

Глава 4 эколого-биологические последствия чернобыльской катастрофы
4.1. Особенности биологических эффектов радиации низкой эффективности Установлено, что радиация в достаточно больших дозах оказывает повреждающее действие на живые системы (ЖС) различного уровня организации. Небольшие же дозы радиации, как полагали (и считают до настоящего времени некоторые исследователи), не вызывают поражений, оказывают стимулирующие биологические эффекты (БЭ), либо организмы, находясь достаточно длительное время в радиационном поле, адаптируются к новым условиям существования без проявлений каких-либо нефизиологических изменений или фатальных отдаленных последствий. Таким образом, предполагается существование порога вредного действия радиации на здоровье. В пользу этого положения говорят данные об отсутствии неблагоприятных изменений у животных и человека, обитающих в условиях постоянного повышенного фона радиации и дополнительной добавки к нему излучений от искусственных источников, а- также большая практика относительно безвредного применения рентгеновских диагностических мероприятий. Анализ результатов многочисленных исследований показывает, что риск проявления отрицательных отдаленных последствий облучения радиации низкой интенсивности в области величины доз, находящихся ниже предполагаемого порога вредного действия и считавшихся безопасными, возрастает на несколько порядков по сравнению с ожидаемыми результатами при экстраполяции БЭ из области больших доз излучений высокого уровня. По современным представлениям все виды радиации, независимо от природы, в конечном итоге разменивают свою энергию, взаимодействуя с электронной оболочкой .атомов, и приводят, таким образом, к их ионизации и возбуждению. Существенное отличие действия различных видов радиации заключается лишь в плотности распределения энергии в единице объема. Так, бета-частицы, или электроны, образованные R- или гамма-квантами, приводят к низкой, а альфа-частицы - к очень высокой плотности ионизации. В зависимости от этого количество взаимодействий в данной области, где возможно наличие чувствительных мишеней, БЭ имеют существенные количественные отличия. Повреждение биологически важных структур клетки на физико-химическом этапе действия радиации может быть реализовано двумя способами. В первом способе электроны, образовавшиеся в процессе размена энергии, как правило, приводят к разрыву молекулярных связей той структуры, которая является мишенью. Благодаря этому возникают одно- и двунитевые разрывы молекул ДНК, с определенной вероятностью детерминирующие необратимые повреждения в геноме клетки. Такого рода повреждения линейно зависят от плотности ионизации, дозы излучения, типичны для средних и высоких значений доз и имеют порог действия, величина которого определяется степенью надежности данной ЖС, в первую очередь состоянием ее репаративных систем, и составляет 10 - 100 сГр. Однако при определенных обстоятельствах более опасным по удельному вкладу может оказаться второй способ взаимодействия ионизирующей радиации с ЖС, при котором повреждение биологических структур происходит за счет очень реакционно-способных химических агентов, образующихся в местах размена энергии частицы или кванта, способных мигрировать на значительные расстояния и приводить к повреждению структур-мишеней. Одним из основных активных химических групп, количество которых значительно увеличивается при взаимодействии радиации с ЖС, являются свободные радикалы (СР) кислорода (супероксидный анион -· O2 -, гидроксильный анион -· OH-, другие радикалы - · Н-, · Н2О+, · ОН, · О) и Н2О2 .Наиболее реакционно-способными из них являются · O2 -, · OH-и Н2О2. СР в небольших количествах образуются и при физиологических процессах в ходе энзиматических реакций, выступают в качестве активаторов клеточной пролиферации, митогенеза, дифференциации, участвуют в синтезе простагландинов, лейкотропинов. От содержания некоторых из них зависят физиологическая активность клеток и другие функции. В то же время благодаря высокой химической активности СР при определенных условиях легко вступают в реакции с основными биологическими соединениями, нарушая их структуру и функции. Наибольшего внимания заслуживает радикал · O2 -. Он образуется при захвате электрона молекулой растворенного в среде кислорода и может возникать при неферментативном окислении катехоламинов, тиолов, гидрохинона, ферредоксина и ферментативных процессах, катализируемых альдегидоксидазой, ксантиноксидазой, дигидрооротатоксидазой и др. · O2 -возникает также в системе НАДФН - цитохром С. · O2 -высокотоксичен, его химическая агрессивность выше в гидрофобной фазе, чем в водной, поэтому его повреждающее действие, как предполагается, особенно существенно в гидрофобных зонах мембранных комплексов. При участии супероксиддисмутазы (СОД) радикалы · O2 --дисмутируют до Н2О и О2. Гидроксильные радикалы · OH-инициируются в живых системах при действии ионизирующей радиации в значительных количествах, они проявляют большое сродство к электрону. Полагают, что радикал · OH-является одним из основных агентов свободнорадикальных процессов окисления липидов (СРПОЛ). Н2О2образуется в значительных количествах в ходе химических реакций, инициируемых радиацией, диффундирует на значительные расстояния, принимает активное участие в окислении органических молекул. В ЖС, подвергаемой облучению, очень вероятна реакция Хабера - Вайсса с большим выходом радикалов · OH-: Н2О2+ · O2 -> · OH-+ · OH-+ О2 Н2О2и образовавшиеся СР вызывают повреждения всех биологических молекул: разрывы в ДНК, окисление SH-групп, сшивки полипептидных цепей в белках, инактивацию ферментов, полимеризацию углеводов. И, пожалуй, наиболее трагичным для судьбы клетки процессом является инициирование ими цепей СРПОЛ. Нормально функционирующие ЖС обладают эффективными средствами защиты от СР. Первая из них - металлоферментные системы - СОД, глютатионпероксидаза и каталаза. СОД встречается в разных формах, активный центр каждой из ее субьединиц содержит атом меди, цинка или марганца, не участвующих в реакции, но стабилизирующих структуру фермента. Сu- и Zn-СОД локализованы в цитозоле и кристах митохондрий, Мn-СОД - в матриксе митохондрий. СОД в норме составляет почти 0,1 % общего белка протоплазмы, является специфическим инактиватором радикалов · O2 -. Каталаза локализована в основном в пероксисомах и содержит по одной группе гема. Глютатионпероксидаза содержит в активном центре селен и при участии глютатиона ускоряет разложение Н2O2. Вторым эффективным средством защиты ЖС от СР являются антиоксиданты (АО), которые, легко окисляясь, инактивируют СР. Свойствами АО обладают вещества различной химической природы: токоферол, каротин, глютатион, аскорбиновая кислота, цистеин, белки, содержащие SH-группы, и др. Защитным от активных СР свойством обладают также фосфолипиды мембранного комплекса, в основном за счет их ненасыщенных углеводородных связей. В настоящее время установлены детерминированные связи между различными показателями интенсивности метаболизма, содержанием СР и активностью ферментов-детоксикаторов. В частности, показано, что при интенсивном развитии некоторых процессов указанные ферментные системы не справляются с инактивацией большого количества радикалов. Установлено исключительно важное значение СРПОЛ в повреждении клеток при действии различных физических, химических факторов и их роль в развитии патологических процессов в организме. Стало известным, что интенсификация СРПОЛ связана с уменьшением активности АО ферментов из-за их инактивации, которая может быть причинно обусловлена, в частности, ацидозом на начальном этапе повреждения клеток. При снижении, например, рН с 7,5 до 6,8 активность глютатионпероксидазы снижается в 10 раз. Установлено, что · O2 -- инактивирует каталазу, Н2О2 -СОД. Таким образом, СР и инициируемые ими СРПОЛ - очень агрессивные агенты, которые повреждают биологически важные молекулы и структуры. При этом стабилизирующими защитными факторами выступают специализированные ферментные и АО системы. Описанные выше реакции с участием СР являются основными повреждающими факторами для клеточных мембран. Анализ значительного количества литературных данных позволяет прийти к выводу о существенном значении непрямого механизма действия радиации низких интенсивностей. Особенности его заключаются в следующем. С уменьшением интенсивности (плотности) излучения вероятность повреждения клетки увеличивается, при этом механизме действия чувствительными мишенями являются мембраны. Согласно этому, чем выше интенсивность излучения, тем выше плотность ионизации и тем меньше вероятность достижения образовавшимися активными СР чувствительных мишеней - мембран клетки. С одной стороны, известно, что чем выше плотность ионизации, тем больше актов рекомбинации СР. С другой стороны, плазматическая мембрана клетки обладает электрическим полем [132], притягивающим отрицательно заряженные СР. Показано, что с увеличением плотности радикалов величина электрического поля уменьшается [132]. Следовательно, при действии радиации высокой интенсивности создаются менее благоприятные условия для достижения СР чувствительных мишеней клетки, чем при радиации низкой, близкой к фоновой, интенсивности. Эффект различия наиболее ярко выражен при длительном действии радиации малой интенсивности. Таким образом, длительное воздействие радиации низкой интенсивности при данной полной дозе До (До - величина дозы, при которой происходит удвоение средней нормы генетических дефектов - разрывов мембран или уровня других патологических процессов) представляет большую опасность для мембран, чем однократное облучение радиации большой интенсивности. Отсюда следует очень важный вывод о некорректности сравнения биологической эффективности различных по качеству видов радиации, исходя только из существования зависимости "доза-эффект", без учета интенсивности излучения. Повторяем, что речь идет о малых дозах радиации низкой интенсивности. При значительных дозах и высоком уровне радиации имеет место прямое повреждение генетических структур, а именно они преимущественно в этом случае являются чувствительными мишенями, эффект прямо пропорционален плотности ионизации. Анализ с этой точки зрения результатов исследований показал, что чувствительность ЖС к действию радиации близких к фоновым интенсивностей резко увеличена, с дальнейшим ростом уровня излучения выходит на насыщение и, таким образом, проявляет логарифмическую зависимость. Это позволяет представить [133 - 135] объективную картину радиобиологических эффектов, имеющих место при воздействии на ЖС ионизирующей радиации широкого диапазона доз и интенсивностей. Согласно этому, под влиянием ионизирующей радиации в любой ЖС одновременно реализуются оба механизма взаимодействия - прямой и непрямой, удельный вклад которых в суммарный БЭ зависит от дозы радиации и ее интенсивности. В диапазоне больших доз и интенсивностей радиации превалируют линейно зависимые от дозы повреждения ДНК, До с уменьшением ее уровня возрастает, поскольку все с большей эффективностью включаются репаративные восстановительные системы. Это подтверждается многочисленными данными, в частности, о генетических мутациях, риске опухолевых заболеваний у животных и человека. С ростом уровня радиации величина дозы, при которой генетические повреждения являются причиной неопластической трансформации клеток, незначительно уменьшается. Это свидетельствует о том, что повреждение генетического материала детерминировано прямым попаданием, при котором количество нерепарируемых или ошибочно репарируемых, в основном двунитевых разрывов ДНК, может увеличиваться с ростом интенсивности излучения. В этом диапазоне уровней доз имеет место и непрямой путь взаимодействия, однако его вклад в эффект из-за описанных выше интенсивных рекомбинационных для СР процессов и значительно выраженных последствий, связанных с повреждением структур ДНК-мишеней, неощутим, и для его проявления требуются дозы Д0~ 103- 107сГр. В диапазоне низких интенсивностей излучений восстановительные системы "успевают" достаточно надежно защитить генетический материал от прямого действия радиации, и для проявления биологического эффекта при этом виде взаимодействия у объектов, обладающих достаточно мощной системой восстановления, требуются дозы Д0~ 104- сГр. В то же время косвенные повреждения при данных интенсивностях радиации имеют очень низкие Д0, достигающие единиц или десятых долей сГр. При таких условиях облучения при доминирующем повреждении плазматических мембран предполагается значительное увеличение эволюционных дефектов из-за нарушения процесса размножения клеток, что может проявиться в увеличении количества рожденных с дефектами, самопроизвольных выкидышей, смертей за счет плохого развития, гормональной недостаточности и иммунодефицита. Последнее может явиться причиной снижения сопротивляемости организма к опухолевому росту [130].
4.2. Биологические эффекты у животных 4.2.1. Изменения биофизических и биохимических показателей жизненно важных систем организма Воздействие на людей комплекса радиационных факторов, обусловленных Чернобыльской катастрофой, можно несколько условно разделить на три вида в зависимости от мощности дозы облучения и его продолжительности. А. Острое лучевое поражение явилось следствием кратковременного (часы, сутки) эффекта комплекса радиационных воздействий большой мощности, создававшего дозу тотального, более или менее равномерного, облучения - 1 - 1,2 Гр и выше (острая лучевая болезнь I, II и III степени) либо 0,5 - 1,0 Гр (острая лучевая реакция). Б. Подострое лучевое поражение как следствие работы в 30-километровой зоне ЧАЭС или пребывание в ней до момента эвакуации - в течение нескольких суток, недель; дозы, полученные этим контингентом за этот срок, могут быть весьма приблизительно оценены в пределах 25 - 100 сГр и более. В. Хроническое (постоянное) облучение на протяжении месяцев и лет, обусловленное пребыванием (проживанием) на радиоактивно загрязненных территориях в 30-километровой зоне ЧАЭС и за ее пределами. Лучевые нагрузки, полученные этой категорией пострадавших, еще в меньшей степени поддаются количественной оценке как в силу несовершенства дозиметрии, так и ввиду сложности учета лучевых нагрузок, создаваемых внутренним облучением различными радионуклидами. Объем популяции А - порядка 150 человек, популяции Б - десятки тысяч, популяции В - сотни тысяч человек. Первую популяцию образовали главным образом работники АЭС и пожарные, пострадавшие во время аварии и в первые часы и сутки после нее. В лучевом поражении у них доминировало внешнее гамма- и бета-облучение, как правило, в сочетании с ожогами. Вторую популяцию составили эвакуированные из 30-километровой зоны и непосредственные участники ликвидации аварии ("ликвидаторы") 1986 г. Третью - главным образом жители районов, для которых установлен режим "жесткого контроля", и "ликвидаторы" 1987 - 1988 гг. По мере увеличения продолжительности и уменьшения мощности дозы радиации в этих группах пострадавших возрастал удельный вес внутреннего облучения за счет поступления комплекса радионуклидов с пищей, водой, а в первые месяцы и при выполнении сельскохозяйственных работ - респираторным путем. Экспериментальный анализ этих вариантов лучевого поражения осуществлялся путем моделирования на лабораторных животных. Экспериментальная модель острого лучевого поражения, соответствующего первому варианту, была хорошо изучена и до Чернобыльской катастрофы, в частности, костномозговой синдром. Биохимические и биофизические исследования, выполненные в 1987 - 1993 гг., позволили существенно углубить представления, сложившиеся ранее, и обосновать некоторые новые подходы к коррекции этих изменений. В частности, были изучены закономерности нарушения окислительного гомеостаза, особенности эффектов, создаваемых комбинацией внешнего и внутреннего облучения, ионизирующей и ультрафиолетовой радиацией, а также нерадиационных факторов. Наибольшую ценность представляют данные, полученные при моделировании подострого и хронического облучения в относительно низких и малых дозах, в частности, касающиеся отдаленных последствий такого лучевого воздействия. Установлено, что по мере снижения мощности дозы и суммарной дозы радиации относительно все большее значение приобретают в расчете на единицу дозы явления угнетения иммунореактивности (снижение общего количества лейкоцитов, относительная и абсолютная лимфопения со снижением количества и. активности Т-лимфоцитов, В-лимфоцитов и клеток-киллеров), тромбоцитопения и анемия. Активация перекисного окисления липидов (ПОЛ), выражающаяся в увеличении интенсивности спонтанной хемилюминесценции (СХЛ) сыворотки крови и суспензий клеток радиочувствительных органов лабораторных животных, а также содержания ранних продуктов ПОЛ (гидроперекисей, диеновых и триеновых коньюгатов) наблюдалась непосредственно после острого лучевого поражения и в первые часы и сутки подострого и хронического облучения. В более поздние сроки возрастало содержание поздних и конечных продуктов ПОЛ - малонового диальдегида (МДА) и шиффовых оснований (ШО). Активация ПОЛ обусловливает развитие ответной реакции со стороны физиологической антиоксидантной (АО) системы клеток и тканей, а также системной стресс-реакции. АО-потенциал крови и радиочувствительных органов реактивно возрастает за счет увеличения активности АО-ферментов супероксиддисмутазы (СОД),каталазы. глютатионпероксидазы и глутатионредуктазы клеток и церулоплазмина крови; высвобождения из внутриклеточных и тканевых депо и перераспределения жирорастворимых (токоферол, ретинол, каротиноиды) и водорастворимых (глютатион, другие низкомолекулярные тиолы, аскорбиновая кислота) АО. В результате на определенный срок (несколько суток для варианта А, недель для варианта Б и месяцев для В) наблюдается реактивное подавление ПОЛ, снижение уровня его продуктов (иногда даже ниже исходного), свидетельствующее о сохранности защитных механизмов. Однако со временем, по мере усиленного расходования АО-резервов, наблюдается снижение активности СОД, глютатионпероксвдазы, редукгазы, содержания токоферола и аскорбата и возрастание интенсивности ПОЛ. Вторичная активация ПОЛ является фактором повреждения клеток (прежде всего радиочувствительных) и их гибели, характеризует вступление лучевого поражения в период развернутых клинических проявлений. Следует особо подчеркнуть, что относительная АО-недостаточность алиментарного происхождения, предшествующая лучевому поражению или сопутствующая ему (как было в первые месяцы и годы после Чернобыльской катастрофы из-за ограничения потребления молочных продуктов и фруктов, овощей, а также из-за сезонных вариаций питания), весьма отрицательно сказывается на радиорезистентности организма, ускоряя истощение АО-потенциала, увеличивая тяжесть лучевых изменений. Низкое потребление молока, снижая поступление биологически активного кальция, оказывает неблагоприятное влияние, увеличивая проницаемость и снижая прочность стенок сосудов (кровоточивость) и усугубляя метаболические нарушения. Известно, что после Чернобыльской катастрофы лучевые поражения, как правило, сочетались с ожогами, высокой интенсивностью физического труда, психоэмоциональным напряжением (варианты А и Б), алиментарной АО-недостаточностью, под острым и хроническим психоэмоциональным стрессом, обусловленным страхом перед невидимой опасностью, угрозой отселения, изменением условий проживания и работы и т.п. (варианты Б и В). Экспериментальное моделирование этих условий путем комбинированного лучевого воздействия и физических нагрузок (плавание, бег на третбане), стресса (иммобилизации, гипер- и гипотермии), механической и физической травмы позволило установить интересные и практически важные закономерности. Оказалось, что предшествующий острому лучевому воздействию слабый стресс оказывает небольшое, но достоверное противолучсвое действие - очевидно, в результате предварительной мобилизации защитных систем (АО, репаративной). Одновременное воздействие более или менее длительного облучения и хронического стресса (например, ежедневной иммобилизации по 2 - 3 ч) существенно отяжеляет состояние животных, углубляя индуцированные радиацией изменения структуры и функций органов. Важны данные о минимальных дозах ионизирующей радиации, вызывающих достоверную активацию ПОЛ. Специальные исследования в условиях тотального внешнего фракционированного облучения крыс показали, что достоверная (на 17 - 25 %) активация ПОЛ, тестируемая по содержанию в крови МДА и других продуктов, наблюдается после суммарной дозы 1 Гр. Меньшие суммарные дозы также вызывают колебательную реакцию показателей ПОЛ, но амплитуда колебаний приближается к Физиологической. Активация ПОЛ в мембранах клеток животных, облученных малыми дозами и интенсивностями, проявляется прежде всего в повышении проницаемости мембран (главным образом пассивной) для электролитов и метаболитов; в снижении резистентности эритроцитов к гемолизу; увеличении активности в крови печеночных трансфераз и содержания метгемоглобина крови при снижении уровня трансферрина и его насыщенности железом. Очевидно, что это одна из причин увеличения частоты железодефицитных анемий у детей из контингентов радиационного риска. Наряду с активацией неферментативного ПОЛ под влиянием низкоинтенсивного воздействия радиации возрастает содержание свободных жирных кислот в результате сопряженной с ПОЛ активации фосфолипазного гидролиза. За счет активации цикло- и липоксигеназ возрастает, но в разной степени, продукция эйкозаноидов, оказывающих различное влияние на радиорезистентность клеток. Установлено, в частности, что при нелетальных дозах радиации снижается продукция кардио- и вазопротектора простациклина и возрастает в определенные сроки содержание тромбоксанов, что может способствовать тромбообразованию и развитию атеросклероза, а в конечном счете - сокращению продолжительности жизни. Кровеносные сосуды и их нейрогуморальный регуляторный аппарат являются одной из мишеней лучевого, в частности, низкоинтенсивного, поражения организма. Первоначальные изменения носят чисто функциональный характер (дискоординация), затем обусловливают развитие нейроциркуляторных, вегетативно-сосудистых дистоний. В основе типичных для последствий Чернобыльской катастрофы церебральных и цереброваскулярных нарушений лежат, в частности, дискоординация активностей ферментов; изменения содержания катехоламинов и их воздействия на нейроны через аденилатциклазную систему [10]. Радиация в дозах 0,6 - 1,0 Гр уже вызывает достоверные, хотя и обратимые, изменения характера спонтанной электрической активности нейронов гиппокампа и других подкорковых структур [95]. Наиболее типичны частые синхронно-билатеральные высоко-амплитудные пароксизмы в a -. Q-, реже D-диапазонах, вплоть до эпилептиформной активности, а также цереброваскулярные нарушения, от вегетососудистой дистонии до дисциркудяторной энцефалопатии [69]. При затравке крыс радионуклидами137Cs и90Sr в течение 1 - 3 мес (суммарная поглощенная доза 0,03 - 0,1 Гр) установлена закономерная активация ПОЛ в ткани головного мозга с нарушением структуры мембран нейронов и глиоцитов, особенно в гипоталамусе, с развитием дистрофически-деструктивных процессов по типу энцефалопатии [95]. Развитие аутоиммунных реакций к антигенам мозга усиливает гибель части клеток мозговой ткани даже при столь незначительных лучевых нагрузках [69]. В клетках иммунной системы длительное низкоинтенсивное лучевое воздействие также индуцирует комплекс биохимических и биофизических сдвигов. Усиление спонтанной и люминолзависимой хемилюминесценции нейтрофилов, макрофагов, мононуклеаров наряду с активацией ферментов цикла Кребса, гликолиза, а также щелочной фосфатазы и пероксидазы свидетельствует о первоначальном усилении функции этих клеток. Одновременно несколько возрастает продукция сывороточных Р-белков, активность церулоплазмина и других белков. С увеличением длительности радиационного воздействия начинают доминировать явления угнетения функции иммуноцитов, нарушается соотношение субпопуляций Т-лимфоцитов. Отмечается закономерный рост аутоимунных реакций. Следует подчеркнуть преимущественно неспецифический характер развивающихся изменений, их главным образом реактивную природу как компонентов клеточного и системного стресса. Низкая интенсивность и небольшая суммарная доза лучевого воздействия как будто предопределяют обратимость возникающих нарушений. Однако продолжительность воздействия и наличие нерепарируемой компоненты, вялая мобилизация защитно-компенсаторных и репаративных механизмов способствуют формированию хронического лучевого стресса с исходом в фазу истощения и гибелью части клеточной популяции радиочувствительных органов, головного мозга, элементов сосудистой стенки, миокарда. Другой опасностью этого состояния является формирование таких отдаленных последствий лучевого поражения, как сокращение продолжительности жизни, возникновение радиогенных лейкозов и солидных опухолей, более раннее развитие атеросклероза, нефросклероза, пострадиационной энцефалопатии. Особо следует подчеркнуть, что по мере снижения мощности дозы и суммарной дозы радиации все более важную роль приобретает комбинированное воздействие радиации и факторов нерадиационной природы. В этих комбинированных эффектах важно выявление общих патогенетических звеньев, воздействие на которые способно дать взаимное усиление эффектов. К числу таких общих звеньев наряду с активацией ПОЛ относится повреждение ДНК. Сочетание ионизирующего воздействия с факторами, способными угнетать активность ферментов репарации, способно усилить повреждение ДНК. Так влияет ультрафиолетовое (УФ) излучение. Одновременно с радиацией (радиоактивная пыль, "горячие" частицы) воздействие на слизистую оболочку дыхательных путей табачного, коптильного дыма, выхлопных газов автотранспорта, содержащих канцерогенные и коканцерогенные продукты, способно усилить ингибирование ритмической активности мерцательного эпителия, образование сшивок и других труднорепарируемых повреждений ДНК клеток эпителия и, в перспективе, способно увеличить выход опухолей (бронхогенного рака). Таким образом, "чернобыльский синдром" не следует рассматривать как исключительно радиационный феномен; определенный вклад в него внесли также и факторы нерадиационной природы. 4.2.2. Изменения показателей функции эндокринной системы Острый, подострый и хронический лучевой синдромы, характерные для радиационного поражения различных контингентов риска Чернобыльской катастрофы, характеризуются как общими закономерностями, так и специфическими особенностями изменений со стороны эндокринной системы. К числу наиболее общих проявлений этих радиационных синдромов следует отнести мобилизацию нейрогуморальных систем поддержания гомеостаза, прежде всего симпатоадре-номедуллярной (САМС) и гипоталамо-гипофизо-кортикоадреналовой систем (ГГКАС). Сигналом для запуска этой реакции организма на лучевой стимул, в условиях отсутствия специфической рецепции ионизирующей радиации, являются первичные продукты лучевого воздействия на ткани - свободные радикалы и перекиси. При воздействии Других экстремальных агентов аналогичная реакция (стресс-реакция) развивается в результате дистантной рецепции и возбуждения высших гипоталамических центров САМС и ГГКАС (стресс-реализующих систем) [13, 14, 15]. Увеличение содержания в циркулирующей крови продукции катехоламинов (КA) и глюкокортикоидов (ГК) - гуморальных продуктов деятельности этих систем - отмечено в многочисленных исследованиях по изучению действия низкоинтенсивного облучения и в клинических наблюдениях за участниками ликвидации аварии и жителями районов "жесткого контроля". При этом в экспериментах показано, что в случае радиационного воздействия (как и при других стрессовых ситуациях) наиболее быстро реагирует САМС как "аварийная" система: концентрация КА уже в первые секунды и минуты облучения возрастает. Однако продолжительность гиперкатехолемии в условиях острого облучения невелика. Активация ГГКАС развивается более медленно, но носит более продолжительный характер - 2 - 3 сут и более. Характерно, что в период разгара острой лучевой болезни наблюдается вторичная активация САМС и ГГКАС. Регуляторные функции обеих систем тесно переплетаются, а их гуморальные продукты обладают первичным антиоксидантным действием, что позволяет рассматривать их мобилизацию при радиационном воздействии как проявление отрицательной обратной связи. В условиях постоянного (периодического) низкоинтенсивного облучения функция САМС и ГГКАС, как правило, носит колебательный характер, с периодическими снижениями концентрации их гуморальных продуктов. Однако реактивность обеих систем, в особенности в ответ на более мощное лучевое воздействие, полностью сохраняется [41]. При сочетаниях низкоинтенсивного облучения с эмоционально-болевым стрессом, высокими физическими нагрузками, воздействием тяжелыми металлами и т.п. активация САМС и ГГКАС сохраняется более длительно и устойчиво [10]. Клинические наблюдения за "ликвидаторами" подтвердили факт длительной, на протяжении 4-5 лет, активации САМС и ГГКАС. В условиях воздействия спектра радионуклидов ядерного взрыва (Хиросима, Нагасаки) или ядерной аварии (Чернобыль) первостепенное значение в ранний период приобретает поражение щитовидной железы изотопами йода. Смоделировать в эксперименте развитие радиогенного рака щитовидной железы пока не удалось, однако клинические наблюдения 1992 - 1993 гг. свидетельствуют о возникновении у детей из районов "жесткого контроля" Беларуси и Украины рака щитовидной железы (чего в нерадиационных условиях не наблюдается). Функциональные изменения развиваются быстро, опережая развитие морфологических изменений. Показателями гиперфункции щитовидной железы в ранние сроки воздействия являются повышение уровня в крови тироксина (Т4) и трийодтиронина (Тз); активная резорбция фолликуллярного коллоида; повышение йодсвязывающей способности белка плазмы; гиперплазия железы. Гиперфункция и гиперплазия щитовидной железы при малых дозах радиойода выражены умеренно и сменяются состоянием столь же умеренно выраженной гипофункции и гипоплазией железы. На фоне повышения общего и свободного Т4 крови наблюдается снижение уровня тиреотропина (по механизму отрицательной обратной связи), а по мере снижения Т4 - нормализация содержания и даже, в части случаев, существенное повышение тиреотропина, характеризующего состояние "лабораторного гипотиреоза" у детей, получивших более высокие дозы радиойода [123]; отмечена также тенденция к росту уровня аутоантител к тиреоглобулпну. При поглощенной дозе радиойода 500 - 1000 сГр наблюдаются длительные фазовые изменения тиреоидного эпителия, обратимые дегенеративно-дистрофические изменеия тиреоцитов. При дозах порядка десятков сГр отмечались менее выраженные морфологические изменения без полного восстановления функции железы. Складывается реальная опасность развития аутоиммунного тиреоидита, умеренного фиброза щитовидной железы. Гиперплазия щитовидной железы в условиях постоянного проживания в районах повышенного радиационного риска сохраняется длительно; определенный вклад в ее поддержание, по-видимому, вносит зобная эндемичность этих территорий. Зависимость регистрируемых изменений от дозы радиации невелика. С течением времени изменения в щитовидной железе в части случаев прогрессируют в направлении развития диффузных пролиферативных изменений и узелковых образований [123]. На экспериментальной модели затравки животных нуклидами75Se и37S, при поглощенной дозе на все тело 1,0 Гр, на эндокринные железы - 0,5 Гр, отмечены стойкие изменения - сначала повышение, а после набора полной дозы - снижение уровня гормонов надпочечников и щитовидной железы. Одновременно наблюдалось угнетение у животных репродуктивной функции, а у самок - развитие гормонозависимых опухолей молочной железы, матки, а также щитовидной железы и гипофиза [38]. Эндокринная функция поджелудочной железы в условиях длительного низкоинтенсивного облучения умеренно угнетена на фоне гиперсекреции антагонистов инсулина - глюкокортикоидов. Однако возможности специфической активации секреции инсулина сохранены. Вилочковая железа как лимфоидный орган существенно страдает в условиях пролонгированного облучения. Эндокринная функция тимуса, реализуемая его эпителиальными клетками, более радиорезистентна, но также угнетается глюкокортикоидами и, в условиях низкодозового облучения, понижена. Поскольку тимус является центральным регулятором созревания Т-лимфоцитов и, в целом, иммунной системы, угнетение его эндокринной функции является одной из причин формирования вторичного радиационного иммунодефицита.
4.4.3. Частотный анализ морфогенетических изменений у растений, произрастающих в зоне повышенных уровней хронического облучения Поскольку в контрольной группе растений многие аномалии практически полностью отсутствовали, а на опытных полигонах частота отклонений от нормы в первые 2 года после аварии колебалась от 20 до 26 %, то с полным основанием выявленные тераты могут быть отнесены к индуцированным радиацией, так как при облучении растений на гамма-поле наблюдали образование аналогичных радиоморфозов. Дозовый эквивалент для таких случаев предположительно может соответствовать дозам более 15 Гр. Большинство морфологических изменений (морфозов) связано с изменениями в меристематических тканях, представляющих собой группу растительных клеток в стадии активного деления и роста. В упрощенной форме эта ткань состоит из двух типов клеток: с высокой степенью репродукции; с различной степенью дифференциации. Результаты изучения репродуктивной способности сосны при разных уровнях облучения показали, что в собранных семенах урожая 1986 - 1988 гг. такие признаки, как пустозерность, жизнеспособность, масса семян, энергия прорастания, колеблются в широком интервале в зависимости от дозы облучения [117]. Собранные семена сосны высевали в тепличных условиях для изучения морфологических признаков и их наследуемости в потомстве. Анализ однолетних сеянцев и двухлетних саженцев показал высокую изменчивость по широкому спектру морфологических признаков, большинство из которых наблюдали на взрослых деревьях. Полученные данные свидетельствуют о том, что по мере увеличения продолжительности пребывания многолетних растений в зоне с радиационной аномалией, следует ожидать селективного увеличения полиморфизма и физиологической нестабильности. В этом случае пролонгированное радиационное воздействие на меристемы точек роста может способствовать преобразованию генетического и физиологического гомеостаза, что, по-видимому, может привести к микроэволюционным сдвигам. 4.4.4. Восстановительные процессы у многолетних растений, произрастающих в зоне радиационной аномалии Хронический режим облучения, сложившийся на территории 30-километровой зоны ЧАЭС после острого периода облучения весной 1986 г., затрудняет понимание природы лучевого поражения многолетних растительных организмов, так как радиочувствительность всех их компонентов претерпевает существенные изменения в течение годового цикла развития. Интерпретация результатов осложняется и тем обстоятельством, что в ходе облучения наряду с развитием эффектов поражения протекают восстановительные процессы. После острого периода облучения восстановительные реакции у многолетних древесных и кустарниковых пород наблюдали спустя 1-1,5 года. В качестве критериев восстановления использовали интенсивность таких процессов, как корне-, побего- и порослеобразования, физиологические реакции компенсаторного характера. Многолетние деревья сосны не способны к образованию поросли. У них мощным восстановительным потенциалом вегетативного характера обладает крона, имеющая большое количество спящих почек, защищенных круглогодично фотосинтезирующей хвоей [61,117]. Наблюдения в пострадиационный период (1986 - 1992) показали, что у сосны при дозе облучения свыше 14 Гр (сублетальная доза для сосны при остром весеннем облучении) восстановление происходило за счет дополнительного побегообразования из ранее спящих почек. Этот процесс начинался спустя год после облучения, т.е. в 1987 г., и выражался в появлении в верхней части кроны единичных побегов, представляющих собой пучки очень длинной хвои, сидящей на коротком толстом ауксибласте. Роль этих побегов заключается в обеспечении минимального уровня фотоассимиляции, необходимого для поддержания жизнедеятельности не пораженных облучением аттрагирующих органов и тканей дерева (корни, древесина ствола и ветки). Подтверждением этого является чрезвычайно высокая интенсивность фотосинтеза хвои таких побегов (в 3 - 4 раза выше контрольных), что свидетельствует о компенсаторном характере этого процесса. С каждым последующим годом количество таких побегов в кроне увеличивается, что указывает о возможности постепеного частичного восстановления исходных функций многолетнего растения. Начиная с вегетационного сезона 1988 г., в зоне сильного и среднего поражения при одних и тех же поглощенных дозах интенсивнее проходило восстановление деревьев более старшего возраста, а прирост побеговпоследнего года был выше в верхних мутовках, чем в нижних. Выживаемость сильно пораженных деревьев зависит от прироста побегов последнего года, и и если прирост составляет около 20% нормальной охвоенности при наблюдаемых темпах роста, то для восстановления таких деревьев потребуется 2-3 года. поэтомувозможно выживание отдельных деревьев, у которых количество побегов с зеленой хвоей в настоящее время составляет около 5-10%. Смена вершины дерева - пострадиационная реакция на гибель осевого верхушечного побега; его роль берет на себя один из боковых побегов мутовки, наиболее мощный. Это явление отмечали примерно у 30% сосен (Pinus sylvestris) и елей (Picea excelsa L.), облученных в пределах 11-15 Гр. О возможности восстановления многолетних растений, произростающих в 30-километровой зоне ЧАЭС, свидетельствуюти данные о динамике компенсаторной и репродуктивной генерации при различных дозовых нагрузках. В качестве критериев компенсаторной регенерации исползовали также такие показатели, как заживление ран на ветвях многолетних растений, спилы ветвей определенного диамета, величина вторичного прироста на многолетних растениях после удаления верхшечной почкиюВ качестве критерия протекания репродуктивной регенерации у растений, произрастающих на почвах с различным уровнем загрязнения радионуклидами, использовали такой показатель, как интенсивность ризогенеза у черенков смородины черной (Ribes nigrum), ивы остролисной (Salix acutifolia). Полученные результаты свидетельствуют о том, что при тех ууровнях дозового прессинга, которые наблюдаются на территории 30-километровой зоны ЧАЭС, регистрируется достоверное увеличение интенсивности всех типов регенерационных процессов у многлетних растений. 4.4.5. Мутагенное действие Под влиянием внешнего и внутреннего облучения радионуклидами, выброшенными при аварии, в растениях регистрировали мутации как в соматических, так ив половых клетках. Возникновение мутаций связывается, как правило, с прямым физическим разрушением либо функциональной инактивацией под влиянием продуктов радиолиза участков хромосом и их уникальных структур. О повреждении уникальных структур может свидетельствовать увеличение рахрывов в молекулах ДНК. Используя прием дополнительного гамма-облучения растений, уже получивших определенную дозу радиации, с помощью метода электрофореза однонитевой ДНК и щелочного расплетания, обнаружили в обследованных в 1987г. популяциях травянистых растений, произростающих в окрестностях ЧАЭС, в частости, у радиочувствительного бобового растения горшка мышиного (Vicia cracca L.) повышенный выход однонитевых разрывов ДНК [136]. Формирование генетических изменений и их фиксация в последующих поколениях во многом зависят от функционирования систем репарации ДНК. При этом у растений данные системы играют особо важную роль в пыльце, что обусловлено ее особой функцией и гаплоидностью ядер. Эффективность функционирования систем репарации ДНК должна обеспечить стабильность генома, который может легко повреждаться вследствие слабой защищенности созревшего пыльцевого зерна. поэтому ослабление функционирования систем репарации является свидетельством усиления генетического действия излучения. При излучении спонтанного обновления ДНК и индуцированного острым гамма-облучением внепланового синтеза ДНК, отражающего ее репарацию, было установлено, что формирование пыльцы березы бородавчатой (Betula pendula L.) при высоких концентрациях радионуклидов в почве (7,2·1013 по сравнению с 5,6·109 Бк/км2 в конроле) приводит к подавлению системы темновой репарации ДНК в зрелой хранящейся пыльце [22]. При сохранении у такой пыльцы способности к оплодотворению можно ожидать у растений накопление генетических повреждений. Многими исследователями отмечено увеличение аберраций хромосом в образовательных тканях растений, произростающих в зоне аварии ЧАЭС. Так, обнаружено [40] увеличение в несколько раз числа клеток с аберрациями хромосом в корневых мерисистемах проростков ослинника двухлетнего (Oenothera biennis L.), выращенных в лабораторных условиях из семян растений, произрастающих при уровнях гамма-фона в 1986г. от 0,05 до 60мР/ч. При этом показана характерная прямо пропорциональная зависимость выхода мутаций от мощности дозы и обнаружены множественные аберрации хромосом, которые могут быть следствием действия плотноионизирущего альфа-излучения. При проведении исследований [47] на различных по уровню загрязнения радиоактивными выпадениями участках (от 120 до 800 МБк/м2), наблюдали линейную или близкую к ней зависимость между поглощенной в течение 26.04 - 5.06 (сбор семян) 1986г. суммарной бета- и гамма-дозой растениями озимой посевной ржи (Secale cereale L.) и выходом клеток с аберрациями хромосом в корневых меристемах проростков М2-поколениями (рис. I.4.6). Этот же тест был использован [122] для оценки влияния радиоактивного загрязнения на семена 7 видов растений, собранных в зоне, прилегающей к ЧАЭС. Мощность дозы составила 2 - 3 мР/ч. При этом у отдельных различающихся по радиочувствительности видов было обнаружено от 1 до 8% аберраций хромосом, причем наблюдалось повышение частоты аберраций с увеличением мощности дозы. Но линейной зависимости не установлено, что, по-видимому, обусловлено неравномерным распределением радионуклидов по поверхности почвы или влиянием различных загрязнителей химической природы. Интересно, что на второй год после аварии частота аберраций хромосом у некоторых видов возросла в 1,5 - 3 раза несмотря на 2 - 3-кратное снижение радиационного фона. Этот факт имеет исключительно важное значение с точки зрения гентической опасности не только внешнего, но и внутреннего облучения за счет радионуклидов, поступающих в растения из почвы. При выращивании высокорадиочувствительных конских бобов (Vicia faba L.) сорта Русские черные [36] в 30-километровой зоне при уровнях радиоактивного загрязнения пахотного горизонта почвы по 90Sr около 2·1012 Бк/км2 и по137Cs около 4·1012 Бк/км2 (поглощенная доза для корневой системы за 3 мес выращивания растений сотавляла 0,15 - 0,4 Гр) регистрировали увеличение в 2 - 3 раза количества клеток с аберрациями хромосом.
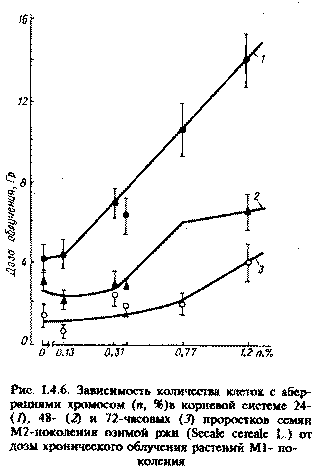
Рис. 1.4.6. Зависимость количества клеток с аберрациями хромосом (n, %) в корневой системе 24-(1), 48- (2) и 72-часовых (3) проростков семян М2-поколения озимой ржи (Secale cereale L.) от дозы хронического облучения растений М1-поколения
Кроме того, было установлено [126] увеличение количества клеток с аберрациями хромосом и хроматид в корневой меристеме проростков скерды кровельной (Crepis tectorum L.), выращенных из семян растений, собранных на территории зоны ЧАЭС при условиях гамма-фона 5-10 мР/ч в первый и 0,02 - 20 мР/ч во второй годы после аварии. В проростках первого года максимальная частота клеток с аберрациями достигала 10,2 - 15,3 %, причем нередко наблюдались клетки со множественными аберрациями. Во второй год на участках с максимальным уровнем фона ьыло обнаружено 1,4 - 2,2% клеток с аберрациями хромосом. При минимальных мощностях облучения частота клеток с аберрациями составила всего 0,3 - 0,5%; эти значения соответствуют спонтанному уровню. Кроме того, отмечали также появление растений с измененным кариотипом, что свидетельствует об активных микроэволюционных процессах в хронически облучаемых популяциях. Имеются данные [88], свидетельствующие о повышении количества стерильной пыльцы у растений, растущих на территориях, загрязненных радионуклидами. Так, исследуя две популяции фиалки утренней (Viola matutina klok), произрастающей на двух участках 30-километровой зоны ЧАЭС, различающихся на 3 порядка по величинам поглощенных доз, выявили в 1987 и 1988 гг. увеличение в 1,5 - 2 раза количества стерильных пыльцевых зерен. При уровне мощности дозы на высоте 2 м 4 - 5 Гр/год наблюдали частичную женскую стерильность сосны (Pinus sylvestris L.), выражавшуюся в снижении гамето-фитной выживаемости семяпочек, опыленных в 1986 г. и уменьшении эмбриональной выживаемости семяпочек, опыленных в 1985 г. Безусловно, стерильность - неспособность организма образовывать гаметы или достаточное их количество, не приводит к генетическому повреждению, а лишь снижает число потомков. Но при действии ионизирующей радиации стерильность, как правило, является следствием генных или хромосомных мутаций, обусловливающих нарушение мейоза - основного звена гаметогенеза. Прямым свидетельством генетического действия ионизирующих излучений на пыльцу является непосредственная регистрация в них мутаций. У растений, выращиваемых на почвах 30-километровой зоны ЧАЭС с общей бета-активностью 2,6·103- 6,3·104Бк/кг, в пыльце ячменя (Hordeum vulgare L.) отмечали увеличение частоты waxy-изменений в 2 - 3 раза [34]. Таблица І.4.7 Количество хлорофилльных мутаций типа альбина у ржи (Secale cereale L.) и ячменя (Hardeum vulgare L.), %
Культура |
Контроль |
1986 г. |
1987 г. |
1988 г. |
1989 г. |
Рожь Киевская 80 |
0,01 |
0,14 |
0,40 |
0,91 |
0,71 |
Рожь Харьковская 03 |
0,02 |
0,80 |
0,99 |
1,20 |
1,14 |
Ячмень |
0,40 |
0,90 |
0,74 |
0,80 |
0,91 |
Примечание. Семена получены в 30-километровой зоне ЧАЭС . В первые месяцы после аварии при выращивании растений ржи (Secale cereale L.) и ячменя (Hardeum sativum jessen) в условиях закрытого грунта на почвах, привезенных из 30-километровой зоны, было обнаружено увеличение частоты встречаемости различных типов хлорофилльных мутаций. Преобладали нежизнеспособные мутации типа альбина. В последующие годы семена, собранные в зоне, высевали вновь. У ржи и ячменя, выращиваемых в 30-километровой зоне на почве с суммарной гамма-активностью 1·105Бк/кг, регистрировали многократное превышение спонтанного уровня выхода хлорофилльных мутаций (табл. 1.4.7). При этом было отмечено, что в каждом последующем поколении мутации не элиминировались, поскольку высеянные семена подвергались последующему прессингу радионуклидов. Выход морфологических аномалий у 4 сортов озимой пшеницы (Triticum vulgare L.) при их выращивании в течение 3 лет на почве с суммарной активностью гамма-излучения 5,2·104- 1,8·105Бк/кг в 1986 - 1987 гг. превысил 40 %, но в 1988 г. уменьшился. Спектр выявленных аномалий довольно широк: в первый год после аварии наиболее частым (до 49 %) было образование в колосьях стерильных зон - череззерница; в 1987 г. количество аномалий этого типа достигало 30 %, а в следующем поколении снижалось до 1,9 %. Часто встречались колосья с дополнительными колосками ("грыжи") и укороченные колосья. К распространенным изменениям структуры колоса следует также отнести аномалии типа скверхед. Встречались также растения с повышенной остистостью, неравномерной остистостью, колос "елка", изменения окраски стебля и другие. У разных сортов выход морфологических аномалий проявляется по-разному, что может быть связано, с одной стороны, с различной их радиочувствительностью, а с другой - с неодинаковой способностью к накоплению радионуклидов. Безусловно, отмечаемые морфологические аномалии, отклонения от нормы, как правило, имеют место лишь в поколениях облучаемых организмов и обычно не закрепляются в потомстве. Тем не менее их стойкое проявление во все последующие годы, вплоть до 1994 г., позволяет предполагать, что многие из этих изменений носят генетический характер и свидетельствуют о том, что под влиянием ионизирующих излучений радионуклидов, выброшенных при аварии на ЧАЭС, в популяциях растений идет мутационный процесс. 4.4.6. Действие на растения инкорпорированных радионуклидов Радиочувствительность растений к облучению, создаваемому в результате накопления в тканях радионуклидов, изучена меньше, чем радиочувствительность к облучению от внешних источников. Главным образом это обусловлено тем, что большинство видов растений обладает более высокой по сравнению с млекопитающими радиоустойчивостью, и даже при накоплении радионуклидов в значительных количествах действие излучений у них проявляется гораздо в меньшей мере. Растениям обычно отводится роль переносчиков радионуклидов по пищевой цепочке "почва - растение - животное - человек" или "почва - растение - человек". Тем не менее при поступлении и накоплении радионуклидов в относительно радиочувствительных видах растений в зависимости от физико-химических характеристик радиоактивных веществ, их количества, мест локализации могут наблюдаться те же радиобиологические эффекты, что и при эквивалентных поглощенных дозах внешнего облучения: радиационная стимуляция, морфологические изменения, лучевая болезнь, ускорение старения и сокращение продолжительности жизни, гибель, генетическое действие. Однако поступившие внутрь растений радиоактивные вещества могут обладать повышенной по сравнению с внешним облучением опасностью, вызванной рядом причин. Первая и основная из них - способность некоторых радиоактивных веществ избирательно накапливаться в отдельных тканях и органах, что приводит к относительно высоким локальным уровням облучения. Вторая причина - увеличение опасности действия альфа-и бета-излучателей, которые мало влияют на растения в условиях внешнего облучения, но могут стать необычайно сильным источником ионизирующей радиации при поступлении внутрь клеток и клеточных структур. Третьей причиной являются, как правило, длительные сроки облучения. Ввиду специфики метаболизма растений радионуклиды, кумулируясь в тканях, практически не выводятся из них в отличие от животных. При огромных периодах полураспада90Sr,137Cs,239Pu даже у многолетних растений эти радионуклиды будут обусловливать облучение в течение всего онтогенеза. И если в первый год аварии облучение растений было обусловлено в значительной степени за счет внешнего облучения, то в последующие годы с начавшейся миграцией радионуклидов по трофической цепочке, оно почти исключительно обусловливается внутренним облучением. Но если у одно-двухлетних растений за сравнительно короткий период вегетации не всегда успевают сформироваться достаточно высокие дозы облучения, то у многолетних растений в течение десятков лет могут сложиться сравнительно высокие уровни облучения. Более того, подавляющее количество видов многолетних растений, как правило, чувствительнее к действию радиации, чем одно-двухлетние травянистые виды. В зоне Чернобыльской катастрофы в кроне древесных растений первоначально было задержано от 60 до 90 % выпавших на лес радионуклидов. После выпадения радиоактивных осадков на лесную растительность начинается их вертикальная миграция под влиянием сил гравитации, атмосферных осадков, движения воздуха, с листовым опалом. в результате чего радионуклиды перемещаются в нижние слои крон, под полог леса. Скорость вертикальной миграции зависит от физико-химических характеристик радиоактивных выпадений, типа и возраста древостоя, метрологических условий, времени года. Спустя некоторый период, который в хвойных лесах может измеряться несколькими годами, основная масса радиоактивных веществ переходит в лесную подстилку и в верхний слой почвы. Именно из него через 4 - 5 лет в лиственном лесу и через 8 - 10 лет в хвойном (что обусловлено особенностями в скорости вертикальной миграции радионуклидов и более быстрой минерализацией листьев по сравнению с хвоей) начинается активное поступление радионуклидов в древесные растения через корни. Если механизмы поступления и усвоения радионуклидов однодвухлетними травянистыми и многолетними древесными растениями в основном не различаются, то характер из накопления имеет принципиальные различия. Многолетние древесные способны к существенному накоплению радионуклидов во всех органах - листьях, ветках, коре, древесине. Многолетний замкнутый цикл радионуклидов по цепочке "листья - лесная подстилка - почва - корни - ствол - листья" и так далее может приводить к существенному радиоактивному загрязнению всех органов. У одного из самых радиочувствительных видов - сосны - это может приводить к самым различным типам нарушений. Повреждаться могут и другие виды. Ель, например, обладает более высокой радиоустойчивостью, но она является кальциефилом и способна накапливать -Sr в больших количествах, чем сосна и другие виды. Различные радионуклиды не только по-разному накапливаются отдельными видами растений, но и по-разному концентрируются в различных органах, приводя к различной степени облучения. На радиоавтографах целых растений, полученных при введении различных радионуклидов, как правило, бывает хорошо определена неравномерность их распределения по отдельным органам. Четко прослеживаются места концентрации, сосредоточенные, как правило, в тканях, которые обладают высокой метаболической активностью (меристема и генеративные органы) и наиболее высокой радиочувствительностью. Именно поэтому как при внешнем, так и при внутреннем облучении радиобиологические эффекты, в первую очередь, проявляются на этих группах клеток. В частности, в меристемах наблюдаются замедление или ускорение деления клеток, формирование различного рода морфологических изменений, замедление роста органов и растения в целом, гибель меристем и др. В год аварии в пяти областях Украины (Киевской, Черниговской, Житомирской, Черкасской и Винницкой), в том числе в различных местах 30-километровой зоны, были собраны образцы семян 9 сортов озимой пшеницы (Tviticum vulgare L.), растения которой в течение 3 мес - с фазы кущения (время аварии) до созревания - подвергались облучению радиоактивными выбросами. Максимальный уровень суммарной бета- и гамма-активности семян достигал 3,7·104Бк/кг. По общепринятым методикам экспериментального мутагенеза в последующих б поколениях изучались частоты и спектр мутационной изменчивости растений. У 18,4 % образцов выявили достоверное увеличение (более чем в 2,7 раза) частоты хромосомных аберраций в клетках меристемы первичного корешка прорастающих семян. В одном случае у проростков из семян растений, выросших в 16 км от станции, наблюдали увеличение в 32 раза количества аберраций. Регистрировали увеличение в 2,7 - 8,7 раза частоты видимых мутаций. Их спектр охватывал 27 типов мутаций по признакам структуры стебля, листа и колоса, физиологическим признакам. Спецификой спектра были сравнительно частые мутации по генам карликовости, а также редко встречающиеся в обычных условиях мутации типа "ощипанный колос" и "разветвленный колос". Мутации полимерных генов по элементам структуры урожая в последующих поколениях привели к изменению продуктивности мутантов и в итоге к потере типичности сортов. При выращивании проростков гороха (Pisum sativum L.) и кукурузы (Zea mays L.) на водных вытяжках почв из 10-километровой зоны ЧАЭС, имеющих суммарную бета-активность 5·10-9- 5·10-8Ки/л, наблюдали [46] заметное снижение относительной скорости роста растений. Радиационную стимуляцию, выражавшуюся в ускорении прорастания семян, роста растений, прохождении фаз развития, повышении фотохимической активности хлоропластов, увеличении содержания хлорофилла, наблюдали у желтого люпина (Lupinus luteus L.), выращенного из семян, которые были собраны в 30-километровой зоне ЧАЭС при уровне загрязнения по137Cs Ки/км2. Такие семена содержали137Cs 2,31·103и90Sr 1,65·102Бк/кг [115]. Наибольшую чувствительность к радиоактивному загрязнению проявили сосновые леса по причине высокой радиочувтвительности этого вида растений, значительной задерживающей способности хвои и, как следствие, высокого вклада бета-излучения в дозовую нагрузку [49]. Общая поглощенная доза в хвое и почках примерно в 10 раз больше дозы внешнего гамма-облучения. Именно поэтому на участках с высоким уровнем загрязнения в активно растущих меристемах обнаруживали наиболее сильные повреждения, приводящие к их массовой гибели, пролифирации боковых почек с неупорядоченной ориентацией, образованию укороченных побегов с мелкой или гигантской хвоей, искривлению иголок хвои, нарушению ориентации молодых вегетативных побегов. К концу лета 1986 г. проявились летальные эффекты облучения сосны. В течение последующего осенне-зимнего периода было выявлено, что общая площадь погибшего лесного массива, примыкающего с запада к промплощадке АЭС, достигает 500 га. Спустя 4 года после аварии наблюдалось снижение в 5 - 10 раз продуктивности семян у аборигенной популяции подорожника ланце-толистного (Plantago lanceolata L.), произрастающей в местах с уровнем гамма-фона от 2,8 до 1334 пА/кг [120]. По-видимому, данный тип радиационного поражения растений можно связать с угнетающим действием многолетнего хронического облучения на популяцию этого радиочувствительного представителя травяного покрова, не выделяя вклад внешнего и внутреннего облучения. Кроме того, без сомнения, можно утверждать, что на 4-й год после аварии, когда внешнее облучение по сравнению с первоначальным поставарийным периодом уменьшилось во много раз, фактор внутреннего облучения в радиационном повреждении растений играет определяющее значение. Особенно это касается травянистых растений с мочковатой корневой системой, формирующих основную массу корней в верхнем, наиболее загрязненном радионуклидами слое почвы. Тем не менее следует признать, что главная опасность накопления растениями радионуклидов состоит не в их традиционной угрозе для растений, а в том, что они являются главным звеном миграционной цепочки на пути передачи радионуклидов более радиочувствительным видам живых организмов - млекопитающим и, в первую очередь, человеку. При оценке функциональной активности лимфоцитов крови и периферических лимфоидных органов выявлены: нарушения реакции на поликлональный Т-клеточный митоген [98] при одновременной активации функции К-клеток (антителозависимая цитотоксичность); подавление кооперативных Т-клеточных реакций - трансплантационного иммунитета, гиперчувствительности замедленного типа [97]. Достаточно типичны волнообразные изменения способности лимфоцитов к контактному взаимодействию с аллогенными тканевыми базофилами. Такое взаимодействие, согласно современным представлениям, определяется степенью дифференцировки лимфоидных клеток и опосредует их участие в регуляции аллергических реакций немедленного и замедленного типов, а также регуляции гуморального иммунитета [37]. К биологическим эффектам постоянного облучения относится и прогрессирующее со временем снижение "спонтанной" антигеннеспеци-фической Т-супрессии [97]. Показатели, характеризующие В-систему иммунитета, более стабильны. При обследовании нескольких поколений линейных мышей, постоянно содержащихся в Чернобыле, не выявлено существенных изменений содержания и пролиферативной активности В-лимфоцитов в периферических лимфатических узлах. Ответ на поликлональный В-митоген (декстрансульфат) и уровни сывороточных иммуноглобулинов, а также специфический гуморальный иммунный ответ на заражение вирусом гриппа у этих животных также значительно не изменялись. О сохранении способности к активному антителообразованию свидетельствует и выраженная стимуляция аллергической реакции немедленного типа - достоверное повышение содержания Jg E-антител в органах дыхания мышей в ответ на иммунизацию аллергеном амброзии [99]. В отдаленные сроки радиационного воздействия выявлено и возрастание уровня аутоантител к собственным эритроцитам и эпителиальному ретикулуму тимуса [20]. Эти данные указывают не только на большую сохранность гуморального иммунитета по сравнению с клеточным, но и на срыв толерантности к собственным тканям. Последнее свидетельствует о высокой вероятности развития ау-тоиммунных поражений облученного организма. Поздние реакции на постоянное действие факторов радиационной аварии другой группы клеток иммунной системы - моноцитов (макрофагов) - изучены в меньшей степени. Известно, что в костном мозгу страдают клетки моноцитарного ряда кроветворения [8]. Выявлены усиление поглотительной активности макрофагов брюшной полости и активация ферментов "дыхательного взрыва" фагоцитирующих клеток до критических уровней. Наряду с этим обнаруживается четкая тенденция к истощению функционального резерва клеток [101,106]. В детальном изучении на экспериментальных моделях нуждается продукция моноцитами (макрофагами) цитокинов, играющих важную роль в развитии воспалительной реакции, в процессах пролиферации и дифференцировки клеток иммунной системы, в противоопухолевой резистентности, межсистемных взаимодействиях, развитии компенсаторных процессов. Важность фундаментальных исследований такого рода, помимо теоретических посылок, обусловлена и тем, что к настоящему времени показано, что у ликвидаторов к числу отдаленных эффектов относится изменение концентрации сывороточных цитокинов этой группы (в первую очередь ИЛ-1b ) [19]. Получены предварительные результаты об изменении уровней регуляторных цитокинов и у постоянно облучаемых экспериментальных животных [21, 107]. При изучении показателей естественной резистентности выявлено снижение лизоцимной активности сыворотки крупного рогатого скота [18, 53]. При интегральной оценке иммунологической реактивности животных, постоянно подвергающихся действию вредных факторов аварии на ЧАЭС, установлено развитие иммунодефицитов, проявлениями которых являются: снижение антимикробной устойчивости кожи у крупного рогатого скота [18] и диких мышевидных грызунов [60]; повышение чувствительности к экспериментальным вирусным инфекциям и прививке экспериментальных штаммов опухолевых клеток у лабораторных мышей [96,98]. Важно отметить, что развитие иммунодефицитных состояний наблюдается на протяжении всего срока проведения исследований (1986- 1993), т.е. иммунодепрессивное действие факторов аварии на ЧАЭС - долговременный биологический эффект. Существенное теоретическое и практическое значение имеет тот факт, что, хотя иммунологическая недостаточность развивается вне зависимости от того, в каком периоде индивидуального развития началось радиационное воздействие, степень нарушений иммунного статуса возрастает, а сроки их появления сокращаются тем значительнее, чем моложе организм. Наибольшие изменения иммунной реактивности обнаружены у потомков облученных родителей, в свою очередь начинающих подвергаться постоянному облучению уже с эмбрионального периода. Анализ динамики изменений в иммунной системе показывает, что на ранних этапах (первые месяцы) постоянного облучения в малых дозах наряду с повреждающим действием факторов радиационной аварии наблюдаются признаки функциональной напряженности, компенсаторной и репаративной реакций. За счет последних отдельные иммунологические показатели могут превышать контрольный уровень, создавая, на первый взгляд, впечатление активации иммунной системы [72]. Однако полноценная адаптация системы иммунитета, видимо, отсутствует, ее компенсаторно-репаративные возможности истощаются и, по мере увеличения возраста животных либо числа поколений, выявляются нарушения преимущественно деструктивного характера, а также значительные нарушения иммунного гомеостаза. Изучение возрастной динамики характеристик иммунокомпетентных органов и иммунных реакций у животных, содержащихся на экспериментальной базе в Чернобыле, позволило высказать предположение об ускорении темпов старения иммунной системы при постоянном внешнем и внутреннем облучении малыми дозами радиации низкой интенсивности [98]. Ускоренное развитие возрастной супрессии прямо показано в модельных экспериментах на линейных мышах, облучаемых дважды в неделю дозой 0,07 сГр в течение месяцев [35]. Следует отметить, что признаки "радиационного старения" тимуса обнаружены при обследовании облученных - "ликвидаторов" и пациентов с проявлением отдаленных последствий острой лучевой болезни [20] Совокупность результатов, полученных разными исследованиями, показывает, что структура и выраженность иммунодефицитов может очевидно, варьировать в зависимости от дозы облучения, спектра и распределения радионуклидов в организме, генетических особенностей (данные, полученные на инбредных мышах разных линий) и исходного физиологического состояния последнего. Однако в большинстве исследований наблюдается преимущественная связь отдаленных эффектов факторов Чернобыльской катастрофы с повреждением тимусзависимого (Т-) звена системы иммунитета. Важно, что подобные закономерности выявлены и в исследованиях, посвященных иммунному статусу взрослого и детского населения, пострадавшего в результате аварии на ЧАЭС, в том числе лиц, принимавших участие в ликвидации последствий аварии [5]. На основании накопленной в настоящее время информации можно предполагать, что патогенез постчернобыльских иммунодефицитов имеет, вероятно, сложный комплексный характер и включает ряд компонентов: прямое и непрямое повреждающее действие ионизирующих излучений на клетки иммунной системы, включая стромальные элементы и вспомогательные клетки; нарушение процессов созревания и дифференцировки иммунокомпетентных клеток (преимущественно Т-лимфоцитов); нарушение функций центрального органа иммунитета - тимуса; развитие аутосенсибилизации (в том числе и к клеткам эпителиального ретикулума тимуса); глубокую дискоординацию иммунорегуляторных процессов и взаимодействий в рамках иммунной системы; изменение гормональной регуляции иммунной реактивности, связанное с нарушением в эндокринной системе. Приведенный перечень может быть далеко не исчерпывающим; не вполне ясно также, какие из перечисленных феноменов первичны, а какие вторичны. Однако более детальное обсуждение механизмов развития иммунодефицитов при постоянном воздействии на организм млекопитающих факторов аварии на ЧАЭС преждевременно. 4.2.5. Влияние на кроветворную систему Костномозговое кроветворение (КМК) относится к тем системам, которые благодаря наличию стволового пула и относительной автономии пролиферации сравнительно быстро реагируют на воздействия ионизирующей радиации. При исследовании системы крови у животных после аварии на ЧАЭС обнаружен ряд особенностей реакции КМК при постоянном пребывании их в условиях внешнего и внутреннего облучения в полях низкой интенсивности и различного качества. Наблюдения велись с июня 1986 г. на диких мигрирующих животных, на экспериментальных животных в условиях вивариев Чернобыля и Киева. Некоторые ветеринарные хозяйства организовали наблюдения над крупным рогатым скотом и овцами. Наблюдения продолжаются по настоящее время, особенно в местах с плотным загрязнением по цезию. Выявленные гематологические эффекты, как правило, превышают ожидаемые для дозовых нагрузок, исходящих из экстраполяции данных, описанных при воздействий на организм больших доз. Некоторые различия в степени выраженности изменений КМК обусловлены особенностями проведения опытов (временем, прошедшим после аварии, удаленностью от реактора места проведения экспериментов, продолжительностью самого эксперимента). С октября 1986 г. систематически проводятся исследования КМК на белых беспородных крысах, завозимых в половозрелом трехмесячном возрасте в Чернобыль и наблюдаемых до момента их естественной гибели. Дозовые нагрузки не превышали 3 сГр за жизнь животного. К настоящему времени систематизирован материал о состоянии КМК у крыс трех серий экспериментов: 1986 - 1989, 1989 - 1991, 1991 - 1993 гг. Наиболее выраженные количественные изменения клеточного состава костного мозга и периферической крови зарегистрированы в первой серии опытов. У животных чернобыльской группы отмечались: умеренно выраженная гипохромная анемия; прогрессирующая с третьего месяца пребывания в зоне лейкопения, преимущественно за счет лимфоцитарной фракции, достигающая к моменту гибели 30 - 40 % от исходного уровня; снижение числа миелокариоцитов на 50 - 60 %. Однако наиболее существенным было наличие гранулоцитопении с очень высоким содержанием эозинофилов. Изменения в миелограмме наблюдались по гипопластическому типу (уменьшение молодых дифференцирующихся элементов с возрастанием доли зрелых гранулоцитов, ретикулярных и плазматических клеток). При этом в киевской группе животных гематологические изменения носили однонаправленный характер, но развивались значительно более медленно. В последующих сериях экспериментов ни в чернобыльской, ни в киевской группах на протяжении жизни животных не наблюдалось значительного снижения клеточности костного мозга и лейкоцитов периферической крови. Обращает на себя внимание тот факт, что в каждой последующей серии снижается исходный уровень лейкоцитов. Это свидетельствует о тенденции к постоянному уменьшению плацдарма КМК под влиянием ухудшившейся радиоэкологической обстановки. К стабильным явлениям, наблюдаемым в каждой серии опытов, относятся относительная и абсолютная эозинофилия и наличие патологических клеток, обычных для радиационных поражений (гигантские гиперсегментированные нейтрофилы, клетки с фрагментозом ядер, косматой структурой хроматина, включением ядерного вещества в цитоплазму, дву- и многоядерные лимфоциты, полиморфно-ядерные лимфоциты, мононуклеары и др.). Эозинофилию и атипичные клетки регистрируют практически все исследователи, изучающие систему КМК у животных. Наблюдаются они и у лиц, принимавших участие в ликвидации последствий аварии и у проживающих на загрязненных радионуклидами территориях. Этот феномен требует тщательного изучения, являясь показателем наличия в организме аутоиммунных реакций и развития эндогенной интоксикации. К особенностям реакции КМК относятся также выявленные изменения пролиферативной активности костного мозга. У животных всех серий экспериментов после 3 - 6-месячного пребывания в Чернобыле отмечалось первичное значительное увеличение митотической активности, в ряде случаев сопровождаемое нарастанием клеточности костного мозга, с последующим выраженным снижением числа митозов. Механизм этого процесса остается невыясненным. Аналогичные результаты получены при изучении системы КМК у Диких грызунов, отлавливаемых в зоне аварии, получавших внешнее гамма-облучение от 5,16·10-9до 5,16·10-5Кл/кг. В реакции крови отмечено две фазы: усиление компенсаторных процессов (активация эритро- и миелопоэза) и декомпенсация (на фоне лейко- и эритропений происходит обильный выход в периферическое русло бластных форм и атипичных клеток). В работах [53 - 55] приведены изменения гематологических показателей крупного рогатого скота, в течение 2 мес пребывавшего на расстоянии 9 - 12 кмј от ЧАЭС. У животных наблюдалась эритропения, снижение концентрации гемоглобина, уменьшение процентного содержания нейгрофилов и моноцитов, эозинофилия, качественные изменения клеток белой крови. У крупного рогатого скота, свободно обитавшего в радиусе 3 - 6 км от аварийного блока до октября 1987 г., в лейкограмме выявлена выраженная эозинофилия, сдвиг в формуле влево, лимфопения, наличие ретикулярных, недифференцированных клеток, фигур митозов, распадающихся форм; отмечалась гаперхромная анемия [б]. Количественные и качественные изменения показателей КМК отмечались и у лабораторных животных, кратковременно экспонированных в реперных точках 30-километровой зоны ЧАЭС. Например, отмечено, что у крыс после их экспонирования в течение 30 дней в с. Янов (доза 0,6 Гр) наблюдаются уменьшение лейкоцитов с 8,8 до 3,0·10-9 кл./л и тенденция к снижению клеточности костного мозга, содержание эритроцитов не изменилось [48]. У мышей, экспонируемых в этой же точке, в периферической крови обнаружили уменьшение числа лимфоцитов и лейкоцитов [81]. Исследования по изучению стволового пула КМК единичны. По данным ряда авторов [82,92], у мышей, экспонируемых на реперных точках в 30-километровой зоне ЧАЭС в 1991 и 1992 гг. (суммарные дозы 24 и 120 мГр), наблюдается изменение стволовых потенций костного мозга. На основании экспериментов с дополнительным облучением животных в дозе 1,5 Гр установлено, что пребывание в зоне повышает радиочувствительность колониеобразующих единиц селезенки, т.е. отсутствует адаптивный ответ при остром облучении после экспозиции мышей в зоне аварии. Основные механизмы поражения КМК, по-видимому, следующие: 1) внешний гамма-фон всей совокупности среды обитания; 2) соприкосновение циркулирующей крови со всем спектром нуклидов, вдыхаемых легкими. При этом все газообразные и аэрозольные радионуклиды могут проникать в кровь через альвеолярные мембраны и мембраны сосудистого эндотелия традиционным путем транскапиллярного обмена. Иными словами, в сосудистом русле, в том числе и в межтрабекулярных полостях и синусах костного мозга, где располагаются кроветворные клетки, может создаваться определенная концентрация не тропных по отношению к костной или кроветворной ткани радионуклидов, постоянно воздействующих на организм, как бы Облучая "извне" кровь и кроветворные органы. Тот же механизм попадания в кровь и соединений, содержащих радионуклиды, заглатываемые с пищей: через ворсинки кишечника, имеющие так назывемые окончатые капилляры, они попадают в систему воротной вены печени, откуда разносятся по всему организму, и, в свою очередь, являются одной из слагаемых лучевого воздействия на костномозговое кроветворение и периферическую кровь; 3) постоянное воздействие излучения на кровь и кроветворные органы имеет и геометрически "обратный" путь воздействия - либо непосредственно путем пробега частиц из фиксированных в тканях радиоизотопов, либо из радионуклидов, находящихся в тканях и клетках в растворимых соединениях, и проникающих обратно в кровь всеми классическими путями транскапиллярного обмена. Иными словами, осуществляется постоянный обмен энергии ионизирующего излучения между циркулирующей кровью и тканями организма с поддержанием относительного постоянства концентрации суммарных радионуклидов в капиллярах и синусоидах костного мозга, непосредственно кровоснабжающих межтрабекулярные полости, в которых содержатся стволовые и дифференцирующиеся элементы КМК; 4) действие остеотропных радионуклидов, таких, как90Sr и239Pu накапливающихся в эндостальной поверхности; кости, т.е. непосредственно прилегающей к трабекулярным поверхностям или поверхностям костномозговых каналов, при том, что все стволовые и бластные клетки костномозговой ткани располагаются строго по периферии. Кроме радиобиологических эффектов, развивающихся по канонам взаимодействия ионизирующего излучения от инкорпорированного .источника с веществом живой ткани, 239Ри, обладая альфа-излучением, имеющим энергию, превышающую 5 мэВ с пробегом частиц до 250 мкм в жидкой фазе, будет обладать и выраженным прямым повреждающим действием на все клетки КМК с преобладанием повреждения стволового и коммитированных пулов, но могущим повредить клетки любой степени дифференцировки, в том числе и зрелые, а ,также стромальные клетки кроветворного микроокружения; 5) и, наконец, соприкосновение всех классов дифференцирующихся в. костном мозге клеток стромального микроокружения, а также клеток периферической крови с "горячими" частицами, создающими .колоссальное энергетическое поле вокруг себя и обладающими очень большим прямым повреждающим действием, прямо зависящим от суммарной энергии ионизирующего излучения "горячей" частицы. Кроме перечисленных механизмов прямого повреждения клеток КМК инкорпорированными радионуклидами, в патогенезе костномозгового синдрома существенную роль играет развивающаяся эндогенная интоксикация [85]. 4.2.6. Отдаленные последствия, заболеваемость и продолжительность жизни Биологические эффекты, причинно обусловленные действием радиогенного фактора на организм животных и человека, классическая радиобиология относит к стохастическим или нестохастическим. Стохастические реакции имеют определенную вероятность появления, преимущественно с линейно-квадратической зависимостью от дозы радиации, существенно зависят от качества излучения и проявляют беспороговую закономерность. В основе механизма реакций лежат повреждения генетического .материала, инициирующие такие специфические эффекты, как мутации или предраковые состояния. Эти эффекты связаны с повреждением одной или нескольких клеток. К стохастическим реакциям, как правило, относят генетические и канцерогенные эффекты. Нестохастические эффекты, как считают, имеют пороговую дозу, превышение которой усиливает тяжесть реакций; они тканеспецифичны. К этим эффектам относят как соматические, так и наследственные, Нестохастические реакции являются следствием повреждения определенного количества клеток. Тяжесть эффекта зависит от количества таких повреждений. При фракционировании дозы радиации с низкой линейной потерей энергии (ЛПЭ) величина эффектов уменьшается. Для излучений с высокой ЛПЭ этой зависимости не существует. Однако эта классификация чисто условная и относительная, поскольку многие нестохастические эффекты могут быть следствием стохастических; они взаимосвязаны и взаимообусловлены многообразием индивидуальных реакций, зависимостью от времени наблюдения и пр. Так, в более ранних сообщениях американских и японских исследователей при традиционных подходах к оценке рисков отдаленных последствий у людей, перенесших атомную бомбардировку, отмечено три класса эффектов: не зависимые от дозы радиации (ускорение старения, бесплодие, врожденные дефекты, увеличение смертности в первом поколении, остеосаркомы, хронические лимфолейкозы); предполагаемо зависимые от дозы (изменение иммунного статуса, некоторые злокачественные новообразования кишечника, пищевода, нервной системы, слюнных желез и лимфомы); с выраженной зависимостью от дозы излучений (катаракта, абберации хромосом, злокачественные опухоли, микроцефалия, дефекты умственного и физического развития у облученных внутриутробно). На основании этих наблюдений рассчитаны риски отдаленных последствий. Однако с учетом дополнительных данных, более тщательного и всестороннего их анализа Комитет до биологическим эффектам ионизирующей радиации Национальной академии наук США (БЭИР-5) пришел к выводу о существенной недооценке рисков многих заболеваний, в частности опухолевого генеза. Согласно другим данным, биологические эффекты малых доз радиации низкой интенсивности могут быть увеличены в 100 - 1000 раз по отношению к ожидаемым при экстраполяции их из области больших доз радиации. Отличительная особенность Чернобыльской катастрофы при сравнению с ранее имевшими место случаями переоблучения людей состоит в том, что огромные контингенты населения, включая его критические группы и потомков облученных родителей, вынуждены постоянно находиться в надфоновых радиационных полях. Эти поля формируются не только за счет внешней g-компоненты, но, что особенно важно, и радионуклидов, попадающих в организм ингаляционным путем и с пищей, инкорпорирующихся в органах и тканях. Особенности стохастических и нестохастических отдаленных последствий такого рода радиационного воздействия в настоящее время изучены недостаточно. В настоящей главе приведены результаты исследования некоторых из них, в частности, заболеваемости и смертности у лабораторных животных, в течение всей жизни (1986 - 1989) находившихся в реальных условиях радиоактивных выпадений после аварии на ЧАЭС. В качестве биологической нормы использованы результаты исследований, полученные до аварии на ЧАЭС (1980 - 1986) у животных того же вида и разводки того же вивария (доаварийная норма). Понимая некоторую относительность выбранного контроля к опытным группам животных, при планировании эксперимента исходили из практических трудностей создания условий, обеспечивающих адекватную биологическую норму. Кроме того, учитывалось , что в доаварийный период на больших выборках этих животных выполнены обширные исследования многих биологических показателей, включая половые, возрастные особенности, а также суточные и сезонные ритмы, не только в норме, но и при однократном воздействии на них радиацией разного вида и в разных дозах. Следует отметить, что значительная часть опытных животных погибла в относительно молодом возрасте (до 2 лет), что не могло не сказаться на общем выходе новообразований. До аварии основная часть неопухолевых животных погибала после 30 мес, и их общая гибель составила 41,7 %. Животные чернобыльской группы без опухолей ,начали погибать с 14 мес, а в киевской группе - с 18 мес. Для этих групп характерно быстрое нарастание во времени процента погибших животных. Так, например, к 30-му месяцу жизни у интактных животных (до аварии) погибло 8 % животных без опухолей, а в киевской и чернобыльской группах соответственно 57 и 52,5 %. В киевской и чернобыльской группах животных значительно отличаются интервалы времени, в течение которых отмечена максимальная скорость гибели (20 - 23 и 19 мес соответственно). Следовательно, увеличение радиационной нагрузки у животных сдвигает пик их гибели на более ранний возраст. Общая гибель животных, у которых не выявлены опухоли, составила в чернобыльской группе 53,4 %, в киевской несколько выше - 58 % исходного числа крыс, взятых в эксперимент. При анализе средней продолжительности жизни (СПЖ) крыс отмечено ее сокращение на 7 - 8 мес по сравнению с биологической нормой. Так, СПЖ до аварии составляла 28,2±0,6 мес, в киевской группе - 21,6±0,5 мес, в чернобыльской - 20,3±0,8 мес. Сокращение продолжительности жизни животных под влиянием постоянного действия радионуклидов в значительной мере связано с общими повреждающими эффектами радиации на различные системы и органы (табл.1.4.5). Непосредственной причиной гибели крыс чаще всего были воспалительные процессы в легких и кишечнике. Заболевания дыхательной системы у киевских животных составили 43,3 %, в чернобыльской группе - 64,3 %. Характерной особенностью заболеваний дыхательной системы у животных, находящихся в Чернобыль, был высокий процент абсцедирующих, геморрагических пневмоний и кровоизлияний в легкие (35,3 %). У киевских животных на этот вид заболеваний приходится 10,3 %. Заболеваемость органов желудочно-кишечного тракта у крыс, находившихся в Киеве и Чернобыле, в процентном отношении Практически совпадает (26,2 и 26,7 % соответственно). Среди заболеваний желудочно-кишечного тракта, аналогично легочной патологии у чернобыльских животных, преобладали геморрагические, язвенно-некротические колиты и энтероколиты (13,6 % по сравнению с 7,2 % в Киеве). Обращают на себя внимание относительно часто встречающиеся изменения в лимфоидных органах. Гиперплазия лимфоузлов, селезенки, тимуса наблюдалась у 12,7 % киевских животных и у 17,6 чернобыльских. Известно, что геморрагические и язвенно-некротические процессы типичны для воздействия на организм больших доз ионизирующего излучения. Значительное количество животных с данной патологией в чернобыльской группе, при незначительном вкладе внешней гамма-компоненты, может быть обусловлен существенным внутренним облучением инкорпорированными радионуклидами с высокой биологической эффективностью. Таблица 1.4.5 Заболеваемость у крыс, постоянно находившихся на экспериментальных базах в Киеве и Чернобыле (1986 - 1989), %
Органы |
Нозологическая форма |
Киев |
Чернобыль |
Дыхания |
Пневмония, бронхопневмония, плевропневмония, плеврит |
24,6 |
17,7 |
Абсцедируювдая пневмония |
6,3 |
22,2 |
|
Кровоизлияния в легкие и геморрагическая пневмония |
4,0 |
13,3 |
|
Пульмонит |
8,4 |
11,1 |
|
Желудочно-кишечныйтракт |
Дистрофия печени |
7,1 |
6,7 |
Колит, энтероколит |
7,1 |
7,8 |
|
Геморрагический колит |
3,2 |
11,1 |
|
Язвенно-некротический энтероколит |
8,8 |
12,2 |
|
Лимфоидные |
Гиперплазия лимфоузлов |
10,3 |
13,2 |
Гиперплазия селезенки, тимуса |
2,4 |
4,4 |
Для сравнения биологической эффективности радиации разного качества представляется целесообразным привести результаты, полученные при воздействии на животных редко и плотноионизирующего излучения. Так, однократное тотальное облучение животных гамма-квантами60Со в дозе 2,0 Гр приводит к смещению начала гибели животных к 16-му месяцу жизни. Быстрые нейтроны со средней энергией 22 МэВ, полученные при бомбардировке протонами бериллиевой мишени на изохронном циклотроне У-240, смещают этот срок к 12-му месяцу. Нейтроны спектра деления, полученные на атомном реакторе ВВР-М, со средней энергией 1,2 МэВ, вызывают начало гибели животных на 8-м месяце жизни. Вероятность гибели у них относительно невелика, поскольку эти виды радиации вызывают значительные канцерогенные эффекты. Однако по критерию начала гибели животных радиационные условия в Киеве и Чернобыле соответствуют однократному облучению гамма-квантами60Со и быстрыми нейтронами с Е 22 МэВ в дозах 2,0 и 1,0 Гр. 4.2.7. Онкологические эффекты Возникновение опухолей в результате воздействия ионизирующих излучений относится к отдаленным радиационным эффектам. В связи с тем что латентный период опухолей человека составляет 10-20 лет (за исключением радиационно-индуцированных опухолей щитовидной железы и миелоидных лейкозов у детей), опыты на экспериментальных животных могут оказаться (с определенными ограничениями) весьма полезными для прогнозирования появления радиогенных опухолей у людей. Однако в эксперименте на животных требуется тщательный анализ факторов, которые могут изменить частоту новообразований в данной популяции. Необходимо также иметь достаточный контроль для разграничения радиогенных и спонтанных опухолей. Суммируя представления об индукции радиогенных опухолей [86], выделяют три канала канцерогенного действия излучений. По клеточному каналу осуществляется инициация, а по тканевому и системному - промоция. Авторы считают одним из самых важных механизмов пострадиационную репопуляцию, при которой увеличивается количество трансформированных клеток. Действие радиации по системному каналу опосредовано изменениями в гормональном и иммунологическом статусах. Реализация радиационных эффектов может осуществляться и по пути нарушения системы механической интеграции ткани повреждения мембран, нарушения ионного гомеостаза и ферментативной активности, адгезивных свойств клеток, что в конечном итоге может приводить к их бласттрансформации. Уникальность аварии на ЧАЭС, не сопоставимая со взрывами атомных бомб в Хиросиме и Нагасаки, заключается в выбросе огромной массы широкого спектра радионуклидов, а также топливных и конденсационных "горячих" частиц, выпавших на огромных территориях. С течением времени внешний гамма-фон и концентрация радионуклидов в окружающей среде в загрязненных районах снизились и продолжают уменьшаться. В настоящее время как люди, так и животные, находящиеся в загрязненных районах, получают ~ 80 % дозы за счёт внутреннего облучения (главным образом с пищей) такими радионуклидами, как137Cs,134Cs,144Ce,106Ru,90Sr,., а в 30-километровой зоне и238Pu,239Pu и241Am. Наибольшую опасность представляют90Sr,239Pu и24lAm. Будучи альфа- и бета-излучателями, они являются источником плотноионизирующего излучения и имеют период полураспада, исчисляемый сотнями и тысячами лет, а стронций и плутоний при определенных условиях активно включаются в обменные процессы в тканях. Создаваемые радионуклидами дозы внутреннего облучения крайне малы и являются по сути Йадфоновыми. Однако постоянно циркулируя в организме или накап-Циваясь в тройных органах, онк постоянно с низкой интенсивностью облучают клетки-мишени, в которых не успевают происходить репарации повреждений ДНК, как в случаях однократного или фракционного облучения. В 50 - 60-Рµ годы в многочисленных экспериментах была выявлена высокая бластомогенная активность целого ряда биологически значимых радионуклидов при их ингаляционном, энтеральном и парентеральном поступлении в организм в больших дозах. Особенности Чернобыльской катастрофы делают весьма сложным моделирование условий облучения на загрязненных территориях. В связи с этим наиболее репрезентативные сведения о канцерогенном действии малых доз излучения низкой интенсивности могут быть получены лишь в натурных экспериментах на диких животных, обитающих в 30-километровой зоне, либо на содержащихся там экспериментальных животных, получающих загрязненные радионуклидами корма. Такие исследования немногочисленны, и если соматические эффекты описаны достаточно подробно, то сведения о канцерогенных эффектах единичны и достаточно противоречивы. Наиболее систематически такие наблюдения проводились на беспородных белых крысах, находящихся в течение жизни в виварии Чернобыля, в сравнении с крысами той же разводки вивария Киева. В первые 3 года после аварии (1986 - 1988) средняя продолжительность жизни (СПЖ) животных в Киеве, и особенно в Чернобыле, резко сократилась по сравнению с доаварийным периодом (1985). Животные погибали главным образом от абсцедирующих геморрагических пневмоний и энтероколитов. Спектр опухолей у обеих групп животных не отличался от спонтанного, с преобладанием доброкачественных опухолей (фиброаденом) молочных желез, которые у чернобыльских крыс появлялись раньше и чаще имели признаки малигнизации. Среди других опухолей доминировали злокачественные опухоли лимфоидной и соединительной ткани (лимфосаркомы и злокачественные гистиоцитомы) легких и (реже) кишечника. При расчете по методу Рейсига с поправкой на интеркуррентную смертность к 20-му месяцу жизни вероятность возникновения опухолей у животных той же разводки составляла в 1985 г. - 5 %, а в 1986 - 1988 гг. у крыс киевской группы - 17 %, a y крыс чернобыльской группы - 35 %. Первые опухоли выявлялись в 1985г. через 16 мес, а в послеаварийном периоде в Киеве - на 14-м, в Чернобыле - через 10 мес. В течение последующих 3 лет (1989 - 1992) количество животных с опухолями составило в киевской и чернобыльской группах 74 %. Как и в первый период после аварии в обеих группах доминировали доброкачественные опухоли молочной железы (ОМЖ) - фиброаденомы с признаками малигнизации у 9,5 % животных в Киеве и 14,7 % в Чернобыле. ОМЖ были множественными: у 27 % животных в Киеве и у 29 % - в Чернобыле; с признаками малигнизации у 9,5 % животных в Киеве и у 14,7 % в Чернобыле. В последней группе ОМЖ значительно чаще сочетались с опухолями других органов (20,25 % в Киеве и 58,8 % - в Чернобыле). За счет этого коэффициент множественности опухолей за все время жизни крыс был в Киеве 1,55, а в Чернобыле 2,52. В первые 3 года после аварии опухоли тимуса у крыс киевской группы не обнаружены, а у чернобыльской они составляли 4,8 % всех опухолей. В 1989 - 1992 гг. тимомы в Киеве были единичными (2,7 %), но в Чернобыле составляли 15,9 % всех опухолей. Сходные соотношения обнаружены также для аденом коры надпочечников: 6,75 - 43,2 % всех обнаруженных опухолей; опухолей щитовидной железы (главным образом аденом) - 15,7 % в Киеве и 43,2 % в Чернобыле; b -клеточных аденом островков Лангерганса поджелудочной железы - 1 % в Киеве и 34,1 % в Чернобыле. Число лимфосарком и злокачественных гистиоцитом легких и кишечника у животных киевской группы было несколько больше, чем у чернобыльской (14,9 и 10 %). Однако у крыс чернобыльской группы появились единичные аденокарциномы легких, главным образом из бронхиальных желез, и плоскоклеточный рак (3,34 %), не наблюдавшиеся у крыс киевской группы и не описанные ранее как спонтанные опухоли у животных этой разводки. Анализ приведенного материала позволяет говорить о развитии характерных для данного вида животных спонтанных опухолей в более ранние сроки (в первые 3 года после аварии) с незначительным увеличением числа малигнизированных ОМЖ, что сохраняется и в последующие 3 года. При изучении особенностей спонтанного канцерогенеза в 1989 - 1992 гг. наиболее характерным отличием у крыс чернобыльской группы является более выраженное, чем у крыс киевской группы, развитие на 2-м году жизни опухолей эндокринных органов в сочетании с ОМЖ и появление аденокарцином и плоскоклеточного рака легких. У крыс обеих групп к настоящему времени характерно развитие гипоплазии костномозгового кроветворения с развитием у отдельных животных гемобластозов (системных ретикулезов). Таким образом, есть основание утверждать, что после аварии на ЧАЭС число спонтанных опухолей у крыс увеличилось как в Чернобыле, так и в несколько меньшей степени в Киеве, появились эпителиальные опухоли легких. Трудности разграничения радиогенных и спонтанных опухолей у крыс, а также наличие в окружающей среде и пище бластомогенных факторов другой природы не позволяют в настоящее время четко выделить вклад ионизирующей радиации в интенсификацию спонтанного канцерогенеза, однако несмотря на очень малые поглощенные дозы облучения, этот вклад несомненно имеет место. 4.2.8. Высшая нервная деятельность в условиях изолированного и комбинированного воздействия ионизирующей, неионизирующей радиации и стресса Центральная нервная система (ЦНС) и высшая нервная деятельность (ВНД) считаются относительно устойчивыми к воздействию ионизирующей радиации, поскольку летальность в первую очередь определяется состоянием кроветворной системы и желудочно-кишечного тракта. Однако, как показали клинические исследования лиц, подвергшихся воздействию радиации в результате чернобыльской аварии, в условиях малых доз облучения функциональные нарушения со стороны основных регуляторных систем организма, включая ЦНС и ВНД, являются ведущими. В реальных условиях влияние ионизирующей радиации происходит на фоне ряда других биологически активных факторов, которые также в первую очередь поражают ЦНС. Наиболее распространенными из них являются психо-эмоциональный стресс и неионизирующие излучения. Для оценки эффектов изолированного и комбинированного воздействия ионизирующей, неионизирующей радиации и стресса на функциональное состояние ЦНС были применены три метода изучения ВНД животных - радиальный лабиринт, челночная камера и камера Скиннера. Причем использовались различные варианты экспериментов, показатели которых отражали двигательную, поисково-исследовательскую активность, краткосрочную и долгосрочную память, процессы выработки и угашения условных рефлексов как в условиях выполнения простых, так и сложных инструментальных задач. Такой подход позволил оценить наиболее подверженные воздействию исследуемых факторов показатели ВНД, роль мотивации в выявлении этих воздействий и наиболее чувствительные схемы постановки эксперимента. Состояние ВНД белых крыс оценивалось чрез 1, 5, 10, 20 и 30 сут после воздействия. Облучение ионизирующей и неионизиоуюшей радиацией проводилось в обеих последовательностях с интервалом 1 ч. Исследования изолированного и комбинированного воздействия ионизирующей (0,1 - 50 Гр) и неионизирующей(0,25; 0,5; 1 и 10 мВт/см2) радиации на ВНД в течении 30 сут после облучения показали зависимость характера и выраженности изменений от уровня факторов, очередности воздействия и времени после воздействия. Кроме того, выраженность эффектов зависит от уровня мотивации и сложности задачи. Изолированное воздействие гамма-радиации в дозах 0,5 - 50 Гр в течение 30 сут после облучения в целом приводит к угнетению ВНД животных с возможными фазами активации. Малые дозы (0,1 и 0,25 Гр) приводят к активации ВНД животных. Изолированное воздействие неионизирующей радиации в течении 3 ч при плотности потока мощности 10 мВт/см2активируют ВНД животных в ранние сроки после облучения (до 5 сут), которая сменяется тенденцией к угнетению через 25 сут после облучения. Более низкие уровни микроволн не вызывают выраженных изменений в поведении животных. При сравнительно высокой плотности потока мощности (10 мВт/см2) и дозе 2 Гр комбинированное воздействие в обеих последовательностях не приводит к качественной модификации эффектов изолированного влияния этих видов излучений. Можно лишь отметить некоторое увеличение степени выраженности изменений у животных, подвергнутых воздействию в последовательности гамма+МВ. При плотности потока мощности 1 мВт/см2и дозе 1 Гр комбинированное воздействие в обеих последовательностях приводит к усилению эффектов, которые при изолированном воздействии наблюдаются лишь на уровне тенденции. Однако предварительное воздействие неионизирующей радиацией, увеличивая угнетающее действие на 5-е сутки после облучения, способствует нормализации показателей через 30 сут. Воздействие в последовательности гамма+МВ, несколько снижая угнетающее действие через 5 сут после воздействия, усиливает процессы активации через 30 сут. При плотности потока мощности 0,5 мВт/см2и дозе 0,5 Гр предварительное воздействие неионизирующей радиацией оказывает активирующее влияние на ВНД, что нейтрализует эффекты гамма-облучения через сутки после воздействия. Комбинированное действие в последовательности гамма+ +СВЧ не приводит к существенной модификации эффектов, наблюдаемых при изолированном воздействии ионизирующей радиации. При плотности потока мощности 0,25 мВт/см2и дозе 0,25 Гр комбинированное действие ионизирующей радиации в последовательности СВЧ+гамма стимулирует условно-рефлекторное поведение крыс в 1-е сутки после облучения, которое заключается в общей активации ЦНС. Комбинированное действие ионизирующей и неионизирующей радиации в последовательности гамма+СВЧ ухудшает рабочую память крыс через 30 еут после облучения. Последовательность воздействия факторов оказалась существенной и при структурно-функциональных исследованиях ЦНС крыс через 1,7 ч после раздельного и комбинированного облучения СВЧ-полем (300 мВт/см2) и гамма-радиацией (50 Гр). При воздействии микроволн после ионизирующего облучения морфологические эффекты тяготеют в сторону усиления, практически не выявляются признаки компенсации со стороны ультраструктуры клеток. Показатели поведения животных при комбинированном воздействии гамма-облучения и кратковременного (2 ч) иммобилизационного стресса также указывают на наличие взаимодействия между этими факторами, которое при дозе 0,5 Гр приводит в целом к угнетению ВНД (увеличение времени выполнения задачи, снижение двигательной активности и долговременной памяти, увеличение количества ошибок). В целом облучение приводит к снижению устойчивости ВНД к воздействию стрессовых факторов. Проведенные исследования позволяют сделать ряд выводов методического характера. Наиболее чувствительными к воздействию ионизирующей радиации являются сложные формы поведения животных при среднем уровне мотивации. Низкие уровни мотивации приводят к большой дисперсии результатов, которая маскирует эффекты. Применение высоких уровней мотивации (например, применение электрошока) приводит к тому, что помимо изучаемых факторов, животное находится под воздействием дополнительного стресса, который может модифицировать или маскировать эффекты основных факторов. Полученные результаты говорят о полидисперсном характере воздействия ионизирующей радиации на ВНД. Последнее особенно справедливо в случае комбинированного воздействия с другими биологически активными факторами окружающей среды.
4.3. Экспериментально апробированные средства модификации ранних и отдаленных радиобиологических эффектов 4.3.1. Антирадикальные и антиоксидантные средства Проблема профилактики (защиты) и лечения острых лучевых поражений для радиобиологии не нова, она бурно развивалась в первые десятилетия после применения американцами ядерного оружия в Хиросиме и Нагасаки в 1945 г. Были открыты и изучены основные классы радиопротекторов, разработаны схемы комплексной терапии острой лучевой болезни. Причем возможности прогресса в этих областях оказались в большой степени исчерпанными к началу 80-х годов, о чем свидетельствовало резкое сокращение числа исследований. Новизна ситуации, сложившейся после Чернобыльской катастрофы, связана с необходимостью поиска средств защиты (профилактики) и лечения от нелетальных лучевых повреждений, ранних и отдаленных, возникающих при длительном низкоинтенсивном облучении. Для этих условий богатый опыт, накопленный радиобиологией, и препараты, эффективные при остром летальном облучении, оказались практически непригодными. Поиск и изучение средств защиты и терапии, эффективных для этих условий, пришлось начинать по сути заново, с использованием принципиально иных экспериментальных моделей и показателей эффективности. Было установлено, что при значительном снижении доз и мощностей доз радиации типичные для острого летального облучения клинические синдромы (костномозговой, кишечный) либо совсем отсутствуют, либо приобретают атипичную затяжную форму. Важную роль приобретают явления радиационного иммунодефицита, геморрагический синдром, вегетососудистая дистония, нарушения нейроэндокринной регуляции. На клеточном уровне на первый план выдвигаются нарушения структуры и функции биомембран, обусловленных активацией ПОЛ. С учетом этих особенностей клиники постчернобыльских поражений серьезную перспективу в качестве средств профилактики и ранней патогенетической терапии имеют препараты антирадикальной и антиоксидантной активности, адаптогены и иммуностимуляторы (модуляторы), особенно если это нетоксичные вещества природного происхождения и если все эти эффекты соединяются в одном препарате. 1. Растительные фенольные соединения со-,n- и рядовым расположением гидроксильных групп в бензольном кольце - флавоноиды (рутин, кверцетин, катехины, антоцианы), фенольные кислоты (галловая, эллаговая, оксикоричные), производные пирокатехина, пиррогалола, гидрохинона - отличаются низкой токсичностью, хорошо растворимы в воде (кроме рутина, кверцетина), обладают АО-антирадикальной, капилляроукрепляющей ("Р-витаминной") активностью, образуют комплексы с тяжелыми металлами, обладают также иммуномодуляторным, гепатозащитным, антитоксическим действием. Эти препараты успешно устраняют или смягчают проявления геморрагического синдрома, явления иммунодефицита, нарушения эндокринной регуляции (смягчают проявления лучевого гиперкортицизма, активизируют функцию тимуса). В основе большинства этих эффектов лежит ограничение лучевой активации ПОЛ клеточных мембран. Фенольные соединения - физиологические синергисты аскорбиновой кислоты в антиоксидантном, сосудоукрепляющем, противолучевом эффектах, и их предпочтительнее использовать в составе комплексных препаратов типа аскорутина, галаскорбина. Наиболее эффективен препарат в составе комплекса катехинов и аскорбата. Еще более перспективно использование АО-комплексов, включающих, кроме указанных препаратов, еще и токоферол, каротиноиды. Природные аналоги таких комплексов - масло облепихи, шиповника, препараты солодки. Препараты в таких комплексах могут применяться длительно как внутрь, так и местно в виде мазей при локальных повреждениях кожи и слизистых оболочек, например, при осложнениях лучевой терапии злокачественных новообразований. Антиоксиданты и их комплексы наиболее эффективны при непродолжительных курсах применения в дозах, близких к физиологическим. В повышенных дозах они легко окисляются в жидкостях организма, выступая уже как факторы прооксидантного действия. А при длительном поступлении в организм они могут в определенной мере ингибировать продукцию собственных эндогенных АО, так что совокупный эффект может оказаться отрицательным, прооксидантным. В комплексах АО взаимно стабилизируют друг друга, и опасность прооксидантного действия снижается. Необходимо избегать длительного введения АО в дозах, многократно превышающих физиологическую потребность в них. Преимуществом природных АО является возможность приема их через рот, в том числе в виде пищевых добавок, лечебных пищевых продуктов. Доказана перспективность применения в качестве АО пищевых добавок продуктов переработки винограда, чая, кофе, какао, фруктов и овощей. Получены эффективные противолучевые добавки из виноградных выжимок, оболочки какао-бобов, из грубого чайного листа[15]. Комплексы природных АО успешно используются в лечении местных лучевых повреждений кожи и слизистых оболочек при воздействии бета-радиоактивной пыли, при лучевой терапии опухолей (масло облепихи, шиповника, препараты фенолов, прополиса, АО-композиции [14,15]. Полифенолы (меланины, таннины, гуминовые кислоты) обладают эффективностью более простых фенольных соединений, но, в отличие от последних/плохо растворимы в воде, а при парентеральном введении токсически действуют на печень, вплоть до канцерогенного эффекта. Поэтому их следует применять главным образом местно. Но полученные на их основе растворимые препараты: - соли гуминовых кислот, продукты гидролиза меланина, гидрофильные фенолы прополиса.- могут успешно вводиться и внутрь. 2. Альфа-токоферол - жирорастворимый АО, стабилизатор липидного бислоя мембран, инактиватор окислительных радикалов О2., ОН-максимально эффективен после встраивания в структуру биомембран, на что требуется не менее 18 ч от момента его введения. Подобно витамину А токоферол транспортируется специализированными белками и частично депонируется в печени. Но при избыточном поступлении может вести себя как прооксидант. Применение токоферола максимально эффективно в комплексе с другими АО - ретинолом, каротином, аскорбиновой кислотой - и в составе природных комплексов. 3. Синтетические АО. Такие пищевые АО, как ионол (дибунол, бутилгидрокситолуол),пропилгаллат, бутилгидроксианизол, а в дальнейшем производные 3-оксипиридина, стали применяться как противолучевые средства. Недостаток ионола - слабая растворимость в воде. Ныне синтезированы и успешно используются АО - производные 3-оксипиридина (эмоксипин), которые водорастворимы. Синтетические АО при длительном введении могут ингибировать синтез эндогенных АО, что в итоге может снижать суммарную АО-активность тканей. Поэтому ионол и другие синтетические АО могут применяться в качестве противолучевого средства местно либо короткими курсами при остром облучении. Длительно он может вводиться в минимальных дозах (3-5 мг/кг) и в составе композиций с природными АО. 4.3.2. Адаптогены В механизме биологического действия ионизирующей радиации, особенно низкоинтенсивной, неспецифический (стрессорный) компонент играет весьма существенную роль. Применение адаптогенов в качестве противолучевых средств обоснованно и подтверждено экспериментально. В силу участия продуктов ПОЛ в механизме стресса [13,14] противолучевой эффект адаптогенов одновременно является и антистрессорным. 1. Аралиевые (женьшень, элеутерококк, заманиха), китайский лимонник. Эти адаптогены - признанные стимуляторы физической и умственной работоспособности, общей резистентности, в частности, в поле постоянного низкоинтенсивного лучевого воздействия. Обладают как радиозащитным, так и ранним лечебным действием [15]. В механизме действия женьшеня и элеутерококка следует выделить наряду с антиоксидантным также антиглюкокортикоидный эффект - способность уменьшать лучевую реакцию гиперкортицизма и, тем самым, ослаблять вызываемое радиацией опосредованное угнетение пострадиационного восстановления в органах кроветворения и иммуногенеза, стимулировать функцию тимуса и других эндокринных желез. Антирадиационная эффективность женьшеня и других адаптогенов показана не только экспериментально; имеются и клинические наблюдения за рабочими урановых рудников, радиологами (Китай, Северная Корея). 2. Препараты из морепродуктов. МИГИ-К - препарат из мидий, экстракт, содержащий растворимые белки, углеводы, набор АО-витаминов и микроэлементов. Нетоксичен, имеет высокую пищевую ценность. Обладает выраженной противолучевой эффективностью при введении как до, так и в процессе длительного низкоинтенсивного лучевого воздействия. В эксперименте и в условиях применения в 30-километровой зоне (участники ликвидации Чернобыльской катастрофы) проявил себя как эффективное противолучевое средство [87]. Из мидий получены и другие эффективные препараты (белково-углеводный концентрат БУК-М), используемые как противорадиационные пищевые добавки [15]. Сходной АО и противолучевой активностью обладают несколько биологически активных соединений (этинохром) и препаратов, выделенных из морских ежей, моллюсков, водорослей [15]. Альгинаты наряду со связыванием и выделением радионуклидов оказывают и умеренно выраженное адаптогенное действие. 3. Высокогорье, нормобарическая прерывистая гипоксия. Пребывание в условиях высокогорья и среднегорья (1000 - 3000 Рј РЅ.у.м.) вызывает в организме реакцию по типу мягкого стресса, активизируя эндогенные (в том числе и АО) механизмы защиты. Период острой адаптации (1-3 сут) сопровождается кратковременным снижением общей и противолучевой устойчивости организма. По мере формирования долговременной адаптации к условиям высокогорья неспецифическая резистентность и противолучевая устойчивость закономерно возрастают, достигая максимума на 30 - 60-Рµ сутки. Эффекты длительного нелетального облучения устраняются практически полностью за счет стимуляции кроветворения, АО систем защиты, иммунореактивности. Наряду с экспериментальными данными, имеется положительный клинический опыт долечивания участников ликвидации Чернобыльской катастрофы, онкологических больных после лучевой терапии, а также вторичных анемий и иммунодефицитов, хронических бронхитов и аллергических состояний. Среди лечебно-оздоровительных факторов высокогорья наиболее важны гипоксия, инсоляция с высоким УФ-компонентом, чистота и низкая бактериальная загрязненность воздуха с высоким содержанием легких отрицательных аэроионов, свежеталая и ледниковая вода. В Киеве созданы установки горного климата, моделирующие в условиях равнины этот комплекс факторов и также используемые для оздоровления участников ликвидации Чернобыльской катастрофы. Барокамерная и нормобарическая прерывистая гипоксия также моделируют лечебный эффект важнейшего фактора высокогорья. Дыхание гипоксическими газовыми смесями с содержанием 8 - 10 % кислорода (ГТС-8, ГГС-10) оказывает выраженное противолучевое действие в эксперименте, успешно используется в онкологической клинике для снятия лучевых реакций и осложнений, находит применение в лечении неврологических, кардиологических, респираторных, иммунологических последствий длительного облучения. 4. Ультрафиолетовое (УФ)-облучение. Использование УФ-излучения как средства повышения общей неспецифической резистентности (адаптогена) опирается на весьма многочисленные экспериментальные и клинические данные, воспроизводит эффект еще одного оздоровительного фактора высокогорья. УФ(световое)-голодание в условиях Крайнего Севера, у шахтеров и т.п. приводит к прогрессирующему снижению устойчивости организма к вредным факторам, росту заболеваемости. В основе общестимулирующего эффекта УФ-облучения лежит мобилизация эндогенных механизмов защиты и резистентности, сшмуляция кроветворения, иммунореакгивности, работоспособности и тл. Однако такое действие наблюдается под влиянием низких (субэритемных или пороговых эритемных) доз УФ-радиации, тогда как более высокие дозы оказывают обратное действие. Курс субэритемных УФ-облучений областей А и В (290 - 400 нм) стимулирует выведение организма тяжелых металлов, химических вредностей и радионуклидов. Локальное УФ-облучение интактного участка кожи по схеме (10 сеансов, 10 - 16 биодоз суммарно) существенно ослабляет эффект общего и местного ионизирующего облучения, ускоряет заживление лучевых язв у онкологических больных после лучевой терапии. В последние годы широко применяется УФ- (а также лазерное) облучение аутокрови (АУФОК). Образующиеся при этом в облученной крови активные формы кислорода при возврате в циркуляцию стимулируют физиологические АО-системы организма, кроветворение, клеточные и гуморальные факторы иммунитета, ускоряют реабилитацию больных после лучевой и цитостатической терапии, что особенно важно для больных, подвергшихся ранее длительному низкоинтенсивному облучению. Красное излучение гелий-неоновых и аргоновых лазеров успешно используется для лечения ран, трофических язв, ожогов, операционных травм. 4.3.3. Лазерное излучение как средство иммунокоррекции радиационных поражений организма Как известно, под влиянием проникающей радиации снижается защитная реактивность организма, подавляется система иммунитета вообще и противоопухолевого, в частности развивается депрессия кроветворения. Поэтому среди путей повышения эффективности реабилитационной терапии лиц с радиационными поражениями важное место занимает поиск подходов, приводящих к иммуномодуляционным сдвигам и стимулирующих гемопоэз. Одним из применяемых в последние годы средств модификации ранних и отдаленных последствий влияния на организм проникающей радиации является низкоэнергетическое (неповреждающее) излучение гелий-неонового (Не-Ne) лазера красной области спектра (-0,63 мкм). Данные о радиопротекторной активности красного лазерного света (КЛС) были получены в опытах in vitro, а также in vivo при воздействии на экспериментальных животных. Радиозащитное действие КЛС было отмечено на бактериях E.coli, на клетках культуры ткани китайского хомячка, кожно-мышечной ткани эмбриона человека, в культуре нормальных и трансформированных фибробластов, клетках опухолевой ткани HeLa, на регенерирующей скелетной мышце. КЛС стимулировал пострадиационные регенераторные процессы в организме животных, оказывал радиопротекторное действие на кроветворные и эпителиальные ткани облученных крыс, тормозил развитие дистрофических процессов селезенки мышей и локально облученной печени крыс, увеличивал выживаемость облученных животных. Предприняты попытки использования радиозащитного действия He-Ne лазера в клинической онкологии, в частности для профилактики острых лучевых реакций кожи при нейтронной терапии опухолей головы и шеи, в лечении послеоперационных гнойных осложнений у онкологических больных, получавших лучевую терапию. Наряду с поверхностным (местным) действием недавно был предложен новый метод лазерной биостимуляции организма - внутривенное лазерное облучение крови (БЛОК). Суть метода заключается в прямом действии излучения He-Ne лазера на циркулирующую кровь. Излучение направляется в кровеносный сосуд по тонкому световоду (d= 0,2 - 0,4 РјРј), один конец которого присоединен к лазеру, а другой с помощью инъекционной иглы введен в сосуд. Было изучено иммуномодулирующее действие БЛОК на такой показатель состояния неспецифического иммунитета, как фагоцитарная активность (ФА) макрофагов у мышей, которые подвергались действию гамма-излучения (0,5 Гр). На 5-е сутки после облучения ФА мышей в контрольной группе (только гамма-излучение) возрастала на 20 - 30 %, а в группе животных, подвергнутых сочетанному действию (0,5 Гр+ВЛОК) - на 50 %. На 14-е сутки после облучения в контрольной группе было отмечено значительное (до 70 % по отношению к исходному уровню) снижение ФА, тогда как при сочетанием воздействии уровень ФА был на 10 % выше исходного. Получены данные, свидетельствующие об иммунокорригирующем эффекте БЛОК и при длительном воздействии на организм малых доз ионизирующей радиации. Опыты были выполнены на мышах, половина из которых в течение 1 мес содержалась в виварии Чернобыля. У контрольных животных, находившихся в виварии Киева, БЛОК вызывал незначительную (до 7 %) стимуляцию естественной киллерной активности. В опытной же группе БЛОК приводил к значительному (троекратному угнетению изначально повышенной цитотоксической активности. Сходным образом изменялась под влиянием БЛОК и фагоцитарная активность макрофагов. Принимая во внимание данные о стимулирующем действии излучения He-Ne лазера на иммунокомпетентные клетки in vitro, изучался иммунный ответ организма при адаптивном переносе лазер-активированных лимфоидных клеток. Было показано, что при переносе подвергнутых лазерному воздействию in vitro спленоцитов от ин-тактных мышей-доноров, летально облученным (8 Гр) реципиентам с одновременной иммунизацией последних эритроцитами барана, количество антителообразующих клеток к эритроцитам в селезенке реципиентов почти вдвое превышало контрольный уровень (при переносе интактных клеток). Важными представляются данные об улучшении некоторых показателей гемопоэза при лазерном облучении крови in vitro (БЛОК). Так, при исследовании колониеобразующей способности клеток костного мозга методом экзогенного колониеобразования установлено, что проведение БЛОК у мышей, подвергнутых летальному облучению (8 Гр), приводило к статистически значимому повышению количества колоний в селезенке животных. На основании совокупности экспериментальных данных об иммуностимулирующем эффекте лазерного света, его способности активировать процессы кроветворения, а также об определенном противоопухолевом действии были предприняты первые попытки применения БЛОК с целью профилактики и лечения осложнений радиотерапии у больных раком шейки матки и молочной железы, у которых наблюдались депрессия кроветворения и угнетение системы иммунитета. После курса БЛОК улучшалось общее состояние этих больных, отмечалось повышение количества эритроцитов и лейкоцитов, на 30 % возрастала фагоцитарная активность лейкоцитов. Существенно увеличивалась пролиферативная активность Т-лимфоцитов, а также исходно весьма низкая активность натуральных киллеров. Наблюдалось изменение соотношения субпопуляций Т-лимфоцитов со снижением доли хелперов-индукторов и одновременным ростом киллеров-супрессоров. БЛОК вызывало повышение на 30 - 40 % насыщения крови кислородом. Все эти изменения наблюдались у пациентов на фоне продолжающейся химиолучевой терапии и удерживались спустя 1-2 нед после завершения курса БЛОК. При этом отмечалось значительное снижение частоты постлучевых реакций: эпителиитов, ректитов, циститов и др. БЛОК было применено также при лечении хронического аутоиммунного тиреоидита у лиц, принимавших участие в ликвидации последствий аварии на ЧАЭС и проживавших на территории, загрязненной радионуклидами. Через 3 мес после проведения ВЛОК-терапии у больных достаточно уменьшались объем щитовидной железы и концентрация антител к тиреоглобулину, снижался уровень тиреотропного гормона гипофиза, увеличивалось содержание общего тироксина, состав Т-лимфоцитов изменялся в пользу супрессоров. Излечение тиреоидита было зарегистрировано в 21,8 % случаев, а положительные изменения в структуре и функции щитовидной железы отмечены у 84,4 % больных. Таким образом, как экспериментальный материал, так и первые клинические наблюдения свидетельствуют о том, что методы лазерной стимуляции организма способны найти свое место в комплексной реабилитационной терапии лиц, подвергшихся лучевому воздействию.
4.4.
Радиобиологические эффекты у
растений
4.4.1. Накопление радионуклидов
в компонентах фитоценоза
Особенностью
радионуклидного загрязнения, связанного
с Чернобыльской катастрофой, является
разнообразие химических форм агрегатных
состояний, выброшенных в окружающую
среду радиоактивных элементов. Часть
элементов была выброшена в водорастворимом,
ка-пельно-жидком состоянии, часть же в
виде "горячих" частиц, причем
соотношение этих форм не является
постоянным, а изменяется под влиянием
биотических и абиотических факторов
среды. В этих условиях накопление
радионуклидов растениями, происходящее
в основном за счет водорастворимой и
обменной форм компонентов загрязнения,
отражает весьма сложные переходные
процессы в почве, скорость и направленность
которых определяется биологической
активностью всех компонентов
корнеобитаемого слоя.
Массовый отбор
почвы, растительности, произведенный
летом 1986 г. по всей территории Украины,
и последующий их гамма-спектрометрический
анализ позволил выделить ряд количественных
и качественных особенностей в характере
загрязнения различных районов, в
соответствии с которыми территория
Украины может быть условно разделена
на три зоны: ближняя зона радиусом 10 -
20 км от ЧАЭС; промежуточная зона - от 20
до 150 км; дальняя зона - свыше 100 - 150 км от
ЧАЭС [124]. Конфигурация этих зон
характеризуется различной степенью
вытянутости по направлениям. По уровню
суммарной активности основных продуктов
деления (цезия, церия, циркония, бария)
в почве они различаются следующим
образом: n·10-7-
n·10-8Ки/кг
- дальняя зона; n·10-9-
n·10-10Ки/кг
- промежуточная зона; n·10-7-
n·10-5и
более Ки/кг - ближняя зона. Наряду с
количественными различиями по уровню
суммарной активности радиоактивное
загрязнение почвы и фитомассы в этих
зонах имеет ряд особенностей как по
составу, так и по характеру поглощения
радионуклидов структурными компонентами
фитоценоза, которые имеют принципиальное
значение с точки зрения формирования
дозовых нагрузок на растительные
сообщества, за счет включения их в
продукты питания, в том числе кормовые
и зерновые культуры, приводящие к
заражению молока, мяса и
хлебопродуктов.
Результаты исследования
накопления радионуклидов в растениях,
характеризующихся морфологическими
особенностями видовой принадлежности
и произрастающих на реперных участках
с различными уровнями радиоактивного
загрязнения, показали [34,56], что существуют
межвидовые и даже внутривидовые различия
в накоплении радионуклидов в различных
частях растений. Амплитуда колебаний
значений уровней активности смеси
гамма-излучателей для разных культур
находилась в пределах 2 - 5-кратных
различий, а для люпина (Lupinus luteus L.)
достигала даже 20-кратной величины.
Динамика накопления радионуклидов
многолетними кормовыми травами (люцерна
(Medicago sativa L.), люпин) характеризовалась
резким спадом удельной активности в
1987 г. по сравнению с 1986 г. и менее
значительным - в последующий период.
Более высокой степенью накопления
радионуклидов характеризуется корневая
система по сравнению с надземной частью
растения в целом как у луговых, так и у
злаковых культур (различия от 1,3 до
7-кратных). Среди органов надземной части
растений наибольшей концентрацией
радионуклидов отличались листья и
наименьшей - репродуктивные органы
(соцветия). Таким образом, накопление
радионуклидов растениями определяется
не только их количеством в почве
конкретного региона, но и
анатомо-морфологическими и физиологическими
особенностями растительного организма,
типом корневой системы, степенью ее
развития и глубиной проникновения в
почву.
Лесная растительность обладает
большой поглотительной емкостью по
отношению к радионуклидам, что связано
с наличием значительно и сильно
расчлененных поверхностей (листья,
хвоя, мелкие ветви). Так, надземная часть
сосново-березового леса (типичные
участки Полесья) задержала более 40 %
различных выпадений (90Sr,137Cs,144Се),
сосновый прирост - 90 %, густые сосновые
насаждения - почти 100 %. Большая
поглотительная способность хвойных
пород связана и с тем, что поверхность
многолетней неопадающей хвои круглогодично
концентрирует радионуклиды.
Лесные
насаждения 30-километровой зоны вокруг
ЧАЭС представлены главным образом
молодняками и средневозрастными
растениями (57,3 и 33,3%). Средний возраст
насаждений 35 лет. Наиболее распространенные
лесообразующие породы - сосна обыкновенная
(Pinus sylvestris), дуб (Quercus robur), береза (Betula
pendula), осина (Populus tremula), ольха (Ainobetula
rubra).
Анализ полученных в 1986 г. данных
по вертикальному распределению
радионуклидов на хвойных древостоях
позволил выявить следующие закономерности:
деревьями 20 - 25-летнего возраста,
произрастающими на территории, прилегающей
к ЧАЭС, крупнодисперсные аэрозоли
(частицы более 50 мкм) практически не
задерживались и оседали непосредственно
под пологом леса, из них 30 % - в лесной
подстилке. В надземной фитомассе
древесного яруса задерживалось около
20 % радионуклидов. Значительно загрязнена
поверхность коры, особенно той части,
которая обращена к источнику выброса.
Эффективность захвата аэрозолей
надземными органами растений зависела
как от физико-химических параметров
выпадающих аэрозолей, так и от устройства
листовой поверхности и морфологического
строения этих растений.
Радиоактивные
аэрозоли размером менее 50 мкм селективно
адсорбировались (захватывались)
волосками, содержащими смолистые
вещества, расположенными на поверхности
листа растений. Свыше 30 % общего количества
'продуктов деления, удерживаемого
растительностью, связано с аэрозолями
размером менее 4 мкм. Данные об уровнях
загрязненности фитомассы многолетних
растений и динамике ее снижения
свидетельствуют о превалирующем вкладе
некорневого механизма загрязнения до
1988 г. и о постоянном увеличении вклада
корневого поступления с 1988 г. по настоящее
время.
В первый год после аварии в
надземной фитомассе хвойных лесов
содержалось 18,5 %137Cs
и 6,7 % -90Sr.
Причем, в подстилке находилось 64,5 и 83,2
% соответственно, а в почве 17,0 и 10,1 %
общего количества.
Полученные данные
свидетельствуют, что к 1990 - 1992 гг. в
верхний слой почвы из подстилки
мигрировало около 25 % радионуклидов. В
травянистом покрове под пологим хвойного
леса за 5 лет содержание90Sr
уменьшилось более чем в 2 раза, a137Cs
- в 7,5 раза. В общей форме соотношение
активности, выраженное в процентах от
сухой массы, выглядит так:
хвоя
(листья) > мелкие ветви > большие ветви
> стволы.
В 1992 г. увеличилось корневое
поступление137Cs
в различные органы и ткани. Причем
интенсивность поступления зависит от
степени загрязнения почвы полигонов.
Наиболее интенсивное поступление
отмечено у растений, произрастающих на
полигонах с.Новошепеличи (150 Ки/км2)
и с.Буряковка (70 Ки/км2).
На
рис. 1.4.1, 1.4.2 представлены результаты
радиоавтографии ветвей хвойных и
лиственных пород разного возраста,
отобранных на фиксированных полигонах.
Если в 1986 - 1987 гг. регистрировалось только
поверхностное загрязнение фитомассы,
то автографы 1992 г. свидетельствуют о
значительном включении радионуклидов
непосредственно в ткань хвои и
листьев.
Полученные в 1992 г. данные об
уровнях загрязнения фитомассы, о
топографии распределения радионуклидов
по органам растений свидетельствуют о
значительном повышении вклада корневого
поступления в фитомассу. Эта закономерность
особенно проявляется у растений,
произрастающих на территории с уровнем
загрязнения более
100 Ки/км2.
Эти данные свидетельствуют о необходимости
строгой дифференциации территории зоны
при разработке мероприятий по уходу за
лесными насаждениями.
 Рис.1.4.1.
Радиоавтограф ветви осины 3-летнего
возраста, произрастающей на полигоне
с уровнем загрязнения свыше 200
Ки/км2
Результаты
наблюдений в 1986 - 1992 гг. над характером
вертикальной миграции радионуклидов
по почвенным горизонтам различных почв
на фиксированных полигонах в 30-километровой
зоне ЧАЭС, показали, что независимо от
типа почв и характера надпочвенного
покрова, основные их количества
фиксированы в верхнем 1 - 5-сантиметровом
слое почвы. Однако в последние годы
отмечается тенденция к увеличению
скорости вертикальной миграции,
проявляется зависимость распределения
радионуклидов от генетических особенностей
почв, характера почвенного покрова,
степени увлажненности. Так, в результате
вертикальной миграции к концу
вегетационного периода 1992 г. радионуклиды
в доступных для измерений концентрациях
мигрировали в слой 15 - 20 см минеральной
части почв. Наибольшая скорость миграции
радионуклидов по нисходящей в торфяных,
лугово-болотных, луговых почвах. Низкой
подвижностью137Cs
характеризуется в дерновых слабоподзолистых
глинисто-песчаных почвах.
На занятых
лесами территориях около 85 % радионуклидов
продолжает оставаться в подстилке.
Таким образом, основное количество
радионуклидов сосредоточено в слое
почвы 0 - 5 см. Здесь за весь период
наблюдений запас137Cs
снизился на 12 % , в том числе за последние
2 года на 8 %, а в нижележащих слоях -
увеличился: в слое 5 - 10 см на 1,5 %, в слое
10 - 15 см - на 0,5 %. Миграция137Cs
no профилю почвы происходит приблизительно
на 1 см в год.
Рассчитанные значения
коэффициентов квазидиффузии137Cs
лежат в пределах 1,6·10-9
см2/с
для дерновых слабоподзолистых
глинисто-песчаных почв и 3,5·10-9
см2/с
для торфяных маломощных.
Отсутствие
или наличие лесной подстилки на почве
в сосновом или березовом лесу существенно
отражается на миграции137Cs
и с листового березового или хвойного
соснового опада в поверхностный слой
почвы. Период полупотерь137Cs
из свежего опада, помещенного на почву
с подстилкой. - 5 сут. а на почве без
подстилки - 45 сут.
Рис.1.4.1.
Радиоавтограф ветви осины 3-летнего
возраста, произрастающей на полигоне
с уровнем загрязнения свыше 200
Ки/км2
Результаты
наблюдений в 1986 - 1992 гг. над характером
вертикальной миграции радионуклидов
по почвенным горизонтам различных почв
на фиксированных полигонах в 30-километровой
зоне ЧАЭС, показали, что независимо от
типа почв и характера надпочвенного
покрова, основные их количества
фиксированы в верхнем 1 - 5-сантиметровом
слое почвы. Однако в последние годы
отмечается тенденция к увеличению
скорости вертикальной миграции,
проявляется зависимость распределения
радионуклидов от генетических особенностей
почв, характера почвенного покрова,
степени увлажненности. Так, в результате
вертикальной миграции к концу
вегетационного периода 1992 г. радионуклиды
в доступных для измерений концентрациях
мигрировали в слой 15 - 20 см минеральной
части почв. Наибольшая скорость миграции
радионуклидов по нисходящей в торфяных,
лугово-болотных, луговых почвах. Низкой
подвижностью137Cs
характеризуется в дерновых слабоподзолистых
глинисто-песчаных почвах.
На занятых
лесами территориях около 85 % радионуклидов
продолжает оставаться в подстилке.
Таким образом, основное количество
радионуклидов сосредоточено в слое
почвы 0 - 5 см. Здесь за весь период
наблюдений запас137Cs
снизился на 12 % , в том числе за последние
2 года на 8 %, а в нижележащих слоях -
увеличился: в слое 5 - 10 см на 1,5 %, в слое
10 - 15 см - на 0,5 %. Миграция137Cs
no профилю почвы происходит приблизительно
на 1 см в год.
Рассчитанные значения
коэффициентов квазидиффузии137Cs
лежат в пределах 1,6·10-9
см2/с
для дерновых слабоподзолистых
глинисто-песчаных почв и 3,5·10-9
см2/с
для торфяных маломощных.
Отсутствие
или наличие лесной подстилки на почве
в сосновом или березовом лесу существенно
отражается на миграции137Cs
и с листового березового или хвойного
соснового опада в поверхностный слой
почвы. Период полупотерь137Cs
из свежего опада, помещенного на почву
с подстилкой. - 5 сут. а на почве без
подстилки - 45 сут.
 Рис.
1.4.2. Радиоавтограф сеянцев сосны и
брусники, произрастающих на полигоне
с уровнем загрязнения свыше 200
Ки/км
Скорость разложения свежего
листового опада была в 10 раз больше, чем
в варианте без подстилки.
Миграция
радионуклидов из подстилки почвы в
растения за 5 лет составила около 0,01
%.
Особенности биологического действия
ионизирующих излучений на многолетние
растения в 30-километровой зоне ЧАЭС
связаны с неравномерным распределением
дозовых нагрузок на отдельные части и
органы растений в течение послеаварийного
периода, определяемых процессами
миграции, накопления и перераспределения
радионуклидов как в почве, так и в самом
растении.
Отличительной особенностью
древесных растений является высокая
степень локализации радиочувствительных
тканей. Практически все биологические
эффекты на уровне многолетнего организма
вызваны действием излучения на верхушечную
меристему растений. Даже такие эффекты,
как усиление кущения, интенсивный рост
боковых побегов или пробуждение спящих
почек, связаны с поглощенной дозой в
верхушечной меристеме, поскольку
поражение конуса нарастания проводит
к снятию апикального доминирования.
Поэтому для прогнозирования состояния
многолетних растений в 30-километровой
зоне ЧАЭС необходимо исследовать не
только формирование общей дозовой
нагрузки на биоценоз, но и динамику
формирования дозовых нагрузок на
критические ткани растений (меристемы),
т.е. исследовать геометрию распределения
доз по ярусам многолетних растений.
Для
получения данных об экспозиционной
дозе в разных ярусах Древостоя использовали
термолюминесцентный, биологический и
рентгенографический методы
дозиметрии.
Таблица
1.4.5
Вертикальная
миграция радионуклидов в почве (Бк/кг).
Полигон - с.Новошепеличи, 1991 г.
Рис.
1.4.2. Радиоавтограф сеянцев сосны и
брусники, произрастающих на полигоне
с уровнем загрязнения свыше 200
Ки/км
Скорость разложения свежего
листового опада была в 10 раз больше, чем
в варианте без подстилки.
Миграция
радионуклидов из подстилки почвы в
растения за 5 лет составила около 0,01
%.
Особенности биологического действия
ионизирующих излучений на многолетние
растения в 30-километровой зоне ЧАЭС
связаны с неравномерным распределением
дозовых нагрузок на отдельные части и
органы растений в течение послеаварийного
периода, определяемых процессами
миграции, накопления и перераспределения
радионуклидов как в почве, так и в самом
растении.
Отличительной особенностью
древесных растений является высокая
степень локализации радиочувствительных
тканей. Практически все биологические
эффекты на уровне многолетнего организма
вызваны действием излучения на верхушечную
меристему растений. Даже такие эффекты,
как усиление кущения, интенсивный рост
боковых побегов или пробуждение спящих
почек, связаны с поглощенной дозой в
верхушечной меристеме, поскольку
поражение конуса нарастания проводит
к снятию апикального доминирования.
Поэтому для прогнозирования состояния
многолетних растений в 30-километровой
зоне ЧАЭС необходимо исследовать не
только формирование общей дозовой
нагрузки на биоценоз, но и динамику
формирования дозовых нагрузок на
критические ткани растений (меристемы),
т.е. исследовать геометрию распределения
доз по ярусам многолетних растений.
Для
получения данных об экспозиционной
дозе в разных ярусах Древостоя использовали
термолюминесцентный, биологический и
рентгенографический методы
дозиметрии.
Таблица
1.4.5
Вертикальная
миграция радионуклидов в почве (Бк/кг).
Полигон - с.Новошепеличи, 1991 г.
|
|
|
|
|
|
Общаяактивность, Бк/кг |
Глубина, см |
106Ru, % |
134Cs, % |
137Cs, % |
144Ce, % |
154Eu, % |
|
0-5 |
6,7 |
8,50 |
84,80 |
- |
- |
44172,30 |
5-10 |
- |
7,70 |
92,30 |
- |
- |
285,5 |
0-15 |
- |
- |
82,80 |
17,40 |
- |
187,7 |
5-20 |
- |
- |
63,90 |
36,10 |
- |
76,3 |
20-25 |
- |
- |
100 |
- |
- |
92,1 |
С помощью датчиков, расположенных в ярусах крон представителей голосеменных растений - ель (Picea excelsa), сосна (Pinus sylvestris) - на различной высоте от поверхности почвы, получены данные об изменении дозовых нагрузок на вегетативные апексы в течение вегетационного периода на фиксированных полигонах (села Новошепеличи, Черевач, Залесье, линии электропередач (ЛЭП)), которые свидетельствуют о том, что отмечаются существенные различия в дозовых нагрузках на апексы как по вертикали в пределах каждого растения, так и в пределах каждого из полигонов. Если в первые годы после аварии дозовая нагрузка определялась внешним загрязнением коры и хвои (около 70 % радионуклидов в виде мелких аэрозолей удерживалось обрастающей древесиной, корой, хвоей и только 30 % находилось в листовом опаде и почве), то в настоящее время активность распределилась по исследуемым компонентам следующим образом, %: хвоя, листья - 1 - 3; кора - 4 - 20; почва - 0,2 - 0,4; трава - 4 - 6; подстилка - 78 - 95. Следствием перераспределения дозовых нагрузок явилось резкое усиление восстановительных процессов в апексах верхних ярусов деревьев, что проявилось в значительном уменьшении количества аномалий в формообразовательных процессах. Анализ полученных результатов показал, что максимальную дозовую нагрузку в 1991 г. испытывали точки роста нижнего яруса древостоя, расположенные в непосредственной близости от поверхности почвы. Так, в районе с.Новошепеличи доза облучения меристем апексов верхушечных почек сосны и ели составила 82,3 мГр/год, боковых побегов, расположенных на высоте 50 см - 103,4 мГр/год. Иная закономерность дозовых нагрузок выявлена на полигонах с более низкой плотностью загрязнения. Так, в районе с.Залесье доза облучения почек нижнего яруса составила 28,9 мГр/год, а верхушечных почек - 27,2 мГр/год. Следует отметить, что на высоте первого яруса растений сосны и ели пятилетнего возраста на полигоне с.Новошепеличи мощность экспозиционной дозы более чем на порядок выше, чем в районе полигона с.Залесье (табл. 1.4.6). Таблица 1.4.6 Мощность экспозиционной дозы на различной высоте от поверхности почвы на полигонах 30-километровой зоны ЧАЭС в 1991 г. (24.04 - 16.10)
Высота, см |
с. Залесье |
с. Новошепеличи |
||||||||||||||
Целина |
Пашня |
|||||||||||||||
верх |
середина |
низ |
|
|||||||||||||
мкГр/ч |
мГр/ год |
мкГр/ч |
мГр/ год |
мкГр/ч |
мГр/ год |
мкГр/ч |
мГр/ год |
мкГр/ч |
мГр/ год |
|||||||
250 |
3,1 |
27,2 |
|
|
|
|
|
|
9,4 |
82,3 |
||||||
200 |
3,2 |
28,0 |
|
|
|
|
|
|
10,4 |
91,1 |
||||||
150 |
3,4 |
29,7 |
|
|
|
|
|
|
11,8 |
103,4 |
||||||
100 |
3,2 |
28,0 |
4,0 |
35,0 |
3,9 |
34,2 |
3,8 |
33,3 |
11,1 |
97,2 |
||||||
50 |
3,1 |
27,2 |
4,9 |
42,9 |
3,9 |
34,2 |
3,5 |
30,7 |
11,8 |
103,4 |
||||||
Рис.
1.4.3. Вклад острого и хронического и
хронического облучения в суммарную
дозу на полигонах 30-километровой зоны
ЧАЭС (1986 - 1991):
1 - с. Новошепеличи, 2 - с.
Залесье
Характерной особенностью
вегетативного периода 1992 г., как облучения
в суммарную дозу на полигонах показали
результаты анализа кернов из стволов
растений, явилось значительное корневое
поступление радионуклидов в вегетативные
органы растений, что существенно
сказалось на формировании дозовых
нагрузок за счет инкорпорированных
радионуклидов.
Данные, полученные
за первое полугодие 1992 г., свидетельствуют
о более низких по сравнению с предыдущими
годами дозах, поглощенных корневой
системой многолетних растений. Так,
экспозиционная доза на поверхности
почвы полигона с.Залесье составила 5-8
сГр, а на глубине 50 см - 0,3 - 0,4 сГр. Отчетливо
прослеживается значительное снижение
дозовых нагрузок на корневую систему
многолетних растений по сравнению с
предыдущими годами. В 1991 г., используя
методы термолюминесценции и электронного
парамагнитного резонанса (ЭПР) керамических
образцов, отобранных на полигонах
30-километровой зоны, удалось реконструировать
первичную дозу, полученную многолетними
насаждениями в первый год после аварии.
Результаты этих исследований
свидетельствуют о том, что основная,
ударная доза была получена в 1986 г. Эта
доза составляет 11,0 Гр для полигона с.
Новошепеличи и 4,5 Гр для полигона
с.Залесье. От суммарной дозы, полученной
за 6 лет после аварии, ударная доза
составляет 96 - 97 %. Эти данные существенно
изменяют представление о вкладе острого
и хронического облучения в формирование
радиобиологических реакций многолетних
растений (рис. 1.4.3).
Такое перераспределение
дозовых нагрузок, естественно, сказывается
на характере проявления различных
аномалий в росте и развитии латеральных
и терминальных почек растений.
4.4.2.
Тератогенез в зоне радиационных аномалий.
Действие острого и пролонгированного
облучения на образовательные ткани
Помимо
ростовых реакций на радионуклидный
прессинг, у растений в 30-километровой
зоне ЧАЭС возникли различного рода
аномалии в морфогенетическом процессе,
что связано не только с внешним облучением,
но и с неравномерностью распределения
радиоактивных элементов в тканях
формирующихся растений.
Получены
многолетние данные [34] по выходу
морфологических аномалий у озимой
пшеницы. В 1986 - 1987 гг. количество аномалий
превышало 40 %, а в последующие годы эта
цифра значительно уменьшилась. Спектр
выявленных аномалий довольно широк,
однако частота их встречаемости различна.
Так, в первый год после аварии у растений
озимой пшеницы, находящихся в фазе
кущения, наиболее частым проявлением
повреждений (49 %) было образование в
колосьях стерильных зерен или так
называемая череззерница. В 1987 г. количество
аномалий этого типа достигло 30 %, а в
последующем поколении уменьшилось (1,9
%). Часто встречались колосья с
дополнительными колосками ("грыжи")
и укороченные колосья. Образование
дополнительных колосков в последующих
поколениях возрастало почти в 2 раза. К
распространенным изменениям структуры
колоса можно отнести аномалии типа
скверхед.
Выброс радионуклидов из
разрушенного реактора в период, когда
многолетние древесные растения находились
в состоянии наибольшей радиочувствительности
(закладывались апексы побегов следующего
года, зачатки побегов, сформированных
в 1985 г., проходили этап гистогенеза),
привел их к кратковременному острому
тотальному облучению, перешедшему затем
в хроническое. В результате такого
воздействия на участках, где расчетные
значения доз составили за первый год
7,4 Гр, за второй - 2,54, за третий - 1,55 Гр,
регистрировали появление различного
рода аномалий в формообразовательном
процессе многолетних растений (рис.
1.4.4), Форма проявления аномалий зависела
от дозовых нагрузок, которые определялись
характером распреде ления радионуклидов
по ассимиляционной поверхности
вегетативных органов растений.
Под
воздействием острого облучения в
весенний период 1986 г. (полигон в
10-километровой зоне) в этом же году
произошли изменения листовых пластин
у хвойных и лиственных деревьев. Листовые
пластины ели иногда увеличивались в 3
раза. Резко изменился цвет хвоинок. У
сосны они были светло-желтые, а у ели -
малиновые. Листовые пластинки взрослых
деревьев дуба заметно утратили присущую
им конфигурацию, вплоть до приобретения
формы овала. В связи с большим накоплением
антоцианов лист изменил окраску - стал
малиновым. У однолетних саженцев дуба
(Quercus robur L.) наблюдалось сильное кущение
прикорневых побегов, укорочение черешков,
образование пальмовидных побегов на
одной ветви. Однолетние саженцы каштана
настоящего (Castanea sativa L.) представляли
собой растения с резко укороченными
побегами, небольшими листовыми
пластинками, имевшими зелено-желтый
цвет. Отмечено рассечение листовых
пластин, увеличение количества радиальных
прожилок и уменьшение между ними
мезофильных слоев клеток.
 Рис.
1.4.4. Нарушения в процессе морфогенеза
у вегетативных побегов ели, произрастающей
на полигоне с уровнем радионуклидного
загрязнения свыше 150 Ки/км-
Свето-микроскопические
исследования побегов, на которых выявлены
нарушения формообразовательных
процессов, показали [24], что развитие
латеральных почек в этом случае
заторможено, а организация терминального
апекса нарушена. Наблюдалась элиминация
зоны центральных материнских клеток и
переходной зоны; клетки периферической
зоны формировали значительную часть
почки; закладка кроющих чешуи
модифицировалась. Верхушки побегов с
укороченными стеблями и хвоей не имели
терминальной почки - ее замещали несколько
латеральных.
При сканировании
находящихся в состоянии зимнего покоя
зачатков побегов 1987 - 1988 гг. на их
расширенном основании регистрировали
существенное изменение апикальной
части, утолщение купола зачатка, на ее
месте в зачатке образовывалось углубление,
размеры которого варьировали (рис.
1.4.5).
Таким образом, при остром и
хроническом облучении у многолетних
растений, произрастающих в насаждениях
вокруг ЧАЭС, зона Центральных материнских
клеток с низким уровнем митотической
активности оказывалась более
радиочувствительной, чем инициальный
слой и периферическая зона, где уровень
митотический активности выше, что ведет
к тератогенезу. Вид и размеры формирующихся
терат зависят, по-видимому, от поглощенной
дозы.
Рис.
1.4.4. Нарушения в процессе морфогенеза
у вегетативных побегов ели, произрастающей
на полигоне с уровнем радионуклидного
загрязнения свыше 150 Ки/км-
Свето-микроскопические
исследования побегов, на которых выявлены
нарушения формообразовательных
процессов, показали [24], что развитие
латеральных почек в этом случае
заторможено, а организация терминального
апекса нарушена. Наблюдалась элиминация
зоны центральных материнских клеток и
переходной зоны; клетки периферической
зоны формировали значительную часть
почки; закладка кроющих чешуи
модифицировалась. Верхушки побегов с
укороченными стеблями и хвоей не имели
терминальной почки - ее замещали несколько
латеральных.
При сканировании
находящихся в состоянии зимнего покоя
зачатков побегов 1987 - 1988 гг. на их
расширенном основании регистрировали
существенное изменение апикальной
части, утолщение купола зачатка, на ее
месте в зачатке образовывалось углубление,
размеры которого варьировали (рис.
1.4.5).
Таким образом, при остром и
хроническом облучении у многолетних
растений, произрастающих в насаждениях
вокруг ЧАЭС, зона Центральных материнских
клеток с низким уровнем митотической
активности оказывалась более
радиочувствительной, чем инициальный
слой и периферическая зона, где уровень
митотический активности выше, что ведет
к тератогенезу. Вид и размеры формирующихся
терат зависят, по-видимому, от поглощенной
дозы.
 Рис
I.4.5. Находящийся в состоянии зимнего
покоя зачаток побега с укрупненными
листовыми зачатками и подавленным
апексом
Рис
I.4.5. Находящийся в состоянии зимнего
покоя зачаток побега с укрупненными
листовыми зачатками и подавленным
апексом
4.4.3. Частотный анализ морфогенетических изменений у растений, произрастающих в зоне повышенных уровней хронического облучения Поскольку в контрольной группе растений многие аномалии практически полностью отсутствовали, а на опытных полигонах частота отклонений от нормы в первые 2 года после аварии колебалась от 20 до 26 %, то с полным основанием выявленные тераты могут быть отнесены к индуцированным радиацией, так как при облучении растений на гамма-поле наблюдали образование аналогичных радиоморфозов. Дозовый эквивалент для таких случаев предположительно может соответствовать дозам более 15 Гр. Большинство морфологических изменений (морфозов) связано с изменениями в меристематических тканях, представляющих собой группу растительных клеток в стадии активного деления и роста. В упрощенной форме эта ткань состоит из двух типов клеток: с высокой степенью репродукции; с различной степенью дифференциации. Результаты изучения репродуктивной способности сосны при разных уровнях облучения показали, что в собранных семенах урожая 1986 - 1988 гг. такие признаки, как пустозерность, жизнеспособность, масса семян, энергия прорастания, колеблются в широком интервале в зависимости от дозы облучения [117]. Собранные семена сосны высевали в тепличных условиях для изучения морфологических признаков и их наследуемости в потомстве. Анализ однолетних сеянцев и двухлетних саженцев показал высокую изменчивость по широкому спектру морфологических признаков, большинство из которых наблюдали на взрослых деревьях. Полученные данные свидетельствуют о том, что по мере увеличения продолжительности пребывания многолетних растений в зоне с радиационной аномалией, следует ожидать селективного увеличения полиморфизма и физиологической нестабильности. В этом случае пролонгированное радиационное воздействие на меристемы точек роста может способствовать преобразованию генетического и физиологического гомеостаза, что, по-видимому, может привести к микроэволюционным сдвигам. 4.4.4. Восстановительные процессы у многолетних растений, произрастающих в зоне радиационной аномалии Хронический режим облучения, сложившийся на территории 30-километровой зоны ЧАЭС после острого периода облучения весной 1986 г., затрудняет понимание природы лучевого поражения многолетних растительных организмов, так как радиочувствительность всех их компонентов претерпевает существенные изменения в течение годового цикла развития. Интерпретация результатов осложняется и тем обстоятельством, что в ходе облучения наряду с развитием эффектов поражения протекают восстановительные процессы. После острого периода облучения восстановительные реакции у многолетних древесных и кустарниковых пород наблюдали спустя 1-1,5 года. В качестве критериев восстановления использовали интенсивность таких процессов, как корне-, побего- и порослеобразования, физиологические реакции компенсаторного характера. Многолетние деревья сосны не способны к образованию поросли. У них мощным восстановительным потенциалом вегетативного характера обладает крона, имеющая большое количество спящих почек, защищенных круглогодично фотосинтезирующей хвоей [61,117]. Наблюдения в пострадиационный период (1986 - 1992) показали, что у сосны при дозе облучения свыше 14 Гр (сублетальная доза для сосны при остром весеннем облучении) восстановление происходило за счет дополнительного побегообразования из ранее спящих почек. Этот процесс начинался спустя год после облучения, т.е. в 1987 г., и выражался в появлении в верхней части кроны единичных побегов, представляющих собой пучки очень длинной хвои, сидящей на коротком толстом ауксибласте. Роль этих побегов заключается в обеспечении минимального уровня фотоассимиляции, необходимого для поддержания жизнедеятельности не пораженных облучением аттрагирующих органов и тканей дерева (корни, древесина ствола и ветки). Подтверждением этого является чрезвычайно высокая интенсивность фотосинтеза хвои таких побегов (в 3 - 4 раза выше контрольных), что свидетельствует о компенсаторном характере этого процесса. С каждым последующим годом количество таких побегов в кроне увеличивается, что указывает о возможности постепеного частичного восстановления исходных функций многолетнего растения. Начиная с вегетационного сезона 1988 г., в зоне сильного и среднего поражения при одних и тех же поглощенных дозах интенсивнее проходило восстановление деревьев более старшего возраста, а прирост побеговпоследнего года был выше в верхних мутовках, чем в нижних. Выживаемость сильно пораженных деревьев зависит от прироста побегов последнего года, и и если прирост составляет около 20% нормальной охвоенности при наблюдаемых темпах роста, то для восстановления таких деревьев потребуется 2-3 года. поэтомувозможно выживание отдельных деревьев, у которых количество побегов с зеленой хвоей в настоящее время составляет около 5-10%. Смена вершины дерева - пострадиационная реакция на гибель осевого верхушечного побега; его роль берет на себя один из боковых побегов мутовки, наиболее мощный. Это явление отмечали примерно у 30% сосен (Pinus sylvestris) и елей (Picea excelsa L.), облученных в пределах 11-15 Гр. О возможности восстановления многолетних растений, произростающих в 30-километровой зоне ЧАЭС, свидетельствуюти данные о динамике компенсаторной и репродуктивной генерации при различных дозовых нагрузках. В качестве критериев компенсаторной регенерации исползовали также такие показатели, как заживление ран на ветвях многолетних растений, спилы ветвей определенного диамета, величина вторичного прироста на многолетних растениях после удаления верхшечной почкиюВ качестве критерия протекания репродуктивной регенерации у растений, произрастающих на почвах с различным уровнем загрязнения радионуклидами, использовали такой показатель, как интенсивность ризогенеза у черенков смородины черной (Ribes nigrum), ивы остролисной (Salix acutifolia). Полученные результаты свидетельствуют о том, что при тех ууровнях дозового прессинга, которые наблюдаются на территории 30-километровой зоны ЧАЭС, регистрируется достоверное увеличение интенсивности всех типов регенерационных процессов у многлетних растений. 4.4.5. Мутагенное действие Под влиянием внешнего и внутреннего облучения радионуклидами, выброшенными при аварии, в растениях регистрировали мутации как в соматических, так ив половых клетках. Возникновение мутаций связывается, как правило, с прямым физическим разрушением либо функциональной инактивацией под влиянием продуктов радиолиза участков хромосом и их уникальных структур. О повреждении уникальных структур может свидетельствовать увеличение рахрывов в молекулах ДНК. Используя прием дополнительного гамма-облучения растений, уже получивших определенную дозу радиации, с помощью метода электрофореза однонитевой ДНК и щелочного расплетания, обнаружили в обследованных в 1987г. популяциях травянистых растений, произростающих в окрестностях ЧАЭС, в частости, у радиочувствительного бобового растения горшка мышиного (Vicia cracca L.) повышенный выход однонитевых разрывов ДНК [136]. Формирование генетических изменений и их фиксация в последующих поколениях во многом зависят от функционирования систем репарации ДНК. При этом у растений данные системы играют особо важную роль в пыльце, что обусловлено ее особой функцией и гаплоидностью ядер. Эффективность функционирования систем репарации ДНК должна обеспечить стабильность генома, который может легко повреждаться вследствие слабой защищенности созревшего пыльцевого зерна. поэтому ослабление функционирования систем репарации является свидетельством усиления генетического действия излучения. При излучении спонтанного обновления ДНК и индуцированного острым гамма-облучением внепланового синтеза ДНК, отражающего ее репарацию, было установлено, что формирование пыльцы березы бородавчатой (Betula pendula L.) при высоких концентрациях радионуклидов в почве (7,2·1013 по сравнению с 5,6·109 Бк/км2 в конроле) приводит к подавлению системы темновой репарации ДНК в зрелой хранящейся пыльце [22]. При сохранении у такой пыльцы способности к оплодотворению можно ожидать у растений накопление генетических повреждений. Многими исследователями отмечено увеличение аберраций хромосом в образовательных тканях растений, произростающих в зоне аварии ЧАЭС. Так, обнаружено [40] увеличение в несколько раз числа клеток с аберрациями хромосом в корневых мерисистемах проростков ослинника двухлетнего (Oenothera biennis L.), выращенных в лабораторных условиях из семян растений, произрастающих при уровнях гамма-фона в 1986г. от 0,05 до 60мР/ч. При этом показана характерная прямо пропорциональная зависимость выхода мутаций от мощности дозы и обнаружены множественные аберрации хромосом, которые могут быть следствием действия плотноионизирущего альфа-излучения. При проведении исследований [47] на различных по уровню загрязнения радиоактивными выпадениями участках (от 120 до 800 МБк/м2), наблюдали линейную или близкую к ней зависимость между поглощенной в течение 26.04 - 5.06 (сбор семян) 1986г. суммарной бета- и гамма-дозой растениями озимой посевной ржи (Secale cereale L.) и выходом клеток с аберрациями хромосом в корневых меристемах проростков М2-поколениями (рис. I.4.6). Этот же тест был использован [122] для оценки влияния радиоактивного загрязнения на семена 7 видов растений, собранных в зоне, прилегающей к ЧАЭС. Мощность дозы составила 2 - 3 мР/ч. При этом у отдельных различающихся по радиочувствительности видов было обнаружено от 1 до 8% аберраций хромосом, причем наблюдалось повышение частоты аберраций с увеличением мощности дозы. Но линейной зависимости не установлено, что, по-видимому, обусловлено неравномерным распределением радионуклидов по поверхности почвы или влиянием различных загрязнителей химической природы. Интересно, что на второй год после аварии частота аберраций хромосом у некоторых видов возросла в 1,5 - 3 раза несмотря на 2 - 3-кратное снижение радиационного фона. Этот факт имеет исключительно важное значение с точки зрения гентической опасности не только внешнего, но и внутреннего облучения за счет радионуклидов, поступающих в растения из почвы. При выращивании высокорадиочувствительных конских бобов (Vicia faba L.) сорта Русские черные [36] в 30-километровой зоне при уровнях радиоактивного загрязнения пахотного горизонта почвы по 90Sr около 2·1012 Бк/км2 и по137Cs около 4·1012 Бк/км2 (поглощенная доза для корневой системы за 3 мес выращивания растений сотавляла 0,15 - 0,4 Гр) регистрировали увеличение в 2 - 3 раза количества клеток с аберрациями хромосом.
Рис. 1.4.6. Зависимость количества клеток с аберрациями хромосом (n, %) в корневой системе 24-(1), 48- (2) и 72-часовых (3) проростков семян М2-поколения озимой ржи (Secale cereale L.) от дозы хронического облучения растений М1-поколения
Кроме того, было установлено [126] увеличение количества клеток с аберрациями хромосом и хроматид в корневой меристеме проростков скерды кровельной (Crepis tectorum L.), выращенных из семян растений, собранных на территории зоны ЧАЭС при условиях гамма-фона 5-10 мР/ч в первый и 0,02 - 20 мР/ч во второй годы после аварии. В проростках первого года максимальная частота клеток с аберрациями достигала 10,2 - 15,3 %, причем нередко наблюдались клетки со множественными аберрациями. Во второй год на участках с максимальным уровнем фона ьыло обнаружено 1,4 - 2,2% клеток с аберрациями хромосом. При минимальных мощностях облучения частота клеток с аберрациями составила всего 0,3 - 0,5%; эти значения соответствуют спонтанному уровню. Кроме того, отмечали также появление растений с измененным кариотипом, что свидетельствует об активных микроэволюционных процессах в хронически облучаемых популяциях. Имеются данные [88], свидетельствующие о повышении количества стерильной пыльцы у растений, растущих на территориях, загрязненных радионуклидами. Так, исследуя две популяции фиалки утренней (Viola matutina klok), произрастающей на двух участках 30-километровой зоны ЧАЭС, различающихся на 3 порядка по величинам поглощенных доз, выявили в 1987 и 1988 гг. увеличение в 1,5 - 2 раза количества стерильных пыльцевых зерен. При уровне мощности дозы на высоте 2 м 4 - 5 Гр/год наблюдали частичную женскую стерильность сосны (Pinus sylvestris L.), выражавшуюся в снижении гамето-фитной выживаемости семяпочек, опыленных в 1986 г. и уменьшении эмбриональной выживаемости семяпочек, опыленных в 1985 г. Безусловно, стерильность - неспособность организма образовывать гаметы или достаточное их количество, не приводит к генетическому повреждению, а лишь снижает число потомков. Но при действии ионизирующей радиации стерильность, как правило, является следствием генных или хромосомных мутаций, обусловливающих нарушение мейоза - основного звена гаметогенеза. Прямым свидетельством генетического действия ионизирующих излучений на пыльцу является непосредственная регистрация в них мутаций. У растений, выращиваемых на почвах 30-километровой зоны ЧАЭС с общей бета-активностью 2,6·103- 6,3·104Бк/кг, в пыльце ячменя (Hordeum vulgare L.) отмечали увеличение частоты waxy-изменений в 2 - 3 раза [34]. Таблица І.4.7 Количество хлорофилльных мутаций типа альбина у ржи (Secale cereale L.) и ячменя (Hardeum vulgare L.), %
Культура |
Контроль |
1986 г. |
1987 г. |
1988 г. |
1989 г. |
Рожь Киевская 80 |
0,01 |
0,14 |
0,40 |
0,91 |
0,71 |
Рожь Харьковская 03 |
0,02 |
0,80 |
0,99 |
1,20 |
1,14 |
Ячмень |
0,40 |
0,90 |
0,74 |
0,80 |
0,91 |
Примечание. Семена получены в 30-километровой зоне ЧАЭС . В первые месяцы после аварии при выращивании растений ржи (Secale cereale L.) и ячменя (Hardeum sativum jessen) в условиях закрытого грунта на почвах, привезенных из 30-километровой зоны, было обнаружено увеличение частоты встречаемости различных типов хлорофилльных мутаций. Преобладали нежизнеспособные мутации типа альбина. В последующие годы семена, собранные в зоне, высевали вновь. У ржи и ячменя, выращиваемых в 30-километровой зоне на почве с суммарной гамма-активностью 1·105Бк/кг, регистрировали многократное превышение спонтанного уровня выхода хлорофилльных мутаций (табл. 1.4.7). При этом было отмечено, что в каждом последующем поколении мутации не элиминировались, поскольку высеянные семена подвергались последующему прессингу радионуклидов. Выход морфологических аномалий у 4 сортов озимой пшеницы (Triticum vulgare L.) при их выращивании в течение 3 лет на почве с суммарной активностью гамма-излучения 5,2·104- 1,8·105Бк/кг в 1986 - 1987 гг. превысил 40 %, но в 1988 г. уменьшился. Спектр выявленных аномалий довольно широк: в первый год после аварии наиболее частым (до 49 %) было образование в колосьях стерильных зон - череззерница; в 1987 г. количество аномалий этого типа достигало 30 %, а в следующем поколении снижалось до 1,9 %. Часто встречались колосья с дополнительными колосками ("грыжи") и укороченные колосья. К распространенным изменениям структуры колоса следует также отнести аномалии типа скверхед. Встречались также растения с повышенной остистостью, неравномерной остистостью, колос "елка", изменения окраски стебля и другие. У разных сортов выход морфологических аномалий проявляется по-разному, что может быть связано, с одной стороны, с различной их радиочувствительностью, а с другой - с неодинаковой способностью к накоплению радионуклидов. Безусловно, отмечаемые морфологические аномалии, отклонения от нормы, как правило, имеют место лишь в поколениях облучаемых организмов и обычно не закрепляются в потомстве. Тем не менее их стойкое проявление во все последующие годы, вплоть до 1994 г., позволяет предполагать, что многие из этих изменений носят генетический характер и свидетельствуют о том, что под влиянием ионизирующих излучений радионуклидов, выброшенных при аварии на ЧАЭС, в популяциях растений идет мутационный процесс. 4.4.6. Действие на растения инкорпорированных радионуклидов Радиочувствительность растений к облучению, создаваемому в результате накопления в тканях радионуклидов, изучена меньше, чем радиочувствительность к облучению от внешних источников. Главным образом это обусловлено тем, что большинство видов растений обладает более высокой по сравнению с млекопитающими радиоустойчивостью, и даже при накоплении радионуклидов в значительных количествах действие излучений у них проявляется гораздо в меньшей мере. Растениям обычно отводится роль переносчиков радионуклидов по пищевой цепочке "почва - растение - животное - человек" или "почва - растение - человек". Тем не менее при поступлении и накоплении радионуклидов в относительно радиочувствительных видах растений в зависимости от физико-химических характеристик радиоактивных веществ, их количества, мест локализации могут наблюдаться те же радиобиологические эффекты, что и при эквивалентных поглощенных дозах внешнего облучения: радиационная стимуляция, морфологические изменения, лучевая болезнь, ускорение старения и сокращение продолжительности жизни, гибель, генетическое действие. Однако поступившие внутрь растений радиоактивные вещества могут обладать повышенной по сравнению с внешним облучением опасностью, вызванной рядом причин. Первая и основная из них - способность некоторых радиоактивных веществ избирательно накапливаться в отдельных тканях и органах, что приводит к относительно высоким локальным уровням облучения. Вторая причина - увеличение опасности действия альфа-и бета-излучателей, которые мало влияют на растения в условиях внешнего облучения, но могут стать необычайно сильным источником ионизирующей радиации при поступлении внутрь клеток и клеточных структур. Третьей причиной являются, как правило, длительные сроки облучения. Ввиду специфики метаболизма растений радионуклиды, кумулируясь в тканях, практически не выводятся из них в отличие от животных. При огромных периодах полураспада90Sr,137Cs,239Pu даже у многолетних растений эти радионуклиды будут обусловливать облучение в течение всего онтогенеза. И если в первый год аварии облучение растений было обусловлено в значительной степени за счет внешнего облучения, то в последующие годы с начавшейся миграцией радионуклидов по трофической цепочке, оно почти исключительно обусловливается внутренним облучением. Но если у одно-двухлетних растений за сравнительно короткий период вегетации не всегда успевают сформироваться достаточно высокие дозы облучения, то у многолетних растений в течение десятков лет могут сложиться сравнительно высокие уровни облучения. Более того, подавляющее количество видов многолетних растений, как правило, чувствительнее к действию радиации, чем одно-двухлетние травянистые виды. В зоне Чернобыльской катастрофы в кроне древесных растений первоначально было задержано от 60 до 90 % выпавших на лес радионуклидов. После выпадения радиоактивных осадков на лесную растительность начинается их вертикальная миграция под влиянием сил гравитации, атмосферных осадков, движения воздуха, с листовым опалом. в результате чего радионуклиды перемещаются в нижние слои крон, под полог леса. Скорость вертикальной миграции зависит от физико-химических характеристик радиоактивных выпадений, типа и возраста древостоя, метрологических условий, времени года. Спустя некоторый период, который в хвойных лесах может измеряться несколькими годами, основная масса радиоактивных веществ переходит в лесную подстилку и в верхний слой почвы. Именно из него через 4 - 5 лет в лиственном лесу и через 8 - 10 лет в хвойном (что обусловлено особенностями в скорости вертикальной миграции радионуклидов и более быстрой минерализацией листьев по сравнению с хвоей) начинается активное поступление радионуклидов в древесные растения через корни. Если механизмы поступления и усвоения радионуклидов однодвухлетними травянистыми и многолетними древесными растениями в основном не различаются, то характер из накопления имеет принципиальные различия. Многолетние древесные способны к существенному накоплению радионуклидов во всех органах - листьях, ветках, коре, древесине. Многолетний замкнутый цикл радионуклидов по цепочке "листья - лесная подстилка - почва - корни - ствол - листья" и так далее может приводить к существенному радиоактивному загрязнению всех органов. У одного из самых радиочувствительных видов - сосны - это может приводить к самым различным типам нарушений. Повреждаться могут и другие виды. Ель, например, обладает более высокой радиоустойчивостью, но она является кальциефилом и способна накапливать -Sr в больших количествах, чем сосна и другие виды. Различные радионуклиды не только по-разному накапливаются отдельными видами растений, но и по-разному концентрируются в различных органах, приводя к различной степени облучения. На радиоавтографах целых растений, полученных при введении различных радионуклидов, как правило, бывает хорошо определена неравномерность их распределения по отдельным органам. Четко прослеживаются места концентрации, сосредоточенные, как правило, в тканях, которые обладают высокой метаболической активностью (меристема и генеративные органы) и наиболее высокой радиочувствительностью. Именно поэтому как при внешнем, так и при внутреннем облучении радиобиологические эффекты, в первую очередь, проявляются на этих группах клеток. В частности, в меристемах наблюдаются замедление или ускорение деления клеток, формирование различного рода морфологических изменений, замедление роста органов и растения в целом, гибель меристем и др. В год аварии в пяти областях Украины (Киевской, Черниговской, Житомирской, Черкасской и Винницкой), в том числе в различных местах 30-километровой зоны, были собраны образцы семян 9 сортов озимой пшеницы (Tviticum vulgare L.), растения которой в течение 3 мес - с фазы кущения (время аварии) до созревания - подвергались облучению радиоактивными выбросами. Максимальный уровень суммарной бета- и гамма-активности семян достигал 3,7·104Бк/кг. По общепринятым методикам экспериментального мутагенеза в последующих б поколениях изучались частоты и спектр мутационной изменчивости растений. У 18,4 % образцов выявили достоверное увеличение (более чем в 2,7 раза) частоты хромосомных аберраций в клетках меристемы первичного корешка прорастающих семян. В одном случае у проростков из семян растений, выросших в 16 км от станции, наблюдали увеличение в 32 раза количества аберраций. Регистрировали увеличение в 2,7 - 8,7 раза частоты видимых мутаций. Их спектр охватывал 27 типов мутаций по признакам структуры стебля, листа и колоса, физиологическим признакам. Спецификой спектра были сравнительно частые мутации по генам карликовости, а также редко встречающиеся в обычных условиях мутации типа "ощипанный колос" и "разветвленный колос". Мутации полимерных генов по элементам структуры урожая в последующих поколениях привели к изменению продуктивности мутантов и в итоге к потере типичности сортов. При выращивании проростков гороха (Pisum sativum L.) и кукурузы (Zea mays L.) на водных вытяжках почв из 10-километровой зоны ЧАЭС, имеющих суммарную бета-активность 5·10-9- 5·10-8Ки/л, наблюдали [46] заметное снижение относительной скорости роста растений. Радиационную стимуляцию, выражавшуюся в ускорении прорастания семян, роста растений, прохождении фаз развития, повышении фотохимической активности хлоропластов, увеличении содержания хлорофилла, наблюдали у желтого люпина (Lupinus luteus L.), выращенного из семян, которые были собраны в 30-километровой зоне ЧАЭС при уровне загрязнения по137Cs Ки/км2. Такие семена содержали137Cs 2,31·103и90Sr 1,65·102Бк/кг [115]. Наибольшую чувствительность к радиоактивному загрязнению проявили сосновые леса по причине высокой радиочувтвительности этого вида растений, значительной задерживающей способности хвои и, как следствие, высокого вклада бета-излучения в дозовую нагрузку [49]. Общая поглощенная доза в хвое и почках примерно в 10 раз больше дозы внешнего гамма-облучения. Именно поэтому на участках с высоким уровнем загрязнения в активно растущих меристемах обнаруживали наиболее сильные повреждения, приводящие к их массовой гибели, пролифирации боковых почек с неупорядоченной ориентацией, образованию укороченных побегов с мелкой или гигантской хвоей, искривлению иголок хвои, нарушению ориентации молодых вегетативных побегов. К концу лета 1986 г. проявились летальные эффекты облучения сосны. В течение последующего осенне-зимнего периода было выявлено, что общая площадь погибшего лесного массива, примыкающего с запада к промплощадке АЭС, достигает 500 га. Спустя 4 года после аварии наблюдалось снижение в 5 - 10 раз продуктивности семян у аборигенной популяции подорожника ланце-толистного (Plantago lanceolata L.), произрастающей в местах с уровнем гамма-фона от 2,8 до 1334 пА/кг [120]. По-видимому, данный тип радиационного поражения растений можно связать с угнетающим действием многолетнего хронического облучения на популяцию этого радиочувствительного представителя травяного покрова, не выделяя вклад внешнего и внутреннего облучения. Кроме того, без сомнения, можно утверждать, что на 4-й год после аварии, когда внешнее облучение по сравнению с первоначальным поставарийным периодом уменьшилось во много раз, фактор внутреннего облучения в радиационном повреждении растений играет определяющее значение. Особенно это касается травянистых растений с мочковатой корневой системой, формирующих основную массу корней в верхнем, наиболее загрязненном радионуклидами слое почвы. Тем не менее следует признать, что главная опасность накопления растениями радионуклидов состоит не в их традиционной угрозе для растений, а в том, что они являются главным звеном миграционной цепочки на пути передачи радионуклидов более радиочувствительным видам живых организмов - млекопитающим и, в первую очередь, человеку. 4.5. Влияние аварии на биоценоз 4.5.1. Действующие факторы Чтобы правильно понять причины изменений, наблюдавшихся в популяциях растений и животных после Чернобыльской катастрофы, надо принимать во внимание не только действие радиации, но и многие другие факторы, прямо или косвенно связанные с аварией на ЧАЭС или не имеющие с ней никакой причинной связи. 1. Радиационное поражение растений и животных. 2. Нарушение трофических цепей, освобождение экологических ниш в результате гибели или снижения численности видов-прокормителей или видов-конкурентов. 3. Радиационная стимуляция (гормезис). 4. Химическое загрязнение среды нерадиоактивными веществами, применявшимися для ликвидации последствий аварии на ЧАЭС. 5. Сплошная инженерная дезактивация особо загрязненных лесных кварталов, лугов, населенных пунктов (захоронение, засыпка песком). 6. Подтопление и образование мелководных водоемов после перекрытия мелиоративной сети дамбами и последующий спуск этих водоемов. 7. Подъем минеральной взвеси в р.Припять в результате защитных гидротехнических работ в русле и на берегах, массовое использование комплексообразователей в защитных гидротехнических сооружениях. 8. Искусственный разгон облаков над зоной отчуждения, увеличение уровня солнечной радиации. 9. Эвакуация населения, крупного рогатого скота, отстрел домашних хищников. Ю. Прекращение обработки пахотных земель, сбора урожая, выпаса и сенокошения, ухода за садами, применения удобрений и пестицидов. 11. Прекращение промышленных рубок, рубок ухода, расчистки сухостоя, расчистки просек в лесах зоны отчуждения. 12. Снятие рекреационной нагрузки на природные угодья, прекращение промышленного рыбоводства на пруде-охладителе ЧАЭС, промыслового и спортивного рыболовства и охоты. 13. Продолжающаяся сукцессия растительного и животного населения Киевского водохранилища после его заполнения. 14. Климатические особенности послеаварийных лет: теплая весна 1986 г., многоснежная и холодная зима 1986 - 1987 гг., мягкие зимы последующих лет, низкие уровни весенних паводков, дождливое лето 1988 и 1991 гг., засушливые весна и лето 1992 г. 15. 11-летние циклы солнечной активности, с которыми синхрони-зуется динамика численности многих видов растений и животных. Эти и другие менее значительные факторы порождают сложные переплетающиеся цепочки событий в живой природе, которые выражаются в смене доминирующих видов, перераспределении и изменении численности видов растений, животных и микроорганизмов. Они* затрагивают все главные звенья биоценоза: первичных продуцентов биомассы, консументов разных уровней - растительноядных животных и хищников - и редуцентов, возвращающих органогенные элементы из отмершей биомассы снова в круговорот веществ. Действие радиации непосредственно проявлялось только на ограниченной территории ближней зоны вокруг станции (радиусом 5-10 км), на остальной территории зоны отчуждения (свыше 99 %) ход событий в природных угодьях определяется в основном вторичными экологическими последствиями катастрофы. 4.5.2. Радиационное поражение биоценозов Гибель хвойных лесов.Высокая радиочувствительность хвойных растений, усиленная тем обстоятельством, что авария произошла в период распускания почек, была причиной поражения насаждений сосны обыкновенной (Finns silvestris L.) и разрушения экосистемы сосновых лесов на наиболее загрязненных территориях вблизи атомной станции [61]. Особую роль в поражении фотосинтезирующих органов и ростовых тканей сыграло бета-излучение от осевших на поверхность растений радионуклидов; вклад внешнего гамма-облучения в поглощенную дозу составил всего 10 % [114]. На основании установленной зависимости степени поражения сосны от мощности экспозиционной дозы, рассчитанных поглощенных доз и вызванных ими эффектов поражения была составлена карта (рис. 1.4.8) радиационного поражения сосновых лесов [63]. Зона летального поражения характеризуется полной гибелью сосны. Она ограничена изолинией мощности экспозиционной дозы (МЭД) по гамма-излучению 0,5 Р/ч на 01.06.86. Расчетная поглощенная доза по гамма-излучению в зоне летального поражения составила 80 - 100 Гр. Общая площадь погибшего леса 580 га. Хвоя погибших сосен приобрела яркую коричнево-бурую окраску (так называемый рыжий лес). "Рыжий лес" простирается вдоль основных следов радиоактивного выброса на 5 - 7 км на север, 2 - 3 км на запад, 1,5 - 2 км на юг. Отдельные пятна "рыжего леса" встречаются на расстоянии до 9 км к северу и 6 км к западу от ЧАЭС . Лиственные леса и кустарники, травяные ценозы и мохово-лишайниковый покров в зоне летального поражения испытали усыхание вершин, угнетение ростовых процессов и повреждение репродуктивных органов в течение первого сезона вегетации. В зоне сублетального поражения (расчетная поглощенная доза 10 - 20 Гр при максимальной МЭД 0,1 - 0,5Р/ч) произошла полная гибель молодых экземпляров сосны при частичном омертвении молодых побегов взрослых деревьев. Наряду с молодняками наиболее пострадали ослабленные насаждения сосняков-беломошников на бедных песчаных почвах. Площадь сосновых лесов в этой зоне - 3750 га.
Рис.1.4.7. Радиационное поражение сосновых лесов в ближней зоне ЧАЭС. 1 -леса; 2 - погибший "рыжий лес"; 3 - погибший "рыжий лес", раскорчеванный со сплошным снятием грунта; 4 -луга; 5 - залежи; 6 - участки сплошного снятия грунта; 7 - застроенные территории; зоны поражения сосновых лесов: 8 летального (более 500 мР/ч);9 -сублетального (50 - 500 мР/ч); среднего (20 - 50 мР/ч)
Прирост сосны в зоне сублетального поражения, приостановившийся в 1986 г., в следующем году возобновился. В зоне среднего поражения, которая ограничена изолинией МЭД 50 мР/ч на 01.06.86, расчетная поглощенная доза оценена в 3 - 5 Гр. Зона характеризуется угнетением роста и гибелью точек роста хвойных пород летом 1986 г. и образованием радиоморфозов, особенно сильно выразившихся в 1987 г. В этой зоне оказалось 11920 га сосновых лесов. Оставшаяся часть 30-километровой зоны и ряд радиоактивных пятен за ее пределами могут быть отнесены к зоне слабого радиационного поражения. Внешние признаки морфологических нарушений у хвойных здесь отсутствовали, но в 1986 - 1987 гг. отмечалось снижение всхожести семян и хромосомные аномалии в мейозе. Радиационное поражение лесных биоценозов в зоне Чернобыльской катастрофы не приобрело массового характера и коснулось преимущественно сосновых лесов. Гибель сосновых насаждений охватила менее 0,5 % площади лесов зоны, не оказала существенного влияниянарадиоэкологическую обстановку в целом, хотя и ухудшила ее в непосредственной близости от атомной станции. Гибель животных. Радиочувствительность позвоночных животных выше, чем радиочувствительность хвойных растений. Однако документальных свидетельств о массовой гибели позвоночных в ближней зоне в острый послеаварийный период нет - первые биологические наблюдения в зоне отчуждения были начаты лишь в июне-июле 1986 г. По данным количественных учетов мышевидных грызунов в сентябре 1986 г., в ближней зоне численность этих животных снизилась в 3 - 5 раз (3,5 % ловушко-суток вместо 15 - 20 в контрольных, слабо загрязненных участках). Расчетная поглощенная доза в первый месяц после аварии составляла на учетных площадках 22 и 860 Гр для гамма- и бета-облучения соответственно [112]. Беспозвоночные на один-два порядка более устойчивы к действию ионизирующей радиации по сравнению с позвоночными. Тем не менее три экологические группы беспозвоночных сильно пострадали из-за специфических особенностей облучения в их средах обитания. Скопление радионуклидов в подстилке хвойного леса буквально выжгло мелких беспозвоночных (панцирные клещи, ногохвостки) на учетных площадках в 3 км к югу от аварийного энергоблока - их численность в июле 1986 г. упала на 2 порядка. Резко снизилась также численность почвенных беспозвоночных, лучше экранированных от бета-излучения. Соотношение неполовозрелых и половозрелых особей изменилось в пользу последних. Величина поглощенной дозы 30 Гр не влияла непосредственно на взрослых животных, но заметно сказалась на ювенильных стадиях [64]. В загрязненных лишайниках-эпифитах Hypogymnia physodes (L.) Nyl. на тех же учетных участках в 3 км к югу от блока вообще не удалось обнаружить панцирных и гамазовых клещей, ногохвосток, обычных для этой экологической ниши [75]. Изменение микробного ценоза почвы.В 1986 - 1992 гг. проводился мониторинг таксономического состава и численности почвенных микромицетов в зоне отчуждения и в Вышгородском р-не Киевской обл. Из 670 образцов на дерново - подзолистых и супесчаных почвах выделено свыше 3000 культур грибов, отнесенных после идентификации к 180 видам 72 родов. В течение 1986 - 1988 гг. при общем снижении (в 2 - 3 раза) количества грибных диаспор и длины грибного мицелия в почве всех 7 пунктов наблюдения 30-километровой зоны отмечали преобладание темноокрашенного грибного мицелия на глубине 0-10 СЃРј. Аналогичную ситуацию констатировали также при построении корреляционных плеяд по методу П.В.Терентьева [111], отражающих сформировавшиеся в этом регионе грибные комплексы. Выявлены сложно организованные корреляционные плеяды - показатель стабильности грибных комплексов с преобладанием в их структуре меланинсодержащих видов и родов (рис.1.4.8). Обнаруженное явление квалифицируется как промышленный меланизм почвенных микромицетов [44]. В 1989 г. и позднее количество грибного мицелия приближалось к значениям, известным для "чистых" регионов [25]. Большей частью доминирующее положение по количеству грибной биомассы занимали светлоокрашенные микромицеты, что характерно для микобиоты этого региона до аварии. Однако в таксономической структуре грибных комплексов в 1989 - 1992 гг. доминирующее положение по-прежнему принадлежало темноокрашенным грибам. Эта группа грибов может быть биоиндикатором неблагополучной экологической ситуации, хотя непосредственная причина наблюдаемых изменений и прямая их связь с облучением не установлены.
Рис.1.4.8. Комплексы микромицетов (корреляционные плеяды) в почвах с разным уровнем радиоактивности: I -Cidiodendron;2 -Sordaria;3 -Verticillium;4 -Actmomucor;5 -Scopulanopsis; 0 - Mycelia sterilia( темный ); 7 -Drechslera;8 -Conoplea;9 -AUescheriella;10 -Arthrimum;II -Pithomyces;12 -Aureobasidium;13 -Melanophoma;14 -Phoma;15 -Rhinocladium-16 -Humicola;17 -PeyroneUeae;18 -Rhinoccphalum;19 -Stemphylium;20 -Trichothecium;21 -Mammaria;22 -Botrytis;23 -Zythia;24 -Scolecobasidium;25 -Ulocladium
4.5.3.
Вторичные явления в пораженных
биоценозах
Разрушение трофической
пирамиды.В
первые 1-3 года после гибели сосны
разрушается вся система трофических
связей в лесном биоценозе (рис. 1.4.9). В
первую очередь лишаются пищи разнообразные
насекомые, питавшиеся на хвое и побегах:
личинки пилильщиков, гусеницы бабочек,
тли-хермесы, сосновые клопики. С их
исчезновением была подорвана кормовая
база насекомоядных птиц и муравьев,
собиравших в кронах белковый корм и
сладкие выделения тлей. Численность
почвенных микроартропод в лесной
подстилке восстановилась со спадом
радиационного фона [64], однако их видовой
состав остался обедненным, в том числе
исчезли клещи-орибатиды, промежуточные
хозяева цестод, заражающих мышевидных
грызунов [105].
В зоне среднего поражения
в 1986 - 1987 гг. нарушалась морфология
хвоинок, на 10 - 20 сут задерживался весенний
рост побегов, в 2-3 раза снизился выход
семян сосны, а пустозерность достигла
70 % при норме 5 - 18 % [61]. Это обедняло
кормовую базу фитофагов и нарушало
согласование во времени циклов развития
растений и фитофагов.
Размножение
насекомых-вредителей,В
зоне сублетального и среднего поражения
сосны возникла и сохраняется в течение
последних лет вспышка массового
размножения вторичных стволовых
вредителей.
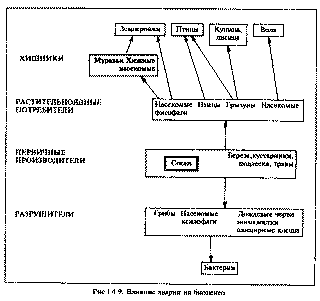 Рис.1.4.9.
Влияние аварии на биоценоз
Отмечено
11 массовых видов, из них главные - большой
и малый сосновый лубоеды (Blastophagus piniperda
L., В.minor Hart.), синяя сосновая златка
(Phaenops cyanea F.). Плотность заселения лубоедами
в Лелевском и Новошепеличском лесничествах
достигла 1,6 - 1,9 особей/дм2.
Для дополнительного питания лубоеды
внедряются в побеги. Хотя абсолютная
численность вредителей возросла лишь
вдвое по сравнению с нормой, но уменьшилось
число побегов и резко увеличился процент
заселения [I]. Опад поврежденных побегов
возрос до 70 - 75 %, против 25 - 30 % в контроле.
В 1990 г. практически все побеги в зоне
сублетального поражения были уничтожены.
Деятельность вредителей ускоряет гибель
пострадавших деревьев.
Через год
после гибели хвойные деревья заселялись
в массовом количестве обитающими под
корой жуками-трухляками Pytho depressus L. и
усачами рода Rhagium. Пик их численности
(несколько сотен особей на дерево) пришел
на 1989 г., а уже в 1991 г. очередные погибшие
деревья были слабо заражены. По-видимому,
медленно засыхавшие деревья, погибшие
в более поздние сроки, были мало
привлекательны для стволовых вредителей
из-за низкого содержания крахмала в
лубе и заболони [68]. Под действием грибов
и насекомых деревья полностью теряют
кору и через 3 - 4 года падают.
Захламленность
окружающих лесов сухостоем поддерживает
кормовую базу вредителей и создает
реальную опасность массового размножения
их в зоне среднего поражения.
Сукцессии
растительности в пораженных лесах. В
зоне летального поражения произошло
полное уничтожение лесных ценозов,
поскольку сосна погибла, а лесные
компоненты травяного покрова были
подавлены вторжением видов-эксплерентов.
В зонах сублетального и среднего
поражения лесные ценозы сохранились.
Во всех этих случаях в ценозах начались
восстановительные сукцессии, отличающиеся
по степени продвинутсти [12]. Гибель
древесного яруса и осыпание хвои вызвали
осветление леса, а также повышение
уровня грунтовых вод, увеличение
влажности почвы. На участках, где
произошла массовая гибель сосны, по
аналогии с зарастанием лесосек и гарей,
это привело на первом этапе к появлению
сорно-опушечных видов, таких, как ослинник
двулетний (Oenothera biennis L.), мелколепестник
канадский (Erigeron canadensis L.), тысячелистник
(Achillea millefolium L.), марьянник обыкновенный
(Melampymm vulgatum Pers.) и др.
При этом сохранились
виды, доминировавшие в определенных
типах сосновых боров. Например, в
зеленомошниках это черника •(Vacciniwn
myrtillis L.), вереск (Calluna vulgaris (L.) Hull), овсяница
овечья (Festuca ovina L.), орляк (Pteridium aquilinum (L.)
Kuhn), в сосняках лишайниковых - тимьян
(Thimus serpillum L.), овсяница овечья, полевица
тонкая (Agrostis tenuis Sibth.), ястребинка
волосистая (Hieracium pfflosella L.). Также
фрагментарно в виде пятен разного
размера сохраняется прежний моховой и
лишайниковый покров.
В дальнейшем
основу травяного покрова образуют,
вплоть до сплошных зарослей, типичные
растения-псаммофиты: булавоносец
седоватый (Corynephonis canescens (L.) Beauv.), иван-чай
узколистный (Chamaertion angustifolium (L.) Holub), щавель
воробьиный (Rumex acetosella L.), цмин песчаный
(Helichrysum arenamim (L.) Moench), но чаще всего и в
большом количестве - вейник наземный
(Calamagrostis epigeios (L.) Roth).
Часто в разреженном
лесу при наличии зачатков образуются
также заросли малины (Rubus idaeus L.) и ежевики
(R. caesius L., R-nessensis W.Hall). В образовавшихся
"окнах" на месте выпавших деревьев
появляются проростки деревьев. В массовом
количестве в первую очередь здесь
появляется молодняк березы поникшей
(Betula pendula Roth.), значительно реже - осины
(Populus tremula L.). Восстановлению березы
способствует активная роющая деятельность
кабанов (Sus scrofa L.). Количество проростков
сосны по сравнению с березой в тысячи
раз меньше и в первые годы жизни их
прирост очень мал. Поэтому при
восстановлении сомкнутого древостоя,
как правило, образуется березовый
(вторичный) лес, при сохранившихся соснах
- сосново-березовый. Сосновый бор
образуется только после отмирания
березового древостоя, т.е. через много
десятков или даже сотен лет.
Миграция
животных в зону поражения,С
соседних участков на территорию "рыжего
леса" постоянно проникали зайцы-русаки
(Lepus europaeus Pall.), лисицы (Vulpes vulpes L.), бродячие
собаки, залетные птицы, вероятно,
мигрировали и мышевидные грызуны. В
марте 1987 г. во время работ по захоронению
следы грызунов были обнаружены в наиболее
загрязненном участке леса, у забора
ЧАЭС [119]. В том же 1987 г. численность
мышевидных грызунов на ст. Янов и на
учетных площадках в "рыжем лесу"
восстановилась за счет миграции
[112].
Участки погибшего леса и пустоши
над захороненным лесом постоянно
посещаются крупными копытными (кабан,
лось Alces alces (L.), в меньшем количестве -
косуля Capreolus capreolus (L.)), однако места их
отстоя, лежек или основных кормежек
находятся на расстоянии 1-2 км, в зеленом
хвойном лесу, вкраплениях лиственного
леса и на заболоченных участках. Об этих
визитах свидетельствуют следы
животных.
Учет численности птиц по
голосам, проведенный в мае 1987 г. в
правобережном участке "рыжего леса"
(мощность экспозиционной дозы составляла
40 мР/ч), показал, что прилетевшие с зимовки
птицы поселились в кронах и среди мертвых
деревьев в количестве до 6 семей на
гектар (были зарегистрированы вертишейка
(Jynx torquffla L.), серая мухоловка (Muscicapa striata
(Pall.)), зяблик (Fringilla coelebs L.), полевой конек
(Anthus campestris L.), белая трясогузка Motacilla
alba L. )). Однако эта численность была в 4
- 5 раз ниже нормальной на контрольных
участках здорового соснового бора или
зеленых сосен, задержанных в росте из-за
лучевого поражения [12].
Сукцессии
растительности на площадях дезактивации.Из
площадей, подвергавшихся дезактивации,
наиболее сильно изменены участок
вырубленного и захороненного леса,
участок между ЧАЭС и Припят-ским затоном,
где была проведена отсыпка песка, и
прирусловая дамба, намытая вдоль левого
берега р.Припять, захоронение дачной
застройки у ст. Янов. В зависимости от
степени влияния разнообразных факторов
на отдельные участки дезактивированных
территорий на них можно наблюдать разные
стадии восстановительных сукцессии
растительного покрова. Ход сукцессии
рассматривается на примере "рыжего
леса".
В урочище "рыжий лес"
высокие деревья были срублены, а низкие
захоронены в траншеи, освобожденная
территория засыпана слоем песка до 0,5
м и более. После этого песчаный покров
был обработан для закрепления химическими
реактивами, образовавшими пленку.
Поскольку она оказалась неустойчивой
и препятствовала естественному
зарастанию, был проведен посев ржи с
подсевом смеси многолетних трав. Из
видов, использованных для залужения, к
1992 г. сохранилась в заметном количестве
лишь ежа сборная (Dactylis glomerataL.).
Рис.1.4.9.
Влияние аварии на биоценоз
Отмечено
11 массовых видов, из них главные - большой
и малый сосновый лубоеды (Blastophagus piniperda
L., В.minor Hart.), синяя сосновая златка
(Phaenops cyanea F.). Плотность заселения лубоедами
в Лелевском и Новошепеличском лесничествах
достигла 1,6 - 1,9 особей/дм2.
Для дополнительного питания лубоеды
внедряются в побеги. Хотя абсолютная
численность вредителей возросла лишь
вдвое по сравнению с нормой, но уменьшилось
число побегов и резко увеличился процент
заселения [I]. Опад поврежденных побегов
возрос до 70 - 75 %, против 25 - 30 % в контроле.
В 1990 г. практически все побеги в зоне
сублетального поражения были уничтожены.
Деятельность вредителей ускоряет гибель
пострадавших деревьев.
Через год
после гибели хвойные деревья заселялись
в массовом количестве обитающими под
корой жуками-трухляками Pytho depressus L. и
усачами рода Rhagium. Пик их численности
(несколько сотен особей на дерево) пришел
на 1989 г., а уже в 1991 г. очередные погибшие
деревья были слабо заражены. По-видимому,
медленно засыхавшие деревья, погибшие
в более поздние сроки, были мало
привлекательны для стволовых вредителей
из-за низкого содержания крахмала в
лубе и заболони [68]. Под действием грибов
и насекомых деревья полностью теряют
кору и через 3 - 4 года падают.
Захламленность
окружающих лесов сухостоем поддерживает
кормовую базу вредителей и создает
реальную опасность массового размножения
их в зоне среднего поражения.
Сукцессии
растительности в пораженных лесах. В
зоне летального поражения произошло
полное уничтожение лесных ценозов,
поскольку сосна погибла, а лесные
компоненты травяного покрова были
подавлены вторжением видов-эксплерентов.
В зонах сублетального и среднего
поражения лесные ценозы сохранились.
Во всех этих случаях в ценозах начались
восстановительные сукцессии, отличающиеся
по степени продвинутсти [12]. Гибель
древесного яруса и осыпание хвои вызвали
осветление леса, а также повышение
уровня грунтовых вод, увеличение
влажности почвы. На участках, где
произошла массовая гибель сосны, по
аналогии с зарастанием лесосек и гарей,
это привело на первом этапе к появлению
сорно-опушечных видов, таких, как ослинник
двулетний (Oenothera biennis L.), мелколепестник
канадский (Erigeron canadensis L.), тысячелистник
(Achillea millefolium L.), марьянник обыкновенный
(Melampymm vulgatum Pers.) и др.
При этом сохранились
виды, доминировавшие в определенных
типах сосновых боров. Например, в
зеленомошниках это черника •(Vacciniwn
myrtillis L.), вереск (Calluna vulgaris (L.) Hull), овсяница
овечья (Festuca ovina L.), орляк (Pteridium aquilinum (L.)
Kuhn), в сосняках лишайниковых - тимьян
(Thimus serpillum L.), овсяница овечья, полевица
тонкая (Agrostis tenuis Sibth.), ястребинка
волосистая (Hieracium pfflosella L.). Также
фрагментарно в виде пятен разного
размера сохраняется прежний моховой и
лишайниковый покров.
В дальнейшем
основу травяного покрова образуют,
вплоть до сплошных зарослей, типичные
растения-псаммофиты: булавоносец
седоватый (Corynephonis canescens (L.) Beauv.), иван-чай
узколистный (Chamaertion angustifolium (L.) Holub), щавель
воробьиный (Rumex acetosella L.), цмин песчаный
(Helichrysum arenamim (L.) Moench), но чаще всего и в
большом количестве - вейник наземный
(Calamagrostis epigeios (L.) Roth).
Часто в разреженном
лесу при наличии зачатков образуются
также заросли малины (Rubus idaeus L.) и ежевики
(R. caesius L., R-nessensis W.Hall). В образовавшихся
"окнах" на месте выпавших деревьев
появляются проростки деревьев. В массовом
количестве в первую очередь здесь
появляется молодняк березы поникшей
(Betula pendula Roth.), значительно реже - осины
(Populus tremula L.). Восстановлению березы
способствует активная роющая деятельность
кабанов (Sus scrofa L.). Количество проростков
сосны по сравнению с березой в тысячи
раз меньше и в первые годы жизни их
прирост очень мал. Поэтому при
восстановлении сомкнутого древостоя,
как правило, образуется березовый
(вторичный) лес, при сохранившихся соснах
- сосново-березовый. Сосновый бор
образуется только после отмирания
березового древостоя, т.е. через много
десятков или даже сотен лет.
Миграция
животных в зону поражения,С
соседних участков на территорию "рыжего
леса" постоянно проникали зайцы-русаки
(Lepus europaeus Pall.), лисицы (Vulpes vulpes L.), бродячие
собаки, залетные птицы, вероятно,
мигрировали и мышевидные грызуны. В
марте 1987 г. во время работ по захоронению
следы грызунов были обнаружены в наиболее
загрязненном участке леса, у забора
ЧАЭС [119]. В том же 1987 г. численность
мышевидных грызунов на ст. Янов и на
учетных площадках в "рыжем лесу"
восстановилась за счет миграции
[112].
Участки погибшего леса и пустоши
над захороненным лесом постоянно
посещаются крупными копытными (кабан,
лось Alces alces (L.), в меньшем количестве -
косуля Capreolus capreolus (L.)), однако места их
отстоя, лежек или основных кормежек
находятся на расстоянии 1-2 км, в зеленом
хвойном лесу, вкраплениях лиственного
леса и на заболоченных участках. Об этих
визитах свидетельствуют следы
животных.
Учет численности птиц по
голосам, проведенный в мае 1987 г. в
правобережном участке "рыжего леса"
(мощность экспозиционной дозы составляла
40 мР/ч), показал, что прилетевшие с зимовки
птицы поселились в кронах и среди мертвых
деревьев в количестве до 6 семей на
гектар (были зарегистрированы вертишейка
(Jynx torquffla L.), серая мухоловка (Muscicapa striata
(Pall.)), зяблик (Fringilla coelebs L.), полевой конек
(Anthus campestris L.), белая трясогузка Motacilla
alba L. )). Однако эта численность была в 4
- 5 раз ниже нормальной на контрольных
участках здорового соснового бора или
зеленых сосен, задержанных в росте из-за
лучевого поражения [12].
Сукцессии
растительности на площадях дезактивации.Из
площадей, подвергавшихся дезактивации,
наиболее сильно изменены участок
вырубленного и захороненного леса,
участок между ЧАЭС и Припят-ским затоном,
где была проведена отсыпка песка, и
прирусловая дамба, намытая вдоль левого
берега р.Припять, захоронение дачной
застройки у ст. Янов. В зависимости от
степени влияния разнообразных факторов
на отдельные участки дезактивированных
территорий на них можно наблюдать разные
стадии восстановительных сукцессии
растительного покрова. Ход сукцессии
рассматривается на примере "рыжего
леса".
В урочище "рыжий лес"
высокие деревья были срублены, а низкие
захоронены в траншеи, освобожденная
территория засыпана слоем песка до 0,5
м и более. После этого песчаный покров
был обработан для закрепления химическими
реактивами, образовавшими пленку.
Поскольку она оказалась неустойчивой
и препятствовала естественному
зарастанию, был проведен посев ржи с
подсевом смеси многолетних трав. Из
видов, использованных для залужения, к
1992 г. сохранилась в заметном количестве
лишь ежа сборная (Dactylis glomerataL.).
На следующий год после посева трав была проведена посадка саженцев древесно-кустарниковых пород: сосна обыкновенная, дуб северный (Quercus borealis Michx.), робиния лжеакация (Robinia pseudoacacia L.), береза повислая. Посадки прижились, но неравномерно. Кроме того, значительная часть их пострадала от личинок хрущей (Melolontha melolontha L.), а многие сосенки объедены лосями [7]. При характеристике изменений видового состава в ходе сукцессии как основной параметр использовали изменения проективного покрытия видов. Для количественного выражения степени демутации рассчитывали соотношение проективного покрытия коренной, восстанавливающейся растительности к общему проективному покрытию биоценоза [II]. Спонтанная демутация растительности на этих песчаных пустошах началась с формирования редкого травяного покрова из видов-псаммофитов. По аналогии с демутациями на залежах видами-пионерами обычно являлись сорняки-малолетники. На территории "рыжего леса" преобладает мелколепестник канадский, встречаются ослинник двулетний (Oenothera biennis L.), икотник серый (Berteroa incana (L.) DS) и др. Характерной особенностью этой территории является широкое распространение специфических карликовых мхов, часто образующих покров и закрепляющих песок до появления травянистых растений: кукушкин лен волосоносный (Polytrichum pilifenim Hedw.) цератодон пурпуный (Ceratodon purpyreus (Hedw) Brid.) , брий серебристый (Bryum argenteum Hedw.). Доминантами-ценозообразователями новообразующихся ассоциаций в этих условиях в зависимости от микрорельефа выступают вей-ник (Calamagrostis epigeios (L.) Roth), булавоносец седоватый (Corynephorus canescens (L.) Beauv.), полевицы собачья (Agrostis canina L.) и виноградниковая (A-vinealis Schreb.), мятлик узколистный (Роа angustifolis L.) и пырей ползучий (Elytrigia repens (L.) Hevski). В зависимости от условий все названные виды трав и мхов образуют сообщества с доминированием одного или нескольких видов - до 27 ассоциаций. Всего же здесь зафиксировано в 1992 г. 133 вида сосудистых растений. Несмотря на хорошую адаптированность образующихся травяных сообществ к экологическим условиям, созданным в местах дезактивации, со временем их неизбежно вытеснят лесные березовые, а позднее - сосновые сообщества, а в пойме - заросли шелюги (Salix acutifolia Willd.) и позднее - дубовый лес. Виды, создающие дугообразный покров, на протяжении определенного времени будут содоминантами растительного покрова в молодых лесах. Сукцессии растительности на площадях дезактивации протекали по обычной схеме. Так же, как и на залежах, не отмечено каких-либо отклонений, вызванных повышенным радиационным фоном. Для животного населения песчаных пустошей характерны заяц-русак, куропатка серая (Perdix perdix L.), из насекомых - жуки-скакуны (Cicindela), типичные обитатели песчаных арен; новообразованные луга регулярно посещаются дикими свиньями, лосями, косулями. После высева ржи, осенью, когда зерна стали осыпаться, поля привлекали грачей и голубей. Их деятельность положила конец надеждам на регулярное возобновление искусственных посевов путем самосева. Радиочувствительность экосистемы. Г.Г. Поликарпов предложил принцип зональности проявления биологического действия разных мощностей доз ионизирующих излучений [87]. Выделяют зоны: а) явного экологического нарушения (свыше 4-9 Гр/год); б) экологической маскировки (4-0,04 Гр/год); в) физиологической маскировки (0,04 - 0,005 Гр/год); г) радиационного благополучия (5-10 ~ 3 - 4·10-5Гр/год);· д) радиационной недостаточности, или неопределенности. Значимость биоценотических сдвигов после облучения можно обосновать, используя различные, принятые в экологии критерии (коэффициент сходства, энтропийная мера разнообразия, измерение биомассы и др.), для расчета которых требуются подробные сведения о видовом составе, численности и продуктивности видов, составляющих исследуемый биоценоз, подлежащий сравнению с контрольным [131]. По различным причинам такие всесторонние биоценотические учеты в ближней зоне не были проведены, тем более что в монокультурном сосновом лесу тотальное разрушение биоценоза вследствие гибели сосны было очевидным. В этом случае мерой радиационной устойчивости экосистемы может служить радиочувствительность ее доминирующей компоненты. У большинства экосистем радиочувствительность доминанта ниже других компонент на один-два порядка. Радиочувствительность доминанта в экосистеме не эквивалентна индивидуальной летальной дозе, а зависит еще и от ценотических связей. Радиочувствительность доминанта, выраженная в дозовых единицах, есть предел дозы, облучение свыше которого нарушает целостность экосистемы - экологический предел дозы [94]. Для хвойных деревьев значение летальной дозы и, следовательно, экологического предела дозы, обратно зависит от мощности дозы и изменяется в пределах от 10 Гр, полученных за срок в несколько дней, до 100 Гр, накопление которых было растянуто на 1000 дней. 4.5.4. Вторичные экологические изменения, вызванные эвакуацией населения Оценка антропогенного пресса на природные угодья.Только с территории 30-километровой зоны ЧАЭС было эвакуировано 135 тыс. жителей и 35 тыс. голов крупного рогатого скота. Нетрудно подсчитать, что прямая нагрузка этих составляющих на био(агро)-ценозы зоны составляла соответственно 3,2 т/км2и б - 7 С‚/км2, что на два порядка превосходило плотность населения крупных млекопитающих (зайцы, ко-пытные,лисицы) и в 50 раз плотность мышевидных грызунов или птиц. Кроме того, летом 1986 г. было отстреляно свыше 10 тыс. собак, оставленных хозяевами, в опасении возможной вспышки бешенства. На корню был оставлен урожай, в постройках остались запасы пищевых продуктов. Была прекращена промышленная эксплуатация леса и устранена рекреационная нагрузка на природные угодья. Для сорных и дикорастущих растений, диких животных сложились исключительно благоприятные стартовые условия после устранения подавляющих конкурентов - человека, его культурных растений, домашних животных. Была устранена также кормовая база для ряда специализированных насекомых-вредителей. Резко снизилось поступление в открытую среду пищевых отбросов и экскрементов, которыми ранее кормились разнообразные животные-мусорщики и копрофаги. Деградация лесных массивов.Лесные массивы занимают половину площади зоны отчуждения. На 3/4 это послевоенные посадки сосны по старопахотным полям и лугам. Почвенные условия на них часто не были оптимальными для деревьев. 20 - 40-летние сосняки к моменту катастрофы представляли собой монокультурные одновозрастные посадки, далекие от устойчивых биологически разнообразных природных сообществ. Без периодического прореживания и удаления больных и сухих деревьев посадки рано или поздно становятся перегущенными. Деревья тянутся в вышину, не имея для роста достаточно обширной кроны; прирост в толщину замедляется, отставшие в росте деревья засыхают. Ослабленные и мертвые деревья становятся местами размножения вторичных и стволовых вредителей, грибных гнилей. Самовозобновление соснового леса и появление подлеска из лиственных пород затруднено под сомкнутым пологом основной культуры. После эвакуации населения и прекращения промышленной эксплуатации леса на несколько лет приостановился не только уход за лесом (санитарные и проходные рубки), но даже расчистка просек и подъездных путей, прокладка минерализованных противопожарных полос. Из года в год накапливался сухостой, кварталы и просеки захламлялись валежником, образовывались неконтролируемые очаги размножения лубоедов, златок. и других вредных насекомых. Эти явления развивались медленно и были малозаметны. Последствия деградации монокультурных лесов сказались грандиозными лесными пожарами в сухой весенне-летний сезон 1992 г. Весенние пожары, начинаясь поджогами в населенных пунктах, быстро распространялись по сухой ветоши залежных полей (сгорело свыше 30 км2) и охватывали соседние леса. Летние пожары приняли катастрофический характер верховых. Общая площадь горельников только в украинской части зоны отчуждения составила, по разным оценкам, 120 - 150 км2, или 1/7 часть всех лесных массивов. Для сравнения укажем, что в соседних действующих лесхозах площадь потерь от пожаров не превышала 10 км2. Если к площади горельников добавить 35 км2лесов в междуречье рек Припять и Брагинка, которые погибли от подтопления при проведении гидротехнических защитных мероприятий в 1987 г., то потери леса от губительной радиации составили всего 3 - 5 % от рукотворных потерь, имевших причиной нерациональную деятельность, бездеятельность или злой умысел человека. Сукцессии растительности на залежных полях,В 1991 - 1992 гг. были предприняты детальные описания растительных сообществ и заложены геоботанические профили как в зоне экстренного отселения в 1986 гг. (Чернобыльский р-н), так и планомерного в 1990 г. (Полесский и Народичский районы) [39]. Формирование растительного покрова на залежах в целом проходит по известной схеме и включает несколько стадий: однолетних сорняков, олуговения и залесения [128]. В настоящее время зарастание полей, огородов и усадеб в зоне отчуждения находится на разных стадиях и зависит не только от специфики экологических условий, но и от других факторов - давности отселения людей, был ли собран урожай сельскохозяйственных культур, производился ли подсев трав. На значительной части исследованных полей возле сел, отселенных в 1990 - 1991 гг., стадия однолетних сорняков практически выпала и развиваются сообщества, в которых доминируют луговые виды. На приусадебных участках, дворах, клумбах, вдоль дорог формируются рудеральные ценозы, в которых доминирующие растения образуют отдельные куртины. На обогащенных азотом экотопах разрастаются крапива двудомная (Urtica dioica L.), лопух малый (Arctium minus (Hill) Bemh.), а в более бедных условиях - полыни обыкновенная (Artemisia vulgaris L.) и горькая (A-absinthium L.), осот полевой (Cirsium arvense (L.) Scop.) и другие вицы. Значительной является примесь как луговых видов, так и видов, ранее бывших в культуре. Самые сухие и бедные экотопы, песчаные гривы, которые не распахивались, заняты полынью полевой (Artemisia campestris L.), икотником серым (Berteroa incana (L.) DC.), ослинником двулетним (Oenothera biennis L.), цми-ном песчаным (Helichrysum arenarium (L.) Moench), булавоносцем седоватым (Corynephorus canescens (L.) Beauv.) и другими видами. В зоне отселения 1986 г. процессы демутации растительного покрова пошли значительно дальше. Летом 1986 г. сообщества сорняков на залежах формировали виды маревых (Atriplex, Chenopodium, Amaranthus), а на неубранных полях зерновых - щетинник (Setaria). Уже в следующем, 1987 г., а в особенности с 1988 г., сообщества однолетних сорняков были вытеснены сплошным покровом корневищных злаков. Большая часть распаханных ранее территории (до 90%)находится на стадии олуговения и занята сообществами пырея ползучего (Elytrigia repens (L.) Nevski), мощность подстилки в которых достигает до 7 - 10 СЃРј. На богатых почвах содоминирует ежа сборная (Dactylis glomerata L.), на более бедных и сухих - полевица тонкая (Agrostis tenuis Sibth.),BerotHK наземный (Calamagrostis epigeios (L.) Roth), була-воносец седоватый, на более влажных - полевица гигантская (Agrostis gigantea Roth) и щучка дернистая (Deschampsia caespitosa (L.) Beauv.). Стадия корневищных злаков самоподдерживается, так как плотная ветошь препятствует семенному размножению травянистых растений и особенно деревьев. В фауне зоны отчуждения отсутствуют крупные копытные, которые питались бы на полях и разрушали ветошь. В сухой сезон весной и осенью ветошь представляет серьезную пожарную опасность. В таких травянистых сообществах наблюдается появление невысокого подроста деревьев, чаще всего березы повислой, а в более сухих местах и поблизости от лесных насаждений - сосны обыкновенной. Ближе к населенным пунктам появляется клен ясенелистный (Acer negundo L.), а в населенных пунктах он уже формирует очень густые тенистые заросли, под которыми развивается бедный травяной покров. Из фруктовых деревьев нормально размножаются сливы и вишни, у культурных сортов яблони и груши наблюдается усыхание ветвей. На полях отмечены всходы груши обыкновенной (Pyrus communis L.) и яблони лесной (Malus praecox (Pall.) Borkh.), а также осины, на песчаных отложениях - робинии лжеакации. На увлажненных участках разрастаются различные виды ив, ольха клейкая (Ainus glutinosa (L.) Gaertn.), а на бедных песчаных отложениях - тополь белый (Populus alba L.) и черный (Р. nigra L.). Однако вдали от источников осеменения, на залежах, сеянцы деревьев отмечены всего на 5 % учетных площадок. Таким образом, в настоящее время растительность бывших сельскохозяйственных угодий и населенных пунктов находится на разных стадиях развития: сорно-пионерной, злаковой и лигнозной. Эти стадии рассматриваются как определенные временные ступени развития лесной растительности, где наибольшую площадь будут занимать сосновые и дубово-сосновые зеленомошные и черничные леса. Однако без участия человека процесс естественного лесовозобновления затянется на десятилетия. Массовое размножение грызунов.На залежных полях в 1987 г. произошло повсеместное массовое размножение мышевидных грызунов. До аварии численность грызунов на полях была невысокой и не превышала 20 - 30 особей/га, доминировали виды обыкновенной полевки (Microtus arvalis (Pall.)) и домовая мышь (Mus musculus L.), в меньшем количестве (до 14 %) встречались лесная и полевая мыши (Apodemus silvaticus (L.), A-agrarius (Pall.)), единично отмечались рыжая полевка (Clethrionomys glareolus (Schreber)) и землеройка - обыкновенная бурозубка (Sorex araneus L.). Численность грызунов в течение 1986 г. у с.Копачи оставалась низкой и не превышала 30 особей/га. Несмотря на многоснежную зиму 1986/1987 гг., благоприятную для выживания, к апрелю плотность мелких млекопитающих снизилась до 10 - 20 особей/га. Однако с началом репродуктивного периода численность грызунов начала быстро возрастать вследствие интенсивного развития копневищно-злаковой растительности, благоприятных погодных условий, прекращения губительной обработки полей и слабого в начале пресса хищников. К середине лета плотность полевок и мышей уже составляла 500 - 700 особей/га, а к октябрю достигла пиковых значений 2500 особей/га. В декабрьских отловах 1987 г. отмечено значительное снижение плотности мелких млекопитающих - до 750 особей/га. При такой высокой остаточной плотности популяции оставшихся запасов корма грызунам могло хватить лишь на 1 - 2 мес зимовки. Определенную роль в снижении численности грызунов сыграл возросший пресс хищников - лисиц, ласок, а в особенности дневных и ночных хищных птиц, количество которых в конце лета в 30-километровой зоне ЧАЭС значительно увеличилось. Вероятно, многие хищные птицы были привлечены в зону извне, по цепочке зрительной связи между соседними охотничьими участками птиц. В течение зимы 1987 - 1988 гг. произошла массовая элиминация грызунов. В апреле и мае 1988 г. на учетных площадках у с.Копачи встречались только обыкновенные полевки в количестве 80 - 100 особей/га. К сентябрю плотность грызунов снизилась еще вдвое. Вспышка массового размножения и последующий спад протекали одинаково на территориях с высоким радиационным фоном (учетные площадки у с.Копачи и с.Новая Красница) и на менее загрязненных территориях в юго-западном секторе 30-километровой зоны и междуречье рек Уж и Тетерев. В 1989 - 1990 гг. численность мелких млекопитающих на залежах стабилизировалась на уровне 100 - 180 особей/га; видовое разнообразие увеличилось. Добавились мышь-малютка (Micromys minutus (Pall.)), желтогорлая мышь (Apodemus flavicollis (Melchior)), землеройки - малая бурозубка (Sorex minutus L.) и малая белозубка (Crocidura suaveolens (Pall.)). Домовая мышь исчезла на всех полях, хотя и сохраняет высокую численность в постройках. Сохраняющийся видовой состав характерен для луговых ценозов [12]. Скопления птиц на временных водоемах.До аварии на ЧАЭС в Украине велись многолетние количественные наблюдения за местными и мигрирующими птицами в окрестностях Киевского водохранилища и на его акватории. Водно-болотный комплекс птиц - кряква (Anas platyrhynchos L.), чирок-трескунок (A.querquedula L.), лысуха (Fulica atra L.), цапля серая (Ardea cinerea L.), цапля большая белая (Egretta alba (L.)), чайки, лунь болотный (Circus aeruginosus L.) - здесь был обеднен по видовому составу из-за осушительной мелиорации, выпаса скота, сенокошения, охоты, строительства в береговой зоне р.Припять. Центром концентрации птиц были плавни и острова в верховьях Киевского водохранилища. Пространственное распределение водоплавающих птиц было резко нарушено в 1987 г., когда в результате строительства защитных плотин на мелиоративных каналах образовались мелководные пруды. Они привлекли водоплавающую птицу из соседних районов, общая численность уток и лысух в 30-километровой зоне на прудах достигала летом 15-20 тыс. особей, вдвое больше - в начале осени и до 50 тыс. особей - в пик миграции. Плотность населения водоплавающих птиц на различных водоемах 30-километровой зоны ЧАЭС до и после аварии, показана в табл.1.4.8. Осенью 1987 г. временные водоемы были спущены за ненадобностью, а птицы разлетелись. К 1989 г. плотность гнездовой популяции птиц, фоновых на водохранилище, снизилась на порядок (по данным А.Ю. Микитюка). Таблица 1.4.8 Плотность фоновых видов птиц водно-болотного комплекса на водоемах зоны отчуждения ЧАЭС (по данным А.Ю. Микитюка)
Водоем |
Год |
Плотность птиц, особей/га |
|||||
Кряква |
Чирок-трескунок |
Лысуха |
Цапля серая |
Чайка северная |
Крачка речная |
||
Киевское водохранилище |
1984 |
3,53 |
2,55 |
3,10 |
|
|
|
1987 |
0,05 |
0 |
1,57 |
0,022 |
0,955 |
0,511 |
|
1988 |
0,33 |
0,004 |
0,04 |
0,010 |
- |
|
|
1989 |
0,02 |
0,008 |
0 |
0,009 |
0,291 |
0,008 |
|
Рыбные пруды |
1987 |
3,00 |
0,50 |
0,43 |
|
|
0 |
1988 |
0,31 |
0,11 |
0,33 |
0,05 |
0,325 |
0,062 |
|
1989 |
0,04 |
0 |
0 |
0,61 |
0,9+88 |
0,008 |
|
Временные водоемы |
1987 |
14,18 |
1,40 |
0 |
|
0 |
0 |
1988 |
9,01 |
6,91 |
0 |
1,47 |
0 |
0 |
|
1989 |
0,04 |
0,001 |
0 |
2,21 |
0 |
0 |
|
Общая численность чайковых птиц (озерная чайка (Larus ridibundus L.), речная крачка (Sterna hirundo L.), малая крачка (Sterna albifrons Pall.), малая чайка (L. minutus Pall.)) сохраняется на высоком уровне, хотя продолжается их пространственное перераспределение. В пределах 30-километровой зоны ЧАЭС, по данным аэровизуальных учетов, в 1984 г. гнездилось 920 пар озерной чайки, в 1985 г. - 230, в 1986 г. - всего 200. Исчезли 8 колоний. Однако с 1987 г. численность чаек стала нарастать - соответственно 1340, 1640 и 1790 пар в 1987 г., 1988 г. и 1989 г. без учета громадной колонии в 2 тыс. пар, загнездившейся на струеразделяющей дамбе пруда-охладителя ЧАЭС. Несмотря на привлекательные условия обитания, успех размножения озерной чайки и речной крачки на загрязненных территориях был низким и исчислялся единицами процентов. К началу осени 1990 г. колония на пруде-охладителе полностью прекратила свое существование, исчезла также колония в устье р.Припять. По-видимому, рост популяции чаек происходил за счет миграции [12]. Население птиц, связанных с хозяйством человека.После эвакуации населения в 30-километровой зоне осталась домашняя птица и некоторое количество домашних хищников. Практически никто из них вне обитаемых населенных пунктов не перенес многоснежной зимы 1986 - 1987 гг., а домашняя птица была выбита хищниками к концу первой осени. За 1 - 2 года после аварии обезлюдевшие населенные пункты, пустые фермы и невозделываемые поля были покинуты грачами и скворцами (Stumus vulgaris L.). Две колонии грача (Corvus frugilegis L.), общей численностью в 1200 пар, которые существовали до 1986 г. вдоль дороги Чернобыль-Припять и в с.Лелев, после аварии и эвакуации прекратили свое существование. Однако в обитаемых населенных пунктах (Чернобыль, Припять, с.Залесье), где птицы могут кормиться отбросами, грачи скопляются тысячами на зимовку и гнездятся летом (300 пар). Здесь же находят пристанище голуби (Columba Uvia L.) и домашние хищники. Численность белого аиста (Ciconia ciconia (L.) осталась без изменений [12]. Размножение крупных зверей.Росту численности копытных способствовали мягкие, малоснежные зимы с 1987 по 1991 гг., прекращение охоты и беспокойства в лесах, переход зверей к укрытию в обезлюдевших селах (в садах и постройках). Из копытных наиболее быстро увеличилась численность многоплодного вида - кабана. С момента аварии она выросла в 8 - 10 раз. Размножение кабанов в 30-километровой зоне происходит повсеместно не исключая наиболее загрязненных участков ближней зоны (болота в "рыжем лесу" возле ЧАЭС, луга и кустарники припятской поймы). За сезон кабаны переворачивают дерновину на нескольких процентах площади лугов (максимум до 20 %). Эта их деятельность является одной из естественных форм фиксации радионуклидов на местности, а также способствует появлению сеянцев деревьев на минерализованных участках. Несколько медленнее, но пока устойчиво, нарастает численность лося. Рост численности косули заметно отстает из-за меньшей защищенности от крупных хищников - волков и одичавших собак. По усредненным данным наземных и аэровизуальных учетов в украинской [12] и белорусской (В.А. Коржев) частях зоны отчуждения, плотность населения различных видов крупных зверей может быть оценена такими числами ( в расчете на 1 км- угодий): кабан 2-6; заяц-русак до 1; лось 0.5 - 1; косуля и лисица 2-3. Волки (Canis lupus L.) до аварии были немногочисленны, примерно по выводку в каждом из охотничьих хозяйств. За послеаварийные годы численность этих хищников возросла в несколько раз и оценивается примерно в 80 особей на всю зону отчуждения, включая Полесский радиоэкологический заповедник. Животные, занесенные в "Красную книгу Украинской ССР", в зове отчуждения.К настоящему времени в безлюдных природных угодьях 30-километровой зоны сложился режим, близкий к заповедному и нарушаемый лишь браконьерами. Отсутствие беспокойства и хорошие кормовые условия привлекают множество птиц, в том числе из редких и охраняемых видов. Отметим из них орлана-белохвоста (Haliacetus albicffla L.), скопу (Pandion haliaetus L.), черного аиста (Ciconia nigra L.), серого журавля (Grus grus L.). Из охраняемых насекомых в зоне отчуждения и на отселенных землях Народичского р-на Житомирской обл. встречались бабочки махаон (РарШо machaon L.) и черный аполлон (Pamassius mnemosyne L.) [119].
Установленные государственными актами охраняемые территории (заказники, памятники природы, урочища), а их только в пределах украинской части 30-километровой зоны насчитывается 11, являются центрами воспроизведения и расселения ценных и редких видов животных и растений [27]. 4.5.5. Биоценотические изменения в водоемах Население пруда-охладителя ЧАЭС.На первых этапах создания и эксплуатации пруда-охладителя заросли водных макрофитов лишь начинали формироваться в прибрежных биотопах и существенной роли в функционировании водных экосистем не играли. Наиболее подробные исследования высшей водной растительности пруда-охладителя выполнены в 1989 г. Видовой состав растений в пруде-охладителе ЧАЭС в целом значительно беднее, чем в Киевском водохранилище, площади, занятые высшими видными растениями, в пруде-охладителе невелики и составляют около 1 га. Наиболее распространенным растением пруда-охладителя является тростник обыкновенный (Phragmites australis (Cav.) Trin. ex Steud.). Из нетипичных эффектов, зарегистрированных в пруде-охладителе в 1989 г., следует отметить наличие значительного превышения биометрических показателей генеративных органов рогоза узколистного (Typha angustifolia L.) над показателями из водохранилища (рис. 1.4.10). Аэровизуалъная оценка и аэровидеосъемка растительных сообществ пруда-охладителя ЧАЭС, проведенная в 1992 г. В.М. Клоковым, позволяет заключить, что за период с 1989 по 1993 гг. существенных изменений в особенностях распределения и запасах фитомассы не отмечено. В пруде-охладителе ЧАЭС до аварии обитало около 700 видов водорослей. В толще воды наиболее разнообразно были представлены зеленые и диатомовые водоросли, на дне и в обрастаниях твердого неорганического субстрата - диатомовые и синезеленые водоросли [57,90,127]. В качественных и количественных характеристиках фитопланктона и фитоперифитона происходили определенные изменения. Это было обусловлено отличиями в параметрах гидрологического режима при строительстве и реконструкции, дноуглубительными и другими гидротехническими работами, степенью обогрева и формированием специфического термического режима с перегревом до 28 - 30 °С, уровнем обогащения водоема биогенными элементами за счет коммунально-бытового сброса г.Припять, а также загрязнением технологическими стоками АЭС. Средневегетационная численность фитопланктона находилась в пределах 58 - 63 млн кл./л, биомасса - от 7 до 9 мг/л. По мере эксплуатации водоема количественные показатели развития фитопланктона имели тенденцию к повышению. Для водоема, особенно после 1982 г., было характерно "цветение" воды синезелеными водорослями, которое отмечалось обычно с конца мая до октября включительно, что было связано с обогревом. Основной возбудитель "цветения" воды - Microcystis aeruginosa Kutz. emend. Elenk. Показатели разовых запасов зеленых нитчатых водорослей в целом по водоему в 1980 - 1984 гг. колебались в пределах 4,65 - 57,0 т. Основной вклад в эту массу вносила водоросль Cladophora glomerata (L.) Kutz. Разовые запасы микроперифитона, представленного в основном синезелеными и диатомовыми водорослями (Lyngbya putealis Mont., Phormidium autumnale (Ag.) Gom., виды рода Oscillatoria), составляли 0,478 - 2,508 С‚. После аварии на ЧАЭС в 1986 - 1987 гг. существенных изменений как в видовом составе, так и по количественным показателям численности и биомассы водорослей не отмечено. Однако особенностью всех обнаруженных организмов было их плохое жизненное состояние. Проявлялось это в изменении окраски трихомов и нитей, в разрушении содержимого многих клеток, появлении морфологических аномалий. Многие организмы вследствие таких изменений даже не подлежали идентификации. Отмеченный факт можно рссматривать в первую очередь как симптом воздействия поверхностно-активных веществ (ПАВ компонентов дезактивационных смесей), в большом количестве попадающих в пруд-охладитель в результате комплекса проводимых мероприятий. Аналитические определения показали, что даже в устье р.Припять (буй 210) содержание катионакгивных ПАВ в воде регистрировалось на уровне 1,5 - 2,0 РјРі/Р", что по крайней мере на 1 - 2 порядка превосходило их критическую концентрацию для различных видов водорослей. Особенно негативное влияние оказывают ПАВ при повышении температуры среды, что, естественно, имеет место в пруде-охладителе ЧАЭС вследствие работы трех действующих блоков.
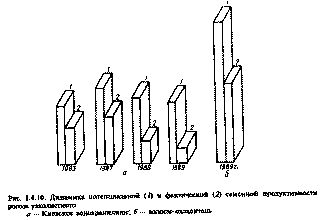
Рис. 1.4.10. Динамика потенциальной(Г)и фактической (2) семенной продуктивности рогоза узколистного: а -Киевское водохранилище;б -водоем-охладитель
Пруд-охладитель ЧАЭС до аварии использовался для промышленного рыбоводства. Подкормка рыбы после аварии на несколько лет прекратилась. Тем не менее собственных биологических ресурсов пруда хватало для поддержания населения рыб. Содержавшиеся в садках толстолобики белый (Hypophthalmichthys molitrix (Val.) и пестрый (Aristichthys nobilis (Richardson)) потребляли бактерио- и зоопланктон, фитоперифитон, детрит в количестве, достаточном для нормальной жизнедеятельности рыб [28]. У плотвы (Rutilus rutilus (L.)), красноперки (Scardinius erythrophthalmus (L.)) уклей (Albumus albumus (L.)), леща (Abramis brama (L.)), густеры (BUcca bjoerkna (L.)), карасей серебристого (Carassius auratus gibelio (Bloch)) и золотистого (С. carassius (L.)) не найдено специфических нарушений развития, которые можно было бы связать с действием радиации. Качество икры, ее оплодотворяемость, процент выхода мальков из оболочек не отличались от соответствующих показателей у рыб из Киевского водохранилища [103]. Популяции гидробионтов в Киевском водохранилище.Заполнение ложа Киевского водохранилища началось в 1965 г. Биоценоз этого рукотворного водоема не мог быть изначально устойчивым, так как население застойных и слабопроточных участков формировалось за счет поступления из рек, а дно - залитые пойменные луга и вырубленные леса - было обогащено органическими остатками, их минерализация продолжалась несколько лет. За счет подмыва берегов на десятилетие затянулось формирование литорали. Последние годы перед катастрофой ознаменовались новым антропогенным явлением - усилением стока органогенных элементов, поступавших с удобрениями на поля в бассейнах р. Днепр и р. Припять. Тем не менее к 1985 г. состав растительного и животного населения водохранилища стабилизировался, хотя отдельные виды испытывали обычные колебания численности в силу разных климатических, гидрологических и биотических причин. После Чернобыльской катастрофы продолжались наблюдения за видовым составом, численностью и биологическими показателями всех компонентов гидробиоценоза. Поэтому изменения в распространении, численности и жизненности отдельных видов оказались надежно зарегистрированными для сопоставления с данными многолетних наблюдений в доаварийный период [42]. Несмотря на заметное радиоактивное загрязнение воды и особенно донных отложений, дозовые нагрузки (максимум nЕ - 3 Гр/сут, обычно nЕ - 5 ...nРo - 6 Гр/сут) оказались недостаточными, чтобы вызвать экологически значимое радиационное поражение живых организмов. Полностью сохранилось биологическое разнообразие первичных продуцентов - планктонных водорослей и высшей водной растительности. Несколько теплых зим подряд (после 1987 г.) с отсутствием сплошного ледостава, теплые летние сезоны с прогревом воды на 2 °С выше многолетней средней температуры способствовали нарастанию биомассы фитопланктона на порядок в 1986 - 1987 Рі., массовым размножениям коловраток (класс Rotatoria), комаров-звонцов (семейство Chironomidae), моллюсков живородок (род Vivipams), распространению ряски (Spirodela polyrrhiza (L.) Schleid.), кувшинок желтой (Nuphar lutea (L.) Smith) и белой (Nymphaea alba L.). Эти вспышки, как водится, обрывались последующей депрессией численности. Увеличенный сток р. Припять летом 1988 г. привел к временному снижению биомассы первичных продуцентов в водохранилище и к усиленному выносу гуминовых веществ из заболоченного бассейна реки. Наряду с этими обычными явлениями последствия катастрофы отрицательно сказались на численности и видовом разнообразии некоторых особо чувствительных компонентов биоценоза. Но это были не радиационные последствия, а, как ни странно, результаты защитных мероприятий весны и лета 1986 г.: гидротехнические работы при постройке защитных сооружений в русле р.Припять и в водохранилище; дноуглубительные работы в Тетеревском заливе, вызвавшие взмучивание минеральной взвеси; забрасывание горящего реактора свинцом (2500 т), что привело к возгонке части этого опасного запаса, к повышению концентрации свинца в р. Припять в 8 раз к июню 1986 г.; отмывка дорог и техники растворами поверхностно-активных веществ, которые не поддаются быстрой биологической деструкции, и, в конечном счете, с поверхностями стоками частично попадают в р. Припять. Кроме того, поступление в воду тяжелых металлов (не обязательно радиоактивных) и постоянный радиолиз воды вызвали нежелательное усиление окислительных гидрохимических реакций, опасное для некоторых организмов [93]. Характерные примеры резких колебаний численности гидробионтов приводится в табл. 1.4.9. Они ранжированы по степени причинной связи с аварией от последствий химического загрязнения до независимых от аварии климатических колебаний и солнечной активности. Если на первых ступенях трофической пирамиды еще удается более или менее определенно высказываться о причинах изменений численности, то высшие ступени пирамиды оказываются вовлеченными в столь сложные экологические взаимосвязи, что трудно надежно утверждать, каковы движущие причины изменения численности отдельных видов рыб. Например, по данным учетов 1987 - 1989 Рі., заметно возросла встречаемость плотвы и язя (Leuciscus idus (L.)) в контрольных уловах и уменьшилась встречаемость красноперки, горчака (Rhodeus sericeus amarus (Bloch)), густеры, уклей. Таблица 1.4.9 Колебания численности фоновых компонентов биоценоза в Киевском водохранилище после Чернобыльской катастрофы [42]
Объект исследования |
Год |
Характеристика |
Участок отбора проб |
Численность |
Кратность |
Вероятная причина |
Фитопланктон в поверхностном слое воды |
1986 - 1990 |
Снижение содержания хлорофилла в массе воды |
Припятский залив |
7 - 49 мгк/л |
1,5 - 5 |
Химическое загрязнение, сброс в Припятьповерхностно-активных веществ |
Бактерио-планктон |
1986 - 1988 |
Численность упала до минимума за все время существования водохранилища |
Акватория водохранилища |
с 3 - 6 до 1,2 млн кл /л |
3 |
То же |
Зоопланктон |
Май 1986 |
Снижение численности |
Припятский залив |
0,03 г/м3 |
6 |
Химическое загрязнение свинцом, взмучивание минеральной взвеси |
Беантические ракообразные |
Лето 1986 |
Снижение численности до исчезновения |
Тетеревский залив |
- |
- |
Взмучивание минеральной взвеси при дноуглубительных работах |
Синезеленые водоросли |
Лето 1986 |
Вспышка размножения |
Акватория водохранилища |
30-50г/м3 |
5 -15 |
Потепление воды, усиленная инсоляция из-за искусственного разгонаоблаков в районе ЧАЭС |
КоловраткаBrachionus calyciflorus Pall. |
1989 |
Беспрецедентная вспышка массового размножения |
То же |
80 г/м3 |
|
Потепление воды, отсутствие сплошного ледостава зимой (вспышка началась еще в1985г.) |
Личинки комаров Criocotopus silvestris F. |
1986-1988 |
Вспышкаразмножения в заросляхпогруженной растительности |
Остров Домонтовский |
|
4 |
Потепление воды, наложившееся на11 летний цикл вспышек (1964 -1975 -1986 гг.) |
Ряска Spirodela polyrrhiza (L.) |
1987 -1988 |
Массовое размножение, увеличение размера листецов |
Акватория водохранилища |
|
|
Потепление воды, увеличенный речной сток радиостимуляция |
Моллюски Viviparus viviparus (L.) |
1986-1988 |
Вспышка размножения |
Устье Припяти, остров До-монтовский |
400-2000экз./м |
2,5-6 |
Потепление воды (однако в Днепровском заливе вспышки не было) |
Личинки трематод из моллюсков |
с 1987 |
Увеличение экстенсивности заражения |
Тетерев-ский залив |
79% |
5 |
Рост численности хозяев(оборванный эпизоотией в1989 г.) |
Зоопланктон |
1988 |
Снижение видового разнообразия |
Припятский залив |
20 видов |
3 |
Повышенное поступление гуминовых веществ летом 1988г. |
Водяной ослик Asellus aquaticus (L.) |
1986-1988 |
Снижение численности в донных ценозах миграция на растения |
Остров Домонтовский |
80 - 100 экз/м |
12 -18 |
Замор зимой1986/1987г. подтолстым ледовым покровом |