

8.5.Основні результати експериментальних досліджень та теоретичного анілізу.
Клітинний рівень ієрархії. Мірою радіобіологічного ефекту у клітин меристеми є їх здатність до ділення та утворення мікро- та макроколоній різного розміру. Практично всі клітини меристеми в залежності від ступеня ураження приймають участь у формуванні радіобіологічних ефектів на більш високих рівнях ієрархії.
Для математичного опису радіаційнного ураження клітин меристемі ряски найбільш адекватною виявилась ймовірностна модель радіаційного ураження - Ю.Г.Капульцевіча. Аналітично модель ураження клітин виражена слідуючою формулою:
![]() (8.5)
(8.5)
де Р - ймовірність виживання окремої клітини при опромінення в дозі - D; α – ймовірність втрати здатності до поділу після одного попадання у чутливу мішень клітини з об’ємом – v; m –ударність мішені (кількість попадань, котре здатне повністью інактивувати клітини; ( 1 – α )i -ймовірність для клітини схоронити здатність до поділу у клітини після i –попадань. Аналіз отриманих нами даних для виживаності різних дочірніх нащадків у Спиродели багатокореневої, показав, що для даного об’єкту досліджень характерно утворення більше 10 різних форм інактивації клітин. Наші оцінки параметрів моделі виживаності клітин, за даними експериментів та розрахунків: m ≈ 10, α ≈ 0,1, v ≈ 1/10 Гр . Таким чином спираючись на методи теорії надійності можна розрахувати всі необхідні параметри для моделі ураження на клітинному рівні.
Рівень субпопуляції клітин. Для даного об’єкту досліджень природною мірою радіобіологічного ефекту на рівні субпопуляцій клітин зачатків різного розміру та номеру, є їх здатність утворювати із висхідної кількості клітин зачатків (від 1-2 клітин до тисяч клітин) у процесі поділу необхідне критичне число клітин, достатне для формування зрілого дочірнього щитка –особини рослини. Це витікає з даних морфологічних та цитологічних досліджень даного об’єкту. Оскільки поділ клітин у складі зачатків проходить незалежно по паралельній схемі надійності, то отримано математичний опис радіобіологічного ефекту (виживаності ) на рівні субпопуляцій зачатків різного розміру, що привело до наступної моделі:
S(P)=![]() при
при
![]()
![]() при
при
![]() (8.6)
(8.6)
де S (P) – виживаність субпопуляцій зачатків розміром - no , P – ймовірність виживання поодиноких клітин у складі меристеми при даній дозі опромінення – D, T- середне число поділів клітин даної субпопуляції до формування необхідного критичного числа клітин - Nk. Ці числа Т – різні для різних no –субпопуляцій. Так для малих субпопуляцій пізніх дочірніх генерацій – вони можуть сягати до 10 мітозів, для великих субпопуляцій в тисячі клітин достатньо 1- 2 мітозів. -. Наши розрахунки та експериментальні результати показали, що - Nk дорівнює приблизно 10 тисяч клітин. По формулі (8.6) були обраховані теоретичні криві виживаності субпопуляцій різного розміру - no з різним припустимим числом мітозів –Т при різних дозах опромінення – D. На рис 8.6 представлені розрахункові дані про виживаність
|
|
Рис 8.6. Теоретичні криві вижваності S(p) субпопуляцій клітин різного розміру- no, за їх здатністю у відповідному числі клітинних поділів –Т утворити- Nk –критичне число клітин , достатне для виживаності особин при різних дозах опромінення– D. |
Рис
8.7. Криві виживаності ( |
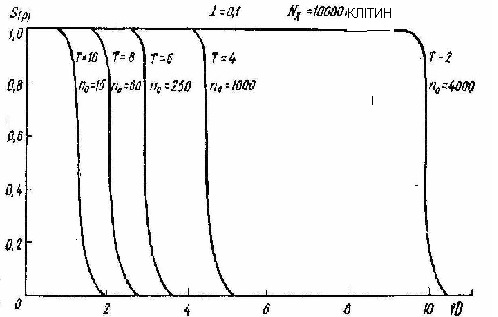
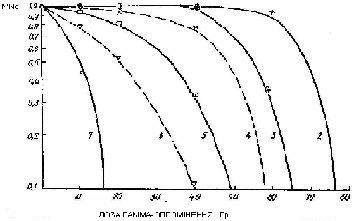
субпопуляцій різного розміру, а на рис 8.7 –представлені отримані нами експериментальні дані про виживаність дочірніх щитків різних генерацій. Видно, що теоретичні та експериментальні криві виживаності для різних генерацій по характеру, виду та основним параметрам непогано корелюють одне з одним. Особливо слід звернути увагу на форму дозових залежностей виживаності. Так, якщо для клітин по моделі Капульцевича криві виживаності мають класичну форму, що закінчується експоненційною ділянкою, то криві виживаності для субпопуляцій мають практично пороговий характер. Це пов’язано з тим, що якщо дозі такі , коли число виживаних клітин здатне за Т – мітозів сформувати критичне число клітин - Nk , достатне для повної виживаності особин , то виживаність буде становити 100%, а якщо недостатньо, то виживаність буде дорівнювати – 0. Тому і виникає пороговий тип дозової залежності.
Рівень виживаності популяції клітин усієї меристеми. Тут природня міра радіобіологічного ефекту - це кількість сформованих дочірніх щитків, оскількі основна функція меристеми - здатність генерувати нащадків. З урахуванням особливостей біології об’єкту досліджень, ясно що процес генерації дочірніх особин - це процес незалежний одне від одного, тобто практично діє паралельна схема забезпечення надійності генеративного процесу. Тому для формули виживаності меристеми можна використати спрощену модель для паралельного процесу у вигляді суми виживаності дочірніх нащадків. Оскільки виживаність змінюється від 1 до 0, то необхідне нормування на загальну кількість дочірніх генерацій-k. Тому виживаність меристеми, згідно теорії надійності, буде описуватися слідуючою простою формулою :
![]() (
8.7)
(
8.7)
визначається виживаністью саме критичного органу – меристеми - Sm.
На організменному рівні спостерігається найбільш чіткий прояв процесів старіння (Рис 8.7). Встановлено, щоСпиродела багатокоренева проявляє чітко виражений процес старіння. Видно, що ймовірність виживаності щитків навіть у контролі помітно зменшується , починаючи з 6-ої генерації, а вихід щитків у останніх 12-14 генераціях дуже малий. Відомо, що опромінення здатне прискорювати процес старіння.
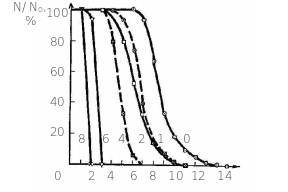
Рис
8.8. Залежність виживаності (![]() -%
)дочірніх щитків у різних генераціях
в нормі (0) та при різних дозах
гамма-опромінення 1 крад (10 Гр), 2 крад(20
Гр), 4 крад (40 Гр), 6 крад (60 Гр) та 8 крад (80
Гр).
-%
)дочірніх щитків у різних генераціях
в нормі (0) та при різних дозах
гамма-опромінення 1 крад (10 Гр), 2 крад(20
Гр), 4 крад (40 Гр), 6 крад (60 Гр) та 8 крад (80
Гр).
Для математичного опису нормального процесу старіння найбільш придатна модель і формула Гомпертца :
![]() (8.8)
(8.8)
де Sст - ймовірність генерації щитків n –генерації; В - параметр швидкості старіння; R – параметр накопичення помилок , як основної причини старіння. По експериментальним кривим старіння в нормі і при опроміненні можуть бути розраховані значення параметрів процесу старіння.
Аналіз на основі теорії надійності показав, що елементарні процеси радіаційного ураження і старіння незалежні одне від одного. Це дозволяє визначити виживаність організму, як добуток вживаності від радіаційного ураження на виживаність, що визначається процесами старіння . Математичний опис таких уявлень дозволяє запропонувати для оцінки виживаності організму слідуючу просту формулу :
![]() (
8.9)
(
8.9)
де Sm – виживаність організму ряски, що визначається, як раніше показано, виживаністью меристеми (), а Sст – виживаність ряски в процесі старіння.
Для аналізу реальних даних процесів старіння за допомогою формули Гомпертца зручно зробити перетворення цієї формули, таким чином , щоб привести її до лінійної форми. Це перетворення слідуючого типу :
![]() (8.10)
(8.10)
Воно дозволяє представити криві старіння у лінійній формі. Дані на рис 8.8 можуть бути представлені у лінієризованій формі. Такі криві старіння можна розрахувати для норми та при опромініенні у різних дозах. По них методом екстраполяції до осі ординат можуть бути розраховані та отримані дані про значення -В , та про значення – R. Зокрема, по жаних експерименту, оцінка цих параметрів показала, що в нормі В =0,04 , R = 0,38. А при опроміненні в дозі 20 Гр ці значення складають : В =0,08 , R = 0,37. Видно, що опромінення у два рази підвищує швидкість процесу старіння і мало впливає на параметр накопичення помилок. Таким чином, розроблені на основі теорії надійності моделі ураження багатоклітинного рослинного організму дозволяють прогнозувати закономірності радіаційного ураження на різних рівнях інтеграції рослини, ураховувати участь процесів старіння в радіобіологічних ефектах. Отримані моделі формують перспективний підхід до прогнозу і моделювання радіобіологічних ефектів у багатоклітинних системах на різних рівнях ієрархії.
Рівень популяції рослин. Дослідження про зміну кількості рослин в водній культурі Спиродели багатокореневої дозволили отримати експериментальні дані про радіобіологічні реакції вже на рівні популяції рослин. Встановлено, що початковий лаг-період змінюється на криву логарифмічного росту кількості особи в популяції рослин. Можна отримати дані також про зміни чисельності життєздатних особин (зелені щитки), нежиттєздатних (жовті щитки) , та чисельності адаптивних форм (туріонів – це форми, що зимують у водоймах). Така структура популяції рослин Спиродела багатокоренева у водній культурі. Мірою виживаності на рівні популяції природно вважати відношенні швидкостей росту чисельності в досліді до швидкості росту чисельності у контролі. На рис 8.9 представлені крива виживаності на рівні
|
Рис 8.9. Дозові криві виживаності для популяції Спиродели багато кореневої ( Cd / C0 ). Тут виживаність визначається через відношення швидкостей розмноження рослин при опроміненні і в контролі. Для порівняння тут наведено криві виживаності поодиноких рослин (N/ N0 ) від дози гамма-опромінення. |
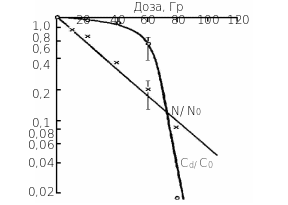
популяції рослин , а для порівняння тут наведено криву виживаності на рівні рослини, що ми вже описували. Видно сутт`єву різницю у формі та ході кривих виживаності на двох рівнях ієрархії. Виживаність на рівні організму поступово зменшується з дозою гамма-опромінення, а виживаність популяції спочатку мало міняється, а вже потім при дозах 60-80 Гр різко падає. Природа цих розходжень зрозуміла. При дозах опромінення 60 Гр і менше чисельність та життєздатність особин ряски високі (біля 20 %) і вони здатні підтримувати високу чисельність в популяції рослин своєю значною швидкістью розмноження (біля 60%), що дає змогу цьому виду рослин утримувати ареал життя і не давати себе витіснити конкурентам (зокрема іншим видам ряски – Lemna minor , Lemna tryasulka ). При збільшенні дози до 80 Гр особини ряски, що вижили(біля 10 %) по своєму стану та швидкості розмноження, вже не здатні підтримувати помітну швидкість розмноження (лише 9 %) , хоча їх ще достатньо багато.
В сосудах де вирощують популяцію ряски, спочатку експерименту їх чисельність відносно мала, тому вони можуть розмножуватись практично незалежно одне від одного, тобто по паралельній схемі щодо їх надійності. Для математичного опису росту такої популяції запропоновано і показана можливість використовувати слідуюче диференційне рівняння :
![]() (
8.11)
(
8.11)
де-
![]() - швидкість росту чисельності популяції
; N
– чисельність життєздатних особин в
популяції; K
– параметр швидкості росту чисельності
популяції ;C
– параметр, що характеризує швидкість
загибелі особин в популяції з урахуванням
їх старіння; Е - то параметр, що визначає
конкуренцію у популяції за рахунок
нестачі мінерального харчування та
перекривання від джерела світла, що
зменшує ефективність фотосинтезу і
харчування особин. Зрозуміло, що параметр
Е – починає діяти і себе проявляти
тільки при значній чисельності
популяції. Нами по експериментальних
даних отримані значення параметрів
цієї моделі росту для популяції. (рис
8.10).Показано, що з дозою опромінення
параметр швидкості росту (К-С),
- швидкість росту чисельності популяції
; N
– чисельність життєздатних особин в
популяції; K
– параметр швидкості росту чисельності
популяції ;C
– параметр, що характеризує швидкість
загибелі особин в популяції з урахуванням
їх старіння; Е - то параметр, що визначає
конкуренцію у популяції за рахунок
нестачі мінерального харчування та
перекривання від джерела світла, що
зменшує ефективність фотосинтезу і
харчування особин. Зрозуміло, що параметр
Е – починає діяти і себе проявляти
тільки при значній чисельності
популяції. Нами по експериментальних
даних отримані значення параметрів
цієї моделі росту для популяції. (рис
8.10).Показано, що з дозою опромінення
параметр швидкості росту (К-С),
|
Рис 8.10. Зміна величини параметрів диференційного рівняння росту популяції Спіродели багато кореневої в залежності від дози гамма-опромінення. |

помітно зменшується , а параметр конкуренції Е – зростає. Тобто, побудована на основі теорії надійності модель радіобіологічного ефекту на рівні популяції рослин адекватно описує суть процесу та отримані експериментальні дані.
Таким чином, побудовано множину якісних та кількісних моделей радіаційного ураження біосистем на різних рівнях ієрархії. Отримані моделі адекватно описують наші експериментальні дані для вищої рослини –Спірордели багатокореневої, для всіх ії радіобіологічних реакцій. Вважамо, що розроблені підходи та моделі на основі, розробленої теорії надійності біосистем, після відповідних змін придатні для опису радіобіологічних реакцій різних багатоклітинних організмів. Закладені в моделях уявлення створюють метод для дослідження радіобіологічних та біофізичних процесів в ієрархічних біосистемах. Використані тут ідеї та підходи базовані на теорії надійності можуть знайти широкий вжиток в дослідженнях на різних ієрархічних біологічних системах, що дуже важливо для радіобіології та загальної біології.