

18.Естественные киллеры: механизмы активации, функции
ЕК – это клетки естественного врожденного иммунитета, самостоятельная популяция лимфоцитов. Возникают из костно-мозговых предшественников под влиянием ГМ-КСФ и ИЛ-2. Представляют собой крупные гранулярные лимфоциты (5-15% среди лимфоцитов в крови, много в печени и селезенке), имеющие почковидное ядро и азурофильные гранулы в цитоплазме.
Эти клетки не имеют специфического рецептора к антигену. Однако они разрушают клетки-мишени особенно инфицированные вирусом или опухолевые (неиммунный цитолиз). Выделяют цитотоксический белок перфорин (сходен с МАК комплемента), ФНОbb, ферменты гранзимы. Естественные киллеры вызывают апоптоз посредством взаимодействия рецептора апоптоза CD95 с Fas-лигандом этих клеток.
В процессе цитолиза различают 3 основных стадии: распознавание, выделение цитотоксинов ("летальный удар") и лизис клетки-мишени. ЕК участвуют в разрушении опухолевых клеток, могут лизировать и некоторые виды бактерий. Основные маркеры ЕК – CD16 и CD56.
В норме на ЕК-клетках находятся как киллинг-ингибирующие, так и киллинг-активирующие рецепторы. В отсутствие патологии ингибиторные рецепторы реагируют с HLA I класса собственных клеток и тем самым подавляют активацию ЕК. К лизису естественными киллерами становятся чувствительными клетки, утратившие или изменившие антигены HLA I класса, например в результате инфицирования вирусом.
В то время, как Т-киллеры распознают чужеродные антигены, связанные HLA молекулами I класса (измененное «свое»), а Т-хелперы – пептиды в комплексе с НLA II класса гистосовместимости, ЕК лизируют клетки, не имеющие таких молекул – маркеров "своего".
Дефициты функций ЕК служат причиной вирусных, особенно герпетических инфекций и, возможно, развития опухолей.
19.Система комплемента
Комплементом называют сложную систему ферментативных и рецепторных белков (более 30) сыворотки крови. Основные 13 компонентов системы комплемента обозначаются буквой C с соответствующим номером (C1, С2, С3 и т.д.) Они образуются в печени и секретируются макрофагами. Активация системы комплемента протекает классическим, очень сходным с ним лектиновым, а также альтернативным путями. Процесс имеет вид цепной реакции, управляемой регуляторными белками. При этом каждый предыдущий компонент каскада активирует несколько последующих за счет их ферментативного расщепления.
При распаде компонентов комплемента обычно образуется 2 фрагмента. Больший фрагмент обозначается буквой «b» и является активным, продолжая каскад расщепления. Меньшие фрагменты в дальнейшей активации комплемента обычно не участвуют, однако проявляются многообразными биологическими функциями. Они обозначаются буквой «а». Исключение из этого правила – фактор С2. Комплексы активированных компонентов обозначаются сверху чертой.
Белки альтернативного пути активации получили название факторов и обозначаются большими латинскими буквами (В, Н, I и т.д.)
Среди регуляторных белков различают естественный ингибитор компонента комплемента С1 (С1-ингибитор), который тормозит спонтанную активацию C1q компонента. При его дефиците возникает наследственный ангионевротический отек. Кроме этого существует фактор DAF, ускоряющий деградацию С3b компонента комплемента на мембранах собственных клеток организма, предотвращая их лизис.
Классический путь активации запускается комплексом антиген-антитело в присутствии катионов Ca и Mg обычно на поверхности клетки-мишени (рис.1) Эффективными активаторами данного пути являются АТ классов IgG и IgM. Комплекс «антиген-антитело» связывается с компонентом С1q, который присоединяет С1rs, а затем активирует и расщепляет С4 на С4а и С4b . С4b присоединяется либо к С1, либо к поверхности клетки-мишени. Далее к нему присоединяется С2. Он, в свою очередь, расщепляется на С2а и С2b предыдущим компонентом. С2а остается связанным с С4b. Этот комплекс получил название конвертазы классического пути активации комплемента. Она расщепляет С3 компонент на С3а и С3b. С3b присоединяется к конвертазе классического пути, образуется конвертаза С5 компонента, и этот макромолекулярный комплекс активирует компонент С5. Он распадается на С5а и С5b. К С5b на мембране клетки-мишени последовательно присоединяются С6, С7, С8. Этот комплекс встраивается в мембрану клетки-мишени и к нему может присоединиться до 20 молекул С9 компонента.
Са++
Mg++

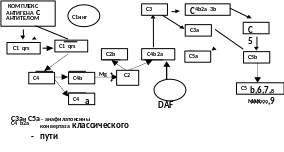
Рис. 1.1. Классический путь активации комплемента
Комплекс С5b-C9 получил название мембраноатакующего комплекса (МАК). В механизме его литического действия много общего с цитотоксическим белком перфорином. МАК встраивается в мембрану клетки-мишени за счет гидрофобных взаимодействий, образуя трансмембранный канал. Через него в клетку поступают ионы натрия и вода, а выходят ионы калия, что приводит к цитолизу.
Для того чтобы МАК не разрушал собственные клетки организма, его образование может быть блокировано растворимыми сывороточными факторами (S-фактор – белок витронектин). Кроме того, имеющийся на мембранах клеток рецептор CD59 препятствует присоединению к МАК компонента С9.
Лектиновый путь активации комплемента отличается только природой иммунного комплекса, запускающего начальный этап активации. Со стороны иммунной системы здесь участвуют белки-лектины, связывающие полисахаридные компоненты бактерий (например, маннансвязывающий лектин, С-реактивный белок и т.д.) Активация происходит через компонент С1, и ее ход не отличается от классического пути.
Альтернативный путь активации комплемента (рис. 1.2) является неспецифическим.
Центральным звеном альтернативного пути является С3b компонент. Следовые его количества постоянно присутствуют в сыворотке вследствие спонтанного гидролиза С3. Этот процесс запускается и усиливается липополисахаридами клеточной стенки бактерий (эндотоксинами), агрегированными иммуноглобулинами, лекарственными препаратами и т.д. Образующийся при этом С3b-компонент в присутствии ионов магния связывается с фактором В сыворотки (неактивная сериновая протеаза). На комплекс С3bB действует фактор D – активная сывороточная протеаза. Она расщепляет фактор В на Ва и Вb. Образующийся комплекс С3bBb представляет собой конвертазу альтернативного пути активации. В норме она неустойчива, но стабилизируется белком пропердином (белок P). Конвертаза альтернативного пути присоединяет еще одну молекулу С3b, образуется конвертаза С5 компонента С3bBbС3b, которая активирует С5. Дальнейшая активация комплемента не отличается от классического пути. Таким образом, С3-компонент является ведущим в активации комплемента по обоим путям, определяя процессы цитолиза.
В процессе активации комплемента образуются биологически активные фрагменты. Так, компоненты С3а, С4а и С5а служат анафилатоксинами, действуя на макрофаги, гранулоциты, тучные клетки. Они вызывают выделение из них медиаторов, дегрануляцию тучных клеток. Возникающий патологический процесс клинически проявляется аллергическими (шок и др.), псевдоаллергическими реакциями, воспалением и повреждением тканей.
При заболеваниях, сопровождающихся образованием иммунных комплексов (аутоиммунные болезни, инфекции), уровень белков комплемента снижается – гипокомплементемия. Уровень комплемента наиболее высок у морских свинок, поэтому их сыворотка крови используется как "комплемент" в серологических реакциях.
Компоненты активированного комплемента связываются с рецепторами комплемента, имеющимися на лейкоцитах. Основной рецептор CR1 (CD35) связывает C3b, кроме того, рецепторами к компонентам комплемента являются лейкоцитарные интегрины. Взаимодействуя с этими рецепторами клеток продукты активации комплемента стимулируют функции лейкоцитов, запускают воспаление; усиливают противомикробный иммунитет.
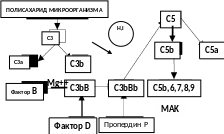
Рис. 1.2. Альтернативный путь активации комплемента