается в гликопротеид. Митохондрии, многочисленные в железистых клетках, дают энергию, необходимую как для синтеза секрета, так и для его выделения. Кроме того, на митохондриях совершается синтез секретов липидной (стероидной) природы. В фазе выделения секрета значительно возрастает потребление кислорода железистой клеткой, повышается внутриклеточное осмотическое давление и увеличивается поступление воды в клетку. В результате в железистой клетке устанавливается ток воды, поступающей через её основание и выходящей через апикальную мембрану. Протекая через цитоплазму, вода выносит накопленный секрет либо в виде раствора, диффундирующего через апикальную мембрану (рис. 2, 2), либо в виде капель, выходящих через её поры (рис. 2, 3). При таком способе выделения секрета, называемом мерокриновой С., железистые клетки не претерпевают никаких разрушений. Если же секрет нерастворим в воде или по др. причинам не может пройти через апикальную мембрану, усиленное вхождение воды в набухающую железистую клетку приводит к тому, что верхушка клетки вместе со скапливающимися в ней гранулами или каплями секрета булавовидно вздувается, а затем либо лопается, либо отшнуровывается. Освобождение секрета путём отрыва верхушки железистой клетки без её гибели называется апокриновой С. (рис. 2, 5, 6). Иногда такая С. ограничивается набуханием и отрывом микроворсинок от железистой клетки (микроапокриновый тип С.) (рис. 2, 4). Наконец, иногда железистая клетка, дегенерируя, полностью превращается в каплю секрета и выталкивается из эпителиального пласта в просвет железы (рис. 2, 7) — голокриновая С. Этот примитивный тип С. в ходе эволюции заменяется более эффективным мерокриновым способом С.
Регуляция обеих фаз секреторного цикла определяется совместным или последовательным влиянием нескольких регулирующих факторов (нервных и гуморальных). Нервные волокна, несущие к железам импульсы, активирующие выделение секрета, называются секреторными. Нервные же эффекты, проявляющиеся увеличением выработки секрета, т. е. влияющие на фазу продукции, именуются трофическими. Однако отчётливого разграничения между секреторными и трофическими нервами нет, т. к. нередко раздражение волокна, иннервирующего железу, вызывает эффекты обоих родов. На деятельность желёз влияют также гуморальные агенты, в том числе некоторые гормоны (особенно в регуляции функциональной активности эндокринных желёз). Так, тиреотропный, гонадотропные и адренокортикотропный гормоны передней доли гипофиза возбуждают деятельность соответственно щитовидной железы, яичников и семенников, коры надпочечников (глюкокортикоидную функцию последней).Секретин, продуцируемый в слизистой оболочке 12-перстной кишки, стимулирует отделение панкреатического сока ацинарными клетками поджелудочной железы.
Наряду с гормонами на функцию желёз могут влиять и другие вещества, образующиеся в организме, такие, как Гистамин, резко усиливающий секрецию фундальных желёз желудка. Влияние гуморальных стимуляторов проявляется на обеих фазах секреторного цикла. Наконец, на С. многих желёз непосредственно влияют некоторые ионы, причём избыток одновалентных катионов (К+ или Na+), как правило, усиливает выделение секрета, тогда как двухвалентные ионы (Ca2+, Mg2+), наоборот, ослабляют С. В основе возбуждения железистой клетки лежит активация аденилциклазы — фермента, локализующегося в поверхностной мембране этой клетки. Аденилциклаза вызывает образование циклического аденозинмонофосфата, который регулирует цепь внутриклеточных реакций, приводящих к возрастанию активности специфических ферментных систем, определяющих процесс С. Поэтому множественность факторов, влияющих на С., объясняется тем, что они одинаково способны активировать аденилциклазный механизм железистой клетки. Способность к секреторной деятельности свойственна также нервным клеткам. Все они вырабатывают и выделяют Медиаторы; в некоторых нервных клетках, называемых нейросекреторными, продукция физиологически активных веществ (нейрогормонов (См. Нейрогормоны)) достигает высокой интенсивности (см.Нейросекреция).
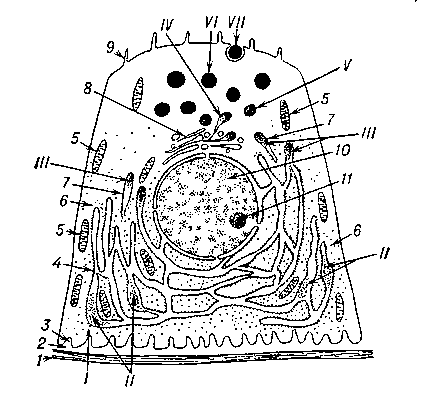
Рис. 1. Железистая клетка в фазе продукции. Схема процессов синтеза белкового секрета: I — поступление исходных веществ в железистую клетку; II — начало синтеза белка на пластинках гранулярной эндоплазматической сети; III — перемещение и накопление предстадий секрета в переходных ответвлениях эндоплазматической сети; IV — конденсация и дозревание секрета в зоне комплекса Гольджи; V — дозревающие и обособляющиеся гранулы секрета; VI — зрелые гранулы секрета; VII — выведение гранулы секрета из железистой клетки; 1 — базальная мембрана; 2 — цементирующее вещество; 3 — выпячивания базальной цитолеммы; 4 — гранулярная эндоплазматическая сеть; 5 — митохондрии; 6 — свободные рибосомы; 7 — переходное ответвление эндоплазматической сети; 8 — комплекс Гольджи; 9 — микроворсинки; 10 — ядро и двойная ядерная мембрана; 11 — ядрышко.
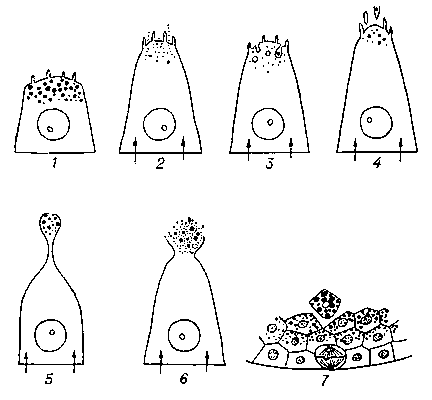
Рис. 2. Фаза секреции. Схема способов выделения секрета, или типы секреции: 1 — железистая клетка в состоянии функционального покоя; 2 — состояние возбуждения клетки (стрелки — усиленное вхождение воды в железистую клетку, что приводит к ее набуханию и растворению секрета); 3 — мерокриновая секреция; 4 — микроапокриновая секреция; 5 и 6 — апокриновая секреция; 7 — голокриновая секреция.