

2)Гипотезы о происхождении стеблей (осей) и листьев высших растений.
Теломная теория
Время: с 30-х годов XX столетия. Основатель: В. Циммерман.
Согласно этой теории, все органы высших растений происходят и независимо развиваются из теломов; высшие растения с настоящими корнями и побегами происходят от риниофитов, тело которых было представлено системой дихотомически ветвящихся простых цилиндрических осевых органов — теломов и мезомов. В ходе эволюции в результате перевершинивания, уплощения, срастания и редукции теломов возникли все органы покрытосеменных растений. Листья семенных растений возникли из уплощённых и сросшихся между собой систем теломов; стебли — благодаря боковому срастанию теломов; корни — из систем подземных теломов. Основные части цветка — тычинки и пестики — возникли из спороносных теломов и эволюционировали независимо от вегетативных листьев.
О происхождении листьев существуют различные гипотезы.
В последнее время наибольшим вероятием пользуется теория, основанная на изучении
строения первичных сухопутных растений - псилофитов, живших в силурийском и
девонском периодах геологической истории Земли.
Впервые они (роды риния, хорнеофитон, астероксилон) были описаны в 1917 г. В
настоящее время известно более 20 родов псилофитов, найденных в отложениях различных континентов. У этих растений не было ни корней, ни
листьев, и все вегетативное тело их состояло из подземной горизонтальной оси, напоминавшей корневище, и отходящих от нее надземных, дихотомически
разветвленных небольших цилиндрических осевых органов, подобных безлистному
стеблю и названных теломами. Концы некоторых разветвлений теломов заканчивались
спорангиями. У некоторых псилофитов (астероксилон) теломы были обильно
покрыты мелкими щетиновидными или чешуйчатыми выростами. В дальнейшем эволюция этих растений пошла в разных направлениях. У одних маленькие выросты на теломах увеличились в размерах и приняли более или менее плоскую форму, более удобную для фотосинтеза. В них развился проводящий пучок, соединенный с проводящей тканью осевого органа. Это направление эволюции привело к мелколистным высшим споровым растениям - плауновидным и хвощевидным, листья которых, следовательно, развились из мелких выростов на осевых органах, так называемых энациев.
Вторая линия эволюции, приведшая к крупным листьям (как, например, у
папоротников), была совершенно иная. Здесь листья возникли из нескольких осевых
дихотомических ветвей - теломов - путем перемещения их в одну плоскость, уплощения
(изменения радиального строения в дорзовентральное), сращения боковыми частями,
утраты способности к неограниченному росту в длину. Листья этого типа по своей
морфологической природе подобны кладодиям, имеют кладодификационное происхождение.
5. Функции корня. Типы корней и развитие корня в онтогенезе растения. Морфология корня. Заложение и развитие боковых корней.
Корень – основной вегетативный орган растения, выполняющий в типичном случае функцию почвенного питания. Корень – осевой орган, обладающий радиальной симметрией и неопределенно долго нарастающий в длину благодаря деятельности апикальной меристемы. От побега он морфологически отличается тем, что на нем никогда не образуются листья, а апикальная меристема всегда прикрыта корневым чехликом.
Кроме главной функции поглощения веществ из почвы, корни выполняют и другие функции:
1) корни укрепляют («заякоривают») растения в почве, делают возможным вертикальный рост и вынесение побегов наверх;
2) в корнях синтезируются различные вещества, которые затем передвигаются в другие органы растения;
3) в корнях могут откладываться запасные вещества;
4) корни взаимодействуют с корнями других растений, микроорганизмами, грибами, обитающими в почве.
Совокупность корней одной особи образует единую в морфологическом и физиологическом отношении корневую систему.
В состав корневых систем входят корни различной морфологической природы – главный корень, боковые и придаточные корни.
Главный корень развивается из зародышевого корешка. Боковые корни образуются на корне (главном, боковом, придаточном), который по отношению к ним обозначается как материнский. Они возникают на некотором расстоянии от апекса, в направлении от основания корня к его верхушке. Боковые корни закладываются эндогенно, т.е. во внутренних тканях материнского корня. Если бы ветвление происходило в самом апексе, это бы затруднило продвижение корня в почве. Придаточные корни могут возникать и на стеблях, и на листьях, и на корнях. В последнем случае они отличаются от боковых корней тем, что не обнаруживают строгого порядка заложения вблизи апекса материнского корня и могут возникать на старых участках корней.
По происхождению выделяют следующие типы корневых систем ( рис. 4.1):
1) система главного корня представлена главным корнем (первого порядка) с боковыми корнями второго и последующих порядков (у многих кустарников и деревьев, большинства двудольных растений);
2) система придаточных корней развивается на стеблях, листьях; встречается у большинства однодольных растений и многих двудольных, размножающихся вегетативно;
3) смешанная корневая система образована главным и придаточными корнями с их боковыми ответвлениями (многие травянистые двудольные).
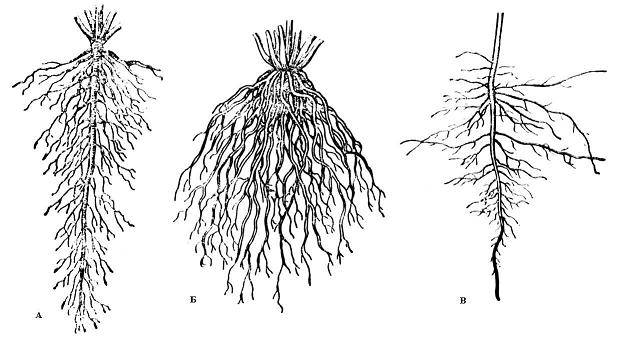
Рис. 4.1. Типы корневых систем : А – система главного корня; Б – система придаточных корней; В – смешанная корневая система (А и В – стержневые корневые системы; Б – мочковатая корневая система).
По форме различают стержневую и мочковатую корневые системы.
В стержневой корневой системе главный корень сильно развит и хорошо заметен среди остальных корней. В мочковатой корневой системе главный корень незаметен или его нет, а корневая система составлена многочисленными придаточными корнями ( рис. 4.1).
Корень обладает потенциально неограниченным ростом. Однако в естественных условиях рост и ветвление корней ограничены влиянием других корней и почвенных экологических факторов. Основная масса корней располагается в верхнем слое почвы (15 см), наиболее богатом органическими веществами. Корни деревьев углубляются в среднем на 10-15 м, а в ширину распространяются обычно за пределы радиуса крон. Корневая система кукурузы заходит на глубину около 1,5 м и примерно на 1 м во все стороны от растения. Рекордная глубина проникновения корней в почву отмечена у пустынного мескитового кустарника – более 53 м.
У одного куста ржи, выращенного в теплице, общая длина всех корней составила 623 км. Суммарный прирост всех корней за одни сутки равнялся примерно 5 км. Общая поверхность всех корней у этого растения составила 237 м2 и была в 130 раз больше поверхности надземных органов.