

Функционирование специализированных секреторных структур у растений
9.2.1
зделение нектара

бранных белков с участием кальмодулина (см. 6.7). Молекулярный механизм везикулярной секреции в растительных клетках не изучен. Однако известно, что и здесь необходим Са2 + . По-видимому, процессы секреции у растений аналогичны тому, что известно для клеток животных.
К наружным секреторным структурам относятся железистые волоски (трихомы), желёзки (солевые, насекомоядных растений), нектарники, осмофоры1 и гидатоды. Внутренние секреторные структуры представлены идиобластами2, вместилищами и млечниками.
Выделение Сахаров у растений осуществляется специализированными секреторными образованиями — нектарниками. Нектарники могут быть локализованы на различных частях цветка (цветковые, или флоральные, нектарники) либо на вегетативных частях растения — стеблях, листьях, прилистниках (внецветковые, или экстрафлоральные, нектарники).
Разнообразно анатомическое строение нектарников. Самые простые не содержат секреторных клеток и состоят из межклеточных полостей, примыкающих с одной стороны к элементам флоэмы, а с другой — к устьицам. Нектар, представляющий собой в этом случае флоэмный сок, выделяется через устьица. Если нектарник состоит из одного слоя эпидермальных клеток, покрытых кутикулой, то нектар выделяется через поры в кутикуле или через ее разрывы. Чаще всего нектарник состоит из эпидермальных клеток с железистыми волосками и устьицами и многоклеточной железистой паренхимной ткани. К секреторной ткани нектарника примыкают либо только флоэмные элементы, либо флоэмные и ксилемные. Нектар выделяется эпи-дермальными клетками или секреторными волосками. В период активного функционирования секреторные клетки содержат много митохондрий, в них хорошо развиты ЭР и пластиды. Вакуоли в секреторных клетках небольшие, АГ менее развит.
Нектар — многокомпонентный продукт. Основу его составляет несколько измененный флоэмный сок. В нектаре преобладают глюкоза, фруктоза и сахароза (во флоэмном соке — сахароза). В небольших концентрациях в нем представлены минеральные ионы (К+, Na+, Са2 + , Mg2+, РО|"), ди- и трикарбо-новые органические кислоты, витамины (аскорбиновая, никотиновая, фолиевая кислоты и др.), белки, иногда липиды. Концентрация Сахаров может меняться от 7 до 87% в зависимости от типа нектарников, вида растений, условий влажности и температуры. Флоральные нектарники могут также образовывать вещества типа стероидных гормонов, необходимые для репродуктивных процессов у насекомых.
1 Осмофоры
—
желёзки,
расположенные в различных частях
цветка, вырабатывающие эфирные
масла, от которых зависит аромат
цветков.
2 Секреторные
идиобласты
—
одиночные
клетки, служащие для отложения
каких-либо веществ. Обычно отличаются
по размерам или djODMe
от
ок ужаюших клеток.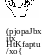
 Рис.
9.1
Рис.
9.1
Схема возможных механизмов транспорта и секреции нектара (U. Luttge, N. Higin-batham, 1979):
СТ — ситовидная трубка, КС —клетка-спутник, КП — клетка паренхимы, СК — секреторная клетка, С — сахароза, Г — глюкоза, Ф — фруктоза. Заштрихованы инкрустации клеточных стенок. Маленькими стрелками обозначен транспорт через плазмодесмы и симпласт. толстыми — метаболически контролируемый транспорт; 1 — активная разгрузка ситовидных трубок, 2 — насос, концентрирующий сахара в секреторных клетках, 3 — активная секреция, 4 — пассивная утечка нектара (тонкая красная стрелка), осуществляющаяся одновременно с активным процессом реабсорбции нектара (толстая синяя стрелка)
На секрецию нектара влияет продуктивность фотосинтеза. Для образования нектара важен транспорт ассимилятов по флоэме из ближайших к нектарнику листьев. В тканях нектарника сахара флоэмного сока метаболизируются (сахароза расщепляется на глюкозу и фруктозу и т. д.), к ним присоединяются сахара, образующиеся из крахмала нектарников. Секреция нектара сопровождается усилением поглощения кислорода тканями нектарников, зависит от температуры и подавляется недостатком 02, ингибиторами дыхательной цепи и окислительного фосфорилирования. При неблагоприятных условиях ткани нектарника могут поглощать (ресорбировать) компоненты нектара. У нектарников различных растений наблюдается суточный ритм деятельности, зависящий от внешних условий.
Этапы образования и выделения нектара изучены недостаточно. Полагают (рис. 9.1), что первым звеном его активного транспорта служит выход флоэмного сока из ситовидных трубок. Дальнейший транспорт преднектара возможен как по клеточным стенкам (по апопласту), так и по симпласту, через плазмодесмы. Для этапа секреции нектара предложен механизм эккриновой секреции, возможно, с помощью Н +-сахар — котранспортного механизма (см. 1.1.1 и 8.2.3). Предполагается также участие гранулокриновой секреции с участием элементов ЭР.
Флоральные нектарники возникли у покрытосеменных растений в меловом периоде мезозойской эры. Наряду с окрашенным венчиком и ароматом цветков нектар явился важным элементом энтомофилии — приспособлением для привлечения насекомых, осуществляющих перекрестное опыление. Появление нектара стало мощным фактором сопряженной эволюции насекомых и растений: множественных изменений цветка и приспособительного изменения режима питания и ротового аппарата насекомых-опылителей. Нектаром экстрафлоральных нектарников питаются муравьи, осы и другие насекомые.
Нектар, обладающий фитонцидными и бактериостатиче-скими свойствами, защищает завязь от микроорганизмов.
9.2.2
Секреция полисахаридов

1(^1К А.
9.2.3
Секреция белков
У растений существуют специализированные структуры, выделяющие полисахаридную слизь. Слизь образуют секреторные идиобласты, в которых она накапливается между клеточной стенкой и плазмалеммой. Слизь может вырабатываться эпителием слизевых ходов, расположенных в различных органах растений, а также выделяется специализированными волосками и желёзками на листьях и стеблях. У этих структур секрет накапливается под кутикулой, разрывы которой затем восстанавливаются.
Выделяемая полисахаридная слизь состоит из кислых и нейтральных полисахаридов, полисахаридов, близких к пектиновым веществам, или из кислых мукополисахаридов (у элодеи). У водорослей слизи сульфатированы. Все слизи растений сильно оводнены. Главная особенность ультраструктуры сли-зеобразующих клеток — преобладание АГ, который находится в состоянии гиперсекреции: содержит многочисленные диктио-сомы с множеством пузырьков. Секреция слизи осуществляется по гранулокриновому типу. Включение везикул аппарата Гольджи со слизью в экстраплазматическое пространство происходит путем экзоцитоза.
В клетках корневого чехлика слизь откладывается между плазмалеммой и клеточной стенкой, затем проникает через стенку и достигает поверхности. Каждая везикула Гольджи освобождается от полисахаридов в среднем за 30 с. Секреторная активность осуществляется ритмично с максимумами через каждые 3 ч, причем процесс секреции синхронен у всех клеток чехлика. По мере секреции клетки чехлика претерпевают прогрессирующую вакуолизацию и в конце концов сами становятся слизью (голокриновая секреция).
Функции, выполняемые секретируемой слизью, различны. У насекомоядных растений это ловчая слизь. В архегониях мхов она является средой, по которой движутся сперматозоиды. Слизь корневого чехлика выполняет защитную функцию (от повреждений кончика корня частицами почвы) и облегчает перемещение корня в почве. Слизи способствуют удержанию воды на поверхности или в полостях растения, участвуют в регуляции прорастания семян, у которых эпидер-мальные клетки выделяют слизь, набухающую при намачивании. В слизи могут содержаться молекулы специфических белков — лектинов, которые защищают орган (или клетку) от проникновения инфекции или способствуют формированию благоприятной для данного растения микрофлоры.
Из протеиногенных секреторных структур растений наиболее известны пищеварительные желёзки насекомоядных растений. Однако секретировать белок могут и желёзки листьев обычных растений. Например, у желёзок тополя и ивы в течение онтогенеза листа состав секрета меняется от терпеноидно-го до гликопротеинового. Еще одним примером протеиногенных клеток служат клетки алейронового слоя злаков.
Особенностью протеиногенных клеток является преобладание в них гранулярного ЭР, который синтезирует секрети-пуемые белки, и активное состояние АГ, в котором осущест-
вляются гликозилирование белков и образование гликопротеи-нов; в клетках много митохондрий.
9.2.4
Протеины могут накапливаться в специализированных алейроновых вакуолях (запасные белки в алейроновых клетках бобовых и злаков), выделяться во внутреннюю среду организма (например, гидролитические ферменты алейроновых клеток в эндосперме злаков) или выводиться наружу (гликопротеины листьев тополя, гидролазы пищеварительного сока насекомоядных растений и др.). Секреция протеаз у насекомоядных растений индуцируется хеморецепторами, чувствительными к азоту и фосфору. Выделение гидролитических ферментов из алейроновых клеток злаков находится под контролем гиббереллина.
11Ш*шшшт у растений имеются три группы секреторных эпидер-Выделение солей мальных образований, участвующих в выделении минеральных веществ:
Солевые железы листьев и стеблей галофитов. Они выделяют избыток ионов, поглощаемых растением при его росте в высокосолевой среде. Одновременно с солями железы теряют много воды. Соли поступают наружу, где смываются дождем или откладываются на кутикуле (рис. 9.2).
Солевые волоски на листьях, состоящие из двух клеток: ножки и головки. Когда в вакуоли головки накапливается много солей, головка отрывается (апокриновая секреция). На ее месте несколько раз в течение роста листа образуется новая головка. Солевые волоски теряют очень мало воды и широко распространены у растений в условиях засоления (рис. 9.2).
Секреторные клетки насекомоядных растений, выделяющие ионы, воду и гидролитические ферменты.
Важно отметить, что секреторные системы, выделяющие соли, не имеют непосредственной связи с проводящей системой растения. Ионы в железах и волосках транспортируются по симпласту, так как боковые стенки их клеток пропитаны суберином и лигнином. Секреция солей осуществляется с помощью мембранных транспортных помп или в везикулах. У солевых желёзок лимониума в плазмалемме наружных клеток функционирует электрогенный хлорный насос, направленный наружу. Сопряженно с хлором пассивно транспортируется натрий. Активность CP-насоса индуцируется повышением концентрации солей в наружном растворе и является необходимой частью механизма адаптации растения к изменению солености среды. Точно так же накопление ионов в головке секреторного волоска наблюдается только при увеличении концентрации NaCl во внешней среде. Транспорт ионов в желёзках и волосках регули-руется светом и осуществляется с затратой энергии АТР.
Секреция этих вторичных метаболитов осуществляется Секреция у растений одноклеточными образованиями (масляные клетки,
терпеноидов секреторные идиобласты, нечленистые млечники) или много-
клеточными структурами (железистые волоски, желёзки, железистый эпидермис, железистый эпителий смоляных ходов и вместилищ, членистые млечники, осмофоры). У многоклеточных секреторных структур (кроме членистых млечников) ос-
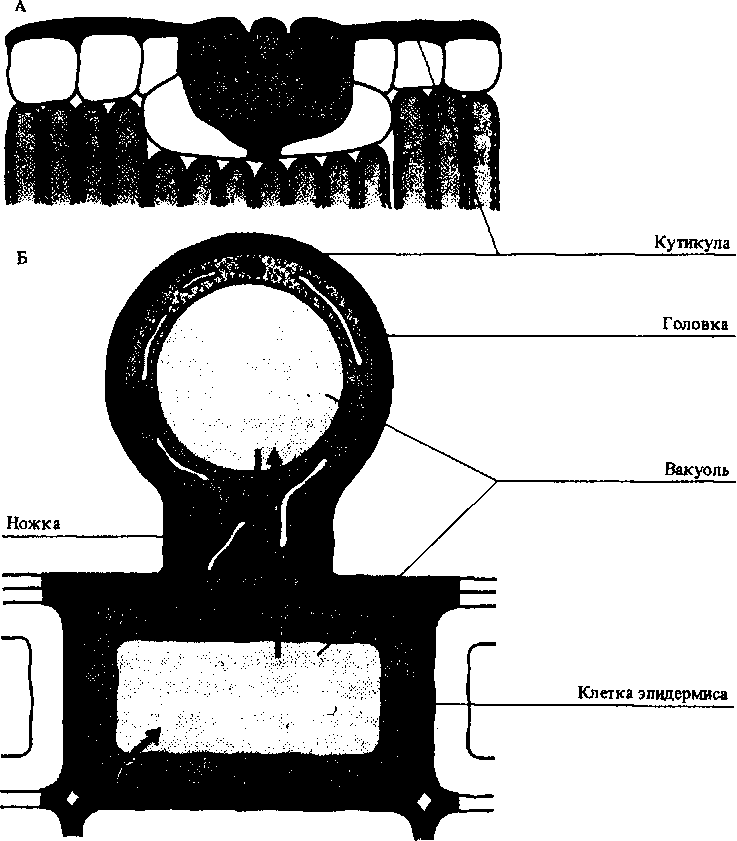
летки,
секретирую-|ие соли. А — солевая ёзка;
Б — солевой Олосок лебеды.
фасными
стрелками
отчем
активный
транс-9рт
ионов

новная масса терпеноидов выводится из клетки, у идиобластов и млечников — накапливается в вакуолях.
Особенность ультраструктуры терпеноидогенных клеток — преимущественное развитие в них агранулярного ЭР, который, как полагают, участвует в синтезе терпеноидов. Для этих клеток характерно также большое количество лейкопластов, митохондрий и липидных капель.
Предполагается, что компоненты эфирных масел и смол могут синтезироваться и накапливаться в агранулярном рети-кулуме, митохондриях, пластидах и даже в ядре. Они выводятся эккриновой секрецией через плазмалемму, поступают в свободное пространство клеточных стенок и оттуда попадают либо в канал смоляного хода, либо (у желёзок) на поверхность растений. Каучук и гутта, по-видимому, остаются в тех клетках, в которых были синтезированы.
Выделяемые растениями эфирные масла, смолы, каучук и гутта являются производными изопрена (С5Н8) и синтезируются в различных клеточных органоидах с участием ацетил-СоА. В состав эфирных масел входят:
а) альдегиды, спирты и эфиры спиртов алифатических терпенов (С10Н16), например спирты линалоол (выделяется цветками ландыша, содержится также в апельсиновом и кориандро
вом масле), гераниол (важный компонент розового масла, масла герани, эвкалипта) и др.;
б) циклические терпены и их производные, например лимонен (в скипидаре, масле укропа, тмина), вторичный спирт — ментол, составляющий 70% эфирного масла перечной мяты, пинен — главный компонент скипидара и др.
Каучук и гутта — полимерные производные изопрена (C5N8). Полиизопреновая цепь каучука может включать 500 — 5000 остатков изопрена, а гутты — около 100. Основной производитель каучука — бразильская гевея; в нашей стране каучуконосами являются кок-сагыз и тау-сагыз, у которых каучук накапливается в "млечниках корней. Гуттаперчу содержат бересклет и субтропическое дерево эвкомия. Гутта сосредоточена в замкнутых вместилищах в коре корней бересклета и в листьях у эвкомии.
Как уже отмечалось, секреты (полисахаридная слизь, раз- Экскреторная личные терпеноиды, смолы и т. д.) могут накапливаться в спе- 1стема растений ^
v циальных вместилищах и млечниках. Секреторные вместилища
(смоляные и слизевые ходы) встречаются во всех частях растения. Они возникают либо путем разъединения ранее плотно прилегающих друг к другу клеток, либо путем растворения самих клеток. В последнем случае полагают, что секрет вырабатывается клетками до их разрушения.
Млечники — это одиночные клетки (нечленистые) или ряды слившихся клеток (членистые), содержащие млечный сок, или латекс. Латекс состоит из жидкой основы, представляющей собой вакуолярный сок млечника (углеводы, органические кислоты, белок, слизи, алкалоиды), и взвешенных в нем углеводородов, относящихся к терпенам (эфирные масла, смолы, каротиноиды, каучук или гутта). У млечников протопласт остается живым и занимает постенное положение. Основной объем клетки заполнен вакуолью с латексом. Как и в ситовидных трубках, в млечниках нет четкой границы между цитоплазмой и вакуолью. Они способны к росту в живых тканях и погибают при отмирании окружающих клеток.
Так как сок из секреторных вместилищ и млечников пассивно вытекает лишь при поранении, эти образования относят к экскреторной системе растений.
Растения выделяют многие вещества как пассивным (экскреция), так и активным способом, т. е. с затратой метаболической энергии (секреция). Процесс секреции осуществляется специализированными клетками и тканями, но присущ также каждой клетке (формирование клеточной стенки, ионные насосы мембран, вторичный активный транспорт). На уровне клетки у растений функционируют те же основные способы выделения веществ, что и у животных. Единой выделительной системы у растений нет. Выделяемые вещества накапливаются внутри клетки (в вакуолях) или в специальных хранилищах (смоляные и слизевые ходы) либо выводятся наружу. Выделения растений играют существенную роль в поддержании гомеостаза клеток самого растения, а также при формировании почвенных и наземных фитоценозов, в сопряженной эволюции растений с другими организмами, например с насекомыми.
