

Рис. З.1. Титульный лист журнала «Polygraph» Американской ассоциации полиграфа

Мы полагаем, что феномен успешного внедрения технологии полиграфных проверок в постсоветской России обусловлен, как минимум, пятью факторами, а именно:
последовательным и скрупулезным обобщением опыта США сотрудниками созданно го в 1975 году профильного подразделения КГБ СССР, а также параллельной ассимиляцией американского опыта «детекции лжи» специализированным подразделением ГРУ ГШ МО СССР;
творческой адаптацией (сотрудниками МВД России) североамериканской методологии полиграфных проверок к русским этнокультурным особенностям, а также к специфике ментали тета народов, населяющих постсоветское пространство;
частичным сохранением в России обломков военно-промышленного комплекса (НИИ, КБ, спецлабораторий), пока еще способных на основе разработанных в советскую эпоху техноло гий производить полиграфные устройства, а также программное обеспечение, не только не усту пающее, но и превосходящее в некоторых отношениях технику американских фирм-монополис тов — «Lafayette» и «Stoelting»;
высоким профессионализмом и энтузиазмом узкой группы бывших сотрудников спецслужб и правоохранительных органов, обеспечивших прочную кадровую основу для коммерческого при менения инструментальной «детекции лжи»;
постоянно растущим спросом на услуги коммерческих полиграфологов со стороны пред принимателей в связи со значительным ущербом, который ежегодно наносится отечественным предпринимателям наемным персоналом.
Возникает вопрос, как в Советском Союзе, несмотря на сформировавшийся в предвоенные и послевоенные годы резкий негативизм в отношении инструментальной «детекции лжи», удалось достичь высокого уровня развития данной технологии?
Как это ни парадоксально звучит, но в отделенный от остального мира «железным занавесом» СССР импульс, определивший в стране возрождение (после экспериментов Лурия и Леонтьева) и прогрессивное развитие «детекции лжи», пришел из-за рубежа.
Более того, импульс этот для представителей высшего военно-политического руководства Советского Союза пришел совершенно неожиданно. Дело заключалось в том, что в начале 60-х годов прошлого века «Штази»35 понесла на территории других государств значительные потери среди нелегальных агентов. Провалы произошли вследствие проведенных в отношении агентов «Штази» полиграфных проверок.
О произошедших инцидентах руководство восточногерманской разведки незамедлительно поставило в известность своих коллег из КГБ.
Почти одновременно с провалами агентуры «Штази» при помощи полиграфа в Соединенных Штатах был разоблачен легендарный советский разведчик Рудольф Абель36.
Случившееся заставило руководство КГБ объективно рассмотреть вопрос о полиграфных проверках и отдать вниз по властной вертикали распоряжение о проведении соответствующих научных исследований.Под руководством академика Павла Васильевича Симонова сотрудники Центральной военно-врачебной комиссии Медицинского управления КГБ СССР В. М. Наумов и А. А. Заничева провели ряд изысканий, направленных на выяснение прикладной ценности полиграфных проверок. Выводы научной группы относительно естественнонаучной обоснованности и практической полезности оказались однозначно положительными.
В 1975 году Председатель КГБ СССР Юрий Владимирович Андропов по итогам проделанной работы подписывает приказ об организации в структуре органов госбезопасности профильного подразделения по проведению полиграфных проверок и обучению собственной агентуры методам противодействия процедуре инструментальной «детекции лжи».
Первыми руководителями этого подразделения стали Юрий Константинович Азаров и Владимир Константинович Носков, которым удалось объединить под своим началом таких известных впоследствии полиграфологов, как В. В. Коровин, А. П. Сошников, В. Н. Федоренко и др.
Однако научно-прикладная работа по проблематике полиграфа, проводимая в 1970— 80-е годы в системе органов госбезопасности, в силу своей закрытости не получала в СССР какой-либо огласки в научной и научно-популярной печати, а о широком и открытом применении полиграфа не могло идти и речи37.
Радикальные изменения в сфере использования технологии полиграфных проверок начались лишь 1989 году, когда Советский Союз уже необратимо вступил в короткий период своего заката.
Начальной точкой отсчета этого процесса можно считать конец 1989 года, когда в Министерстве внутренних дел с целью изучения перспективы внедрения полиграфа в деятельность органов внутренних дел была создана рабочая группа, состоявшая из сотрудников Центрального аппарата.
В марте 1990 года члены группы Владимир Васильевич Гордиенко (в то время начальник отдела Главного управления уголовного розыска МВД СССР) и Сергей Викторович Игнатов (начальник одной из кафедр Академии МВД СССР) были откомандированы в Польшу для ознакомления с положительным опытом польских коллег в области применения полиграфа при раскрытии преступлений.
В результате проведенной работы рабочая группа пришла к выводу о целесообразности применения полиграфа в системе МВД и представила руководству Министерства отчет, содержавший в частности следующие рекомендации38:
использовать полученные данные в ходе проработки вопроса об опытном внедрении по лиграфных устройств в практику борьбы органов внутренних дел СССР с уголовной преступно стью;
подготовить публикации об опыте работы органов внутренних дел Польши по применению полиграфов в изданиях МВД СССР, а также предусмотреть подготовку учебного пособия по ис пользованию полиграфов с учетом опыта МВД Польши;
внедрить полученные материалы в учебный процесс и НИР Академии;
выйти с ходатайством к руководству МВД СССР о командировании специалистов Акаде мии и ГУУР в США для решения вопроса о приобретении контактных и бесконтактных измери телей психологического стресса и обучения методике применения таких устройств.
В период 1990—1991гг. происходит сближение позиций руководителей КГБ СССР и МВД СССР по вопросам применения полиграфа в деятельности указанных ведомств, и по просьбе
3руководства органов внутренних дел зимой 1991 года в одном из закрытых НИИ Комитета государственной безопасности началась теоретическая и практическая подготовка группы сотрудников МВД СССР.
К сожалению, процесс обучения был прерван вследствие приведших к развалу Советского Союза трагических событий 19—21 августа 1991 года39.
День 28 декабря 1994 года, когда был подписан приказ МВД России «Об утверждении инструкции о порядке использования полиграфа при опросе граждан», можно считать днем начала новейшего периода развития отечественной «детекции лжи», в ходе которого технологии полиграфных проверок, носившие до этого момента келейный характер, трансформировались в открытую и динамично развивающуюся высокотехнологичную индустрию.
На основании этого и последовавших за ним других приказов в системе органов внутренних дел стало возможным гласно и на законных основаниях проводить полиграфные обследования граждан в ходе расследования уголовных дел.
Первым руководителем специализированного подразделения центрального аппарата МВД стал Сергей Николаевич Зерин, которому удалось объединить вокруг себя группу молодых энтузиастов (С. В. Поповичев, П. И. Юдин и др.).
В это же время во ВНИИ МВД был организован профильный (А. И. Скрыпников, И. С. Зуб-рилова и др.) отдел, на который была возложена задача подготовки специалистов-полиграфоло-гов для нужд Министерства.
С этого момента Министерство внутренних дел Российской Федерации превращается в крупнейшего государственного пользователя полиграфа, а также в крупного поставщика специалистов в коммерческие структуры.
Ежегодно полиграфологи органов внутренних дел проводят тысячи полиграфных проверок различного целевого назначения, раскрывая при этом сотни преступлений.
На сегодняшний день МВД России является единственной организацией, которая регулярно проводит осенние международные научно-практические конференции, посвященные вопросам совершенствования теории и практики использования полиграфа как в рамках правоохранительной деятельности, так и в интересах бизнеса.
Следует отметить, что на начало 2004 года в системе МВД существуют две школы: московская (профильный отдел «закрытого» Главка) и так называемая «краснодарская» (подчиненный по властной вертикали отдел). Стремясь к максимальной объективности, мы вынуждены в некоторых моментах согласиться с мнением Ю. И. Холодного о том, что «...подготовка специалистов как московской, так и краснодарской школ осуществляется по программам, далеким от норм, установленных в мировой практике подготовки полиграфологов», при этом: «все это в комплексе ведет к тому, что «детектор лжи» из полезного прибора при раскрытии и расследовании преступлений превращается ...в инструмент нарушения прав человека»40.
В частности, мы полностью присоединяемся к точке зрения Ю. И. Холодного в отношении неудовлетворительного качества подготовки специалистов, которая на протяжении ряда лет осуществлялась в так называемой «краснодарской школе».
Косвенным подтверждением справедливости такой критики является то обстоятельство, что из числа успешно работающих коммерческих полиграфологов (бывших сотрудников МВД) мы не в состоянии назвать вообще кого-либо, подготовленного в «краснодарской» школе. В то время как число высокооплачиваемых специалистов, подготовленных на курсах ФСБ России, Московских курсах МВД России и московской коммерческой полиграфной школы «Ареопаг-М» превышает несколько десятков человек.
39
При этом тезис Ю. И. Холодного в отношении московской школы МВД является, на наш взгляд, недостаточно корректным, поскольку программа подготовки, утвержденная в московской школе МВД летом 2002 года (автор — С. И. Оглоблин), полностью соответствует программе курсов ФСБ России, а в некоторых аспектах даже превосходит ее.
Следует отметить, что руководство московской школы привлекает к чтению лекционного материала и проведению практических занятий как полиграфологов «новой волны» (С. И. Оглоблин, А. Ю. Молчанов, Т. П. Бывальцева), так и ветеранов КГБ — ФСБ (В. В. Коровин, А. П. Сошников, В. Н. Федоренко).
Только за 2003—2004 гг. московская школа МВД подготовила на безвозмездной основе41 55 специалистов (в том числе сотрудников других силовых структур) в соответствии с международными стандартами.
В настоящее время, помимо МВД и ФСБ, широко применяют полиграф Министерство обороны, Служба внешней разведки, Госнаркоконтроль, а также другие «силовые» и гражданские государственные организации и ведомства.
Как уже было сказано выше, неуклонно возрастает востребованность квалифицированных специалистов-полиграфологов в сфере бизнеса.
Решающий прорыв в сфере подготовки полиграфологов для оказания услуг частному предпринимательству был связан с основанием в 2002 году московской школы подготовки полиграфологов «Ареопаг-М».
Выпускники указанной школы уверенно завоевывают рынок коммерческого полиграфа в России и странах СНГ.
После появления на рынке профессионального полиграфа «Поларг», не имеющего аналогов в мире, можно констатировать, что московская полиграфная школа «Ареопаг-М» постепенно становится абсолютным лидером в области проведения коммерческих полиграфных проверок, а также в сфере подготовки коммерческих полиграфологов, деятельность которых (в отличие от специалистов государственного сектора) обладает большой спецификой.
Объективно оценивая события последнего десятилетия, мы присоединяемся к мнению Ю. И. Холодного о том, что «... история полиграфа в России вступает в свой следующий этап широкого признания и внедрения метода... в различные сферы государства и общества»42.
Завершая краткий обзор истории зарождения и развития технологии полиграфных проверок, мы позволим себе, во-первых, сделать прогноз о том, что число стран — пользователей полиграфа будет неуклонно возрастать, а во-вторых, признавая, что Соединенные Штаты являются пока еще в этой области «законодателями мод», прогнозировать в отношении американцев более жесткую конкуренцию со стороны ведущих полиграфологов России.
Глава 4.
ОБЩЕЕ ПОНЯТИЕ О ФИЗИОЛОГИИ. ОСНОВЫ АНАТОМИИ И ФИЗИОЛОГИИ НЕРВНОЙ СИСТЕМЫ ЧЕЛОВЕКА
Выше мы уже говорили о том, что генеральный принцип технологии полиграфных проверок заключается в соединении динамики скрытно протекающих в ответ на предъявленный стимул психических процессов с анализом параллельно развивающихся и доступных объективной регистрации физиологических процессов.
Отсюда следует, что специалист-полиграфолог должен ориентироваться как в базовых психологических категориях и законах, имеющих отношение к полиграфным проверкам, так и в основах тех физиологических процессов, которые объективно регистрируются в процессе инструмен -талъной «детекции лжи».
В соответствии с высказанным тезисом данный раздел курса будет посвящен изложению того необходимого объема знаний по физиологии человека, владение которым отличает квалифицированного полиграфолога от дилетанта, рассматривающего возникновение физиологических реакций в ответ на предъявленный в ходе полиграфной проверки стимул как нечто, что «таинственным образом» развивается в «черном ящике» человеческого тела.
С учетом того, что подавляющее большинство полиграфологов традиционно не имеет базового медицинского или психологического образования, мы стремились, с одной стороны, представить физиологическую тематику в максимально упрощенной форме, а с другой — пытались не опуститься при изложении физиологических основ полиграфных проверок до банальной примитивизации учебного материала.
Поскольку физиология человека как научная дисциплина неразрывно связана с анатомией (наукой, изучающей строение тела человека, его органов и систем), то мы были вынуждены при изложении той или иной физиологической тематики дополнять ее хотя бы в минимальном объеме соответствующими сведениями из курса анатомии.
Итак, физиология человека (от греч. physis — природа, logos — учение) — наука, изучающая закономерности функционирования человеческого организма в целом, а также функции его отдельных структур (систем, органов, тканей, клеток).
При этом физиология изучает не только функции структур, но и механизмы их регуляции, направленные на адаптацию организма человека в соответствии с динамикой его биологических потребностей к меняющимся условиям окружающей среды.
Центральным свойством любого организма является обмен веществ. И в одноклеточном, и в многоклеточном организме, вплоть до сложного организма высших животных и человека, обмен веществ представляет собой основу жизни. С прекращением обмена веществ наступает смерть. Поэтому физиология рассматривает человека как чрезвычайно сложный механизм, единственным назначением которого является поддержание собственного гомеостаза1 (постоянных внутренних условий существования организма, таких, например, как температура тела, артериальное давление или содержание кислорода в крови).
В процессе обмена веществ происходит высвобождение энергии, необходимой для жизнедеятельности организма.
В организме человека при распаде сложных питательных веществ или их элементов на более простые выделяется энергия. Эта выделившаяся энергия используется организмом для совершения работы, поддержания деятельности отдельных систем органов, а также для ассимиляторных процессов, т. е. процессов синтеза в организме необходимых ему сложных веществ и создания элементов тканей2.
Процессом, противоположным ассимиляции, является диссимиляция, т. е. разрушение и распад сложных веществ и элементов тканей.
Оба процесса в живом организме тесно связаны друг с другом: не может быть ассимиляции без диссимиляции и, наоборот, диссимиляции без ассимиляции.
В детстве, когда идет усиленный рост, то ассимиляция преобладает над распадом — диссимиляцией. В старости же, наоборот, распад, т. е. диссимиляция начинает преобладать над ассимиляцией.
В организме человека всегда идет непрерывный процесс разрушения тканей. Так, например, постоянно отмирают клетки верхнего слоя слизистой оболочки кишечника. Место этих отмерших клеток занимают новые полноценные клетки, постоянно развивающиеся в слизистой оболочке кишечника.
Непрерывному обновлению подвергается и наша кровь. Например, эритроциты, переносящие в крови кислород, живут всего около 130 дней. В течение каждых 130 дней красные кровяные тельца разрушаются и замещаются новыми. В течение жизни человека кровь полностью обновляется много раз.
Такому же непрерывному обновлению подвергается и наша кожа, у которой верхние слои клеток беспрерывно подвергаются ороговению, отмирают, слущиваются и заменяются новыми клетками.
Человеческая жизнь возможна лишь в том случае, если из внешней среды непрерывно поступают в организм кислород и питательные вещества, а в окружающую среду отдаются продукты диссимиляции, образующиеся в процессе жизнедеятельности организма, причем процессы обмена веществ протекают в гомеостатическом коридоре.
Гомеостаз целостного организма не является простой суммой реакций, направленных на поддержание отдельных параметров гомеостаза. Между системами регуляции отдельных параметров гомеостаза имеются сложные иерархические взаимодействия. При этом учитывается то, что какие-то гомеостатические параметры могут иметь в определенный момент большее значение для обеспечения жизнедеятельности человеческого организма в целом, в то время как изменение других параметров является в данном случае менее существенным. Например, в состоянии покоя на постоянном уровне поддерживается артериальное давление, однако при получении сильной физической нагрузки в первую очередь возникает необходимость полноценного снабжения мышц кровью. Деятельность сердца поэтому резко усиливается, что в свою очередь приводит к повышению артериального давления. Это означает, что на какой-то промежуток времени в системе гомеостатического приоритета угнетается деятельность механизма, направленного на поддержание стабильного уровня артериального давления.
В организме человека существует своеобразная иерархичность биологической значимости функций отдельных органов и систем. Это приводит к тому, что в определенных условиях некоторые системы должны продолжать функционировать даже в случае резкого нарушения других важнейших параметров гомеостаза. Например, для обеспечения нормального функционирования центральной нервной системы (ЦНС) требуется постоянное поступление с кровью большого количества кислорода и питательных веществ. Если в результате кровопотери поступление этих соединений резко снижается, то кровоток перераспределяется таким образом, чтобы обеспечить (насколько это возможно) продолжение нормальной деятельности ЦНС, при этом в других органах местный гомеостаз может нарушаться.
К регуляторным3 механизмам, в совокупности обеспечивающим гомеостаз организма, традиционно относят локальный, гуморальный и нервный механизмы регуляции.
Локальный механизм регуляции обеспечивает сохранение гомеостаза в пределах изолированного органа, в то время как гуморальный и нервный механизмы регуляции обеспечивают сохранность целостного организма.
В качестве примера локальной регуляции можно привести защитную функцию лейкоцитов (белых клеток крови), связанную с обезвреживанием инфекционных агентов. Так при попадании инфекции в порезанный палец лейкоциты оказывают на инфекционный фактор токсическое воздействие, нормализуя состояние тканей в области пореза.
Гуморальный механизм регуляции заключаются в том, что определенные биологически активные соединения (например гормоны коры надпочечников) выделяются во внешнюю среду железами внутренней секреции, а затем разносятся кровью по всему организму, оказывая специфический генерализованный эффект.
Ведущую роль в обеспечении взаимосвязи между отдельными частями организма и в обеспечении его деятельности в целом играет нервная система. Именно нервная система осуществляет наиболее совершенную регуляцию функций отдельных органов и систем организма, а также комплексную адаптацию организма человека к постоянно меняющимся условиям внешнего мира.
В механизме нервной регуляции различают два вида рефлексов4: безусловные, которые являются врожденными, и условные, которые приобретаются прижизненно на основе индивидуального опыта.
Нервная регуляция осуществляется по принципу рефлекторной дуги (пути, по которому идет возбуждение при осуществлении рефлекса). Рефлекторная дуга начинается рецептором, от которого в тот или иной нервный центр идут афферентные (центростремительные) нервные волокна. Из нервного центра к органу (эффектору) командный сигнал поступает по эфферентному (центробежному) нервному волокну (рис. 4.1). Нервный путь регуляции отличается от гуморального
Стимул
(I)
Стимул
,™
г, '—' (3) Сснсор-(2)
Рецептор или нейрон
(5) Эффектор
Реакция
(6) Реакция
Б елое МОЗГ
Серое
елое МОЗГ
Серое
Вставочный нейрон
(4)
Моторный нейрон
Рис 4.1. Схема рефлекторной дуги спинномозгового рефлекса
3 Под регуляцией в физиологии понимают минимизацию отклонений в деятельности систем и органов, оптимизацию их деятельности.
л Рефлекс — это ответная реакция организма на экзогенное (внешнее) или эндогенное (внутреннее) раздражение рецепторов (нервных окончаний, приспособленных к восприятию раздражений определенного вида), осуществляемая через центральную нервную систему.значительно большей скоростью оказания адаптирующего влияния, а также возможностью оказывать воздействие на строго определенный орган.
Сложный организм представляет собой единое целое. Целостность организма, находящегося во взаимодействии с окружающей средой, обеспечивается, прежде всего, нервной системой и ее ведущим отделом — корой головного мозга. Кора головного мозга весьма тонко и точно улавливает изменения внешней среды, а также внутреннего состояния организма и своей деятельностью обеспечивает приспособление организма к окружающей среде.
В последующем изложении избранной физиологической тематики мы, согласно традиции, будем рассматривать деятельность различных органов и систем отдельно, однако надо всегда помнить, что это делается лишь с целью облегчения их изучения. В нормальном же целостном организме все органы тесно связаны между собой, находятся в сложном взаимодействии, объединены в одно целое не только структурно, но и функционально.
В этом разделе мы последовательно рассмотрим деятельность нервной, сердечно-сосудистой и дыхательной систем (поскольку динамику активности именно этих систем анализирует специ-алист-полиграфолог), далее мы обсудим кожно-гальваническую реакцию (КГР) с точки зрения ее использования в процессе инструментальной «детекции лжи». В заключение раздела мы дадим общее понятие о функциональном состоянии.
Итак, обратимся теперь в соответствии с данным планом к изложению особенностей анатомического строения и функционирования нервной системы человека.
Нервная система человека построена из огромного количества разнообразных клеток. Основной функциональной единицей нервной системы является нейрон (нервная клетка), который специально предназначен для приема, обработки, хранения и передачи информации в форме электрических импульсов.
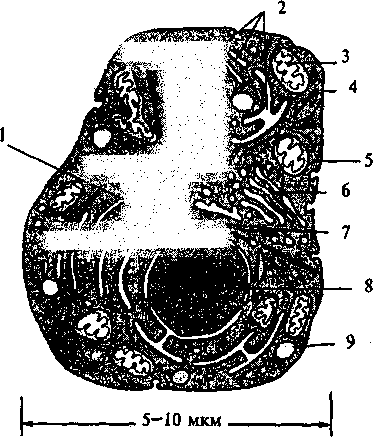
Нейрон содержит в себе все структуры, которые обеспечивают жизнедеятельность типичной клетки (рис, 4.2). Центральное положение в клетке занимает ядро, которое играет роль хранилища генетической информации организма. Кроме ядра в цитоплазме (внутренней среде клетки) содержится множество других структур (органелл). Так, на эндоплазматическом рети-кулуме закреплены рибосомы, которые в соответствии с генетическим кодом участвуют в сборе молекул белка, митохондрии осуществляют поддержание необходимого количества АТФ (аденозинтрифосфата) — основного источника энергии клетки и т. д. Цитоплазма и содержащиеся в ней органеллы отделены от окружающей среды мембраной, имеющей сложное строение.
Рис. 4.2. Строение «стандартной» клетки:
1 — центриоль; 2 — эндоплазматический ретикулум; 3 —
митохондрии; 4 — плазматическая мембрана; 5 — пито-
золь; 6 — аппарат Гольджи; 7 — волокна цитоскелета; 8 —
ядро; 9 — лизосома.
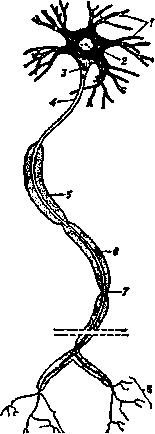
Внешние формы нейронов чрезвычайно многообразны, однако основные части у всех нейронов являются стандартными. Любой нейрон (рис. 4.3) содержит сому (тело) и многочисленные отростки. Отростки подразделяются на аксоны, через которые возбуждение передается
от одного нейрона к другому, и дендриты (от греч. дерево) через которые нейрон принимает информацию через синапсы (от греч. контакт) от других нейронов. Необходимо отметить, что нейрон обладает свойством внутренней односторонней проводимости, т. е. возбуждение в нем передается только от дендрита к аксону.
Главное свойство нейрона — способность к возбуждению (генерации электрического импульса) и передаче возникшего возбуждения (импульса) другим нейронам, мышечным, железистым и иным типам клеток.
В нейроне возбуждение развивается вследствие процессов, происходящих на клеточной мембране, строение которой изображено на рис. 4.4.
Рис. 4.3. Строение нейрона — главной клетки нервной системы:
1 — дендриты; 2 — сома (тело клетки); 3 — аксонный
холмик; 4 — аксон; 5 — миелиновая оболочка; 6 — ядро
шванновской клетки; 7— перехват Ранвье; 8 — эффектор-
ные нервные окончания
Мембрана нейрона имеет многослойное строение: два слоя молекул белка, между которыми находится слой молекул липидов (жиров). Толщина такой мембраны примерно в 10 тыс. раз тоньше человеческого волоса. В нейронную мембрану «вмонтированы» разного рода частицы, которые пронизывают мембрану насквозь, образуя так называемые ионные каналы, через которые способны проходить ионы натрия, калия, кальция, хлора и некоторые другие ионы. Другие частицы закреплены на внешней поверхности мембраны и включают в себя как белки, так и полисахариды. Эти структуры представляют собой рецепторы, чувствительные к различным химическим агентам, например гормонам или медиаторам (передатчикам возбуждения в нервной системе).
Рис. 4.4. Мембрана нейрона
1 — липиды (жиры); 2,3 — различные виды белков; 4 — полисахариды
Ц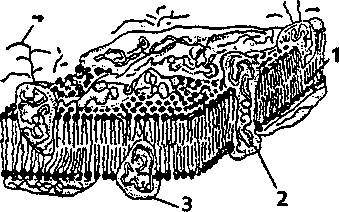 ентральную
роль в возбуждении нейрона играют
ионные
каналы (рис. 4.5). Различают два вида
ионных каналов: ионные насосы (помпы)
и потенциалзависимые каналы.
Ионные насосы работают постоянно,
откачивая из цитоплазмы
нейрона ионы натрия и закачивая внутрь
клетки
ионы калия. Эти насосы постоянно требуют
энергии, однако,
благодаря их работе в цитоплазме (внутри
клетки) поддерживается
разность ионных концентраций ионов
натрия
и калия: внутри клетки концентрация
ионов калия примерно
в 25—30 раз превышает их концентрацию в
межклеточной
среде. Концентрация ионов натрия вне
клетки, напротив, примерно в 50 раз
выше, чем внутри. Таким образом,
благодаря работе ионных насосов, между
цитоплазмой и
ентральную
роль в возбуждении нейрона играют
ионные
каналы (рис. 4.5). Различают два вида
ионных каналов: ионные насосы (помпы)
и потенциалзависимые каналы.
Ионные насосы работают постоянно,
откачивая из цитоплазмы
нейрона ионы натрия и закачивая внутрь
клетки
ионы калия. Эти насосы постоянно требуют
энергии, однако,
благодаря их работе в цитоплазме (внутри
клетки) поддерживается
разность ионных концентраций ионов
натрия
и калия: внутри клетки концентрация
ионов калия примерно
в 25—30 раз превышает их концентрацию в
межклеточной
среде. Концентрация ионов натрия вне
клетки, напротив, примерно в 50 раз
выше, чем внутри. Таким образом,
благодаря работе ионных насосов, между
цитоплазмой и
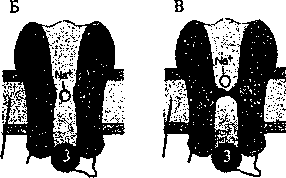

Цитоплазма
Цитоплазма
Рис. 4.5. Натриевый потенциалзависимый канал:
-. — 1знал открыт; Б — канал инактивирован; В — канал инактивирован и закрыт; 1 — мебрана; 2 — потенциалзависимый
канал; 3 — инактивирующая частица
66
внеклеточной средой на мембране нейрона возникает потенциал. Внутренняя сторона мембраны (цитоплазма нейрона) заряжается относительно внешней стороны (внеклеточного пространства) отрицательно на величину около — 70 мВ.
Как мы уже говорили, главное свойство нейрона — это способность возбуждаться и передавать возбуждение другим клеткам. Когда говорят о способности нейрона к возбуждению, то имеют в виду способность нейрона к генерации потенциала действия (возникновения разницы потенциалов обратного знака в сравнении с мембранным потенциалом), обусловленного кратковременными изменениями в ионной проводимости мембраны за счет процессов активации и инактивации потенциалзависимых натриевых и калиевых ионных каналов. Ионы натрия и калия движутся по этому типу каналов в соответствии со своими концентрационными градиентами (из места высокой концентрации к месту низкой концентрации).
В состоянии покоя натриевые каналы находятся в закрытом положении и внутренняя поверхность мембраны заряжена отрицательно по отношению к внешней стороне примерно на 70 мВ. В том случае, если происходит пороговая (приблизительно 10 мВ) деполяризация (уменьшение поляризации) мембраны, то натриевые ионные каналы открываются и внутрь нейрона под давлением концентрационного градиента лавинообразно устремляются положительно заряженные ионы натрия. Положительный ток ионов натрия смещает потенциал мембраны в сторону деполяризации примерно до уровня в + 55 мВ (величина натриевого равновесного потенциала). Эта величина положительного потенциала обусловлена установлением определенных концентраций ионов натрия в цитоплазме и вне клетки. Примерно через 0,5 — 1 мс потенциалзависимые натриевые каналы закрываются специальной белковой молекулой. Одновременно открываются калиевые каналы, и положительные ионы калия начинают в соответствии со своим концентрационным градиентом стремительно выходить из клетки, возвращая мембранный потенциал в исходное состояние (процесс реполяризации). После этого нейрон снова готов к генерации следующего потенциала действия. Общая длительность нейронного потенциала действия составляет около 1 мс. Схематическое изображение процессов, происходящих при генерации потенциала действия, приведено на рис. 4.6.
В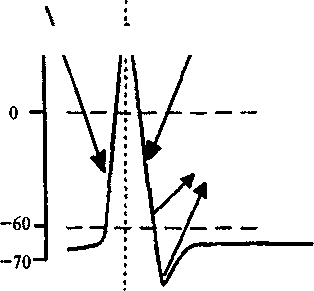 хождение
в нейрон Na+
хождение
в нейрон Na+
56 -| , rl ,^ Натриевая
инактивация
Выход из нейрона К+
02 S
1 мс
Рис. 4.6. Генерация нейронного потенциала действия
После того, как в нейроне возникает возбуждение (генерируется потенциал действия), его необходимо передать другому нейрону или клетке иного типа, например мышечной или железистой.
Импульс покидает тела нейронов по аксонам5, которые для улучшения проводимости, как правило, покрыты миелиновой оболочкой (рис. 4.7).
5 Аксоны отдельных нейронов большей частью объединяются в пучки (нервы). Аксоны отдельных нейронов в нервах называют нервными волокнами.
Гяэва 4
67
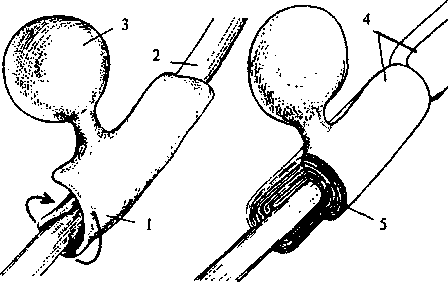
Рис. 4.7. Образование миелиновой оболочки на аксоне:
1 — наматывание слоев миелина; 2 — аксон; 3 — шваннов-ская клетка; 4 — перехват Ранвье; 5 — миелиновый чехол
Миелиновая оболочка периферических нервных волокон образуется специальными шван-новскими клетками, разновидностью глиаль-ных6 клеток. Миелин обладает большим электрическим сопротивлением и служит изолятором аксона. Миелиновый покров прерывается через регулярные промежутки равной длины, оставляя при этом открытыми участки мембраны шириной около 1 мкм. Эти участки получили название перехватов Ранвье. Перехваты образуются примерно через каждые 0,5—1,5 мм. Это обусловлено тем, что разные участки миелиновой оболочки образуются отдельными глиальными клетками, обволакивающими эти участки. Прак--нчески все нервные волокна в центральной не-гзной системе миелинизированы. Лишены мие-:нновой оболочки только волокна, управляю-_,:е медленно работающими внутренними органами, такими, например, как кишечник или мочевой пузырь (а также волокна, проводящие афферентные импульсы о температуре и боли). Про-:f :: миелинизации завершается у человека лишь после окончания полового созревания (у ново-: : : генного миелинизировано около 2/3 нервных волокон). Миелин играет важную роль, выпол-• q изолирующую, опорную, транспортную и, возможно, трофическую роли. Процесс демиели--_;-:>ации нервных волокон ведет, например, к такому тяжелому заболеванию, как рассеянный ; :*_-ероз, постепенно и незаметно приводящий к параличу движения.
Скорость проведения возбуждения по миелинизированному нервному волокну значительно ; к~е. чем по безмякотным нервным волокнам, что обеспечивает высокую скорость работы не-: -• :-:ых сетей мозга человека.
Распространение нервного импульса (воз-
Направление распространения •
•ждения) по немиелинизированным и мие-
"°
" -:ннзированным волокнам осуществляется
-:ннзированным волокнам осуществляется
.-разному. На рис. 4.8 схематично изображе-
3-5 О
ЮД I
В i
33 I -701
Порог
- ' j проведение нервного импульса по безмие-
Область рефрактерности
• -'.новому нервному волокну.
Расстояние, см
Рис. 4.8. Распространение возбуждения по немиели-низированному волокну
На возбужденном участке аксона внутрен--.'Л сторона мембраны заряжена положитель-яю по отношению к экстраклеточной среде. На внутренней стороне мембраны аксона на не-*юз5}:жденном участке находится отрицатель-зм заряд. При этом между возбужденным и •зевозбужденньш участками образуется раз-
ность потенциалов и начинает течь ионный
:«_ Возбуждение (процесс распространения
дэихтяризации) движется по аксону только в
сюльшая часть головного мозга занята клетками глии (от греч. склеивать). Глиальные клетки выполня-
* в нервной системе разнообразные вспомогательные обслуживающие функции, освобождая нейроны от
ЛЕЖгельностл, связанной с защитой от механических повреждений, удалением продуктов диссимиляции, пита-
ивем и т. д. Миелиновый чехол образуется клетками олигодендроглии, одной из разновидностей глиальных
ватт-:'*: Миелнновые клетки в отличие от нейронов сохраняют способность к делению на протяжении всей
68
РАЗДЕЛ
одном направлении, поскольку после возбуждения участка волокна в нем наступает период рефрактерное™7 (невозбудимости), обусловленный инактивацией (блокадой) потенциалзависимых натриевых каналов, которые и обеспечивают деполяризацию мембраны. В немиелинизиро-ванном волокне (аксоне) потенциал действия последовательно развивается в каждом соседнем участке, поэтому скорость распространения возбуждения невелика.
Рис. 4.8. Схема синапса:
1 — везикулы; 2 — митохондрии; 3 — преси-
наптическая мембрана; 4 — синаптическая
щель;5— ионный канал; 6 — постсинаптичес-
кая мембрана
Миелинизированные волокна, напротив, отличаются высокой скоростью распространения деполяризационной волны, т. к. потенциал действия возникает в них только в области перехватов Ранвье, перескакивая через участки изолированной мембраны. Иначе говоря, в миелинизиро-ванном волокне деполяризационная волна движется скачкообразно от перехвата к перехвату. Так, благодаря серии нервных импульсов в нервной системе передается информация. Между нейронами, а также между нейронами и мышечными клетками существуют области контактов, в которых возбуждение передается с нейрона на нейрон или
на другую клетку (к примеру клетку стенки сосудов или клетку сердечной мышцы). Такие контакты (рис. 4.8) получили название синапсов.
Синапс является образованием, имеющим крайне маленький размер (не более 1 мкм). На одном нейроне, как правило, располагаются несколько тысяч синапсов, по которым в нервной системе передается информация. Принцип работы химического синапса заключается в том, что электрический потенциал действия, пришедший по аксону к пресинаптическому окончанию, превращается в нем (пресинаптическом окончании) в химический импульс.
Последовательность событий при активации синапса (рис. 4.9) такова: нервный импульс подходит к пресинаптическому окончанию, что приводит к открытию кальциевых ионных каналов, и ионы кальция начинают входить внутрь окончания, связываясь с везикулами (пузырьками), в которых содержится медиатор (нейротрансмиттер, передатчик) — биологически активное вещество. В результате медиатор, который содержится в везикулах, изливается в синаптическую щель. Затем молекулы медиатора связываются с рецепторами8 на постсинаптической мембране и открываются химическизависимые ионные каналы, расположенные в постсинаптической мембране. Ионы натрия (калия) начинают движение согласно своим концентрационным градиентам, т. е. на мембране происходит изменение потенциала. Это изменение потенциала покоя на мембране постсинаптического нейрона получило название постсинаптического потенциала. Величина постсинаптического потенциала зависит от количества трансмиттера, связанного хеморецеп-торами. При достижении критического уровня деполяризации, т. е. когда постсинаптический потенциал преодолевает порог реакции, на постсинаптической мембране возникает потенциал действия, который начинает распространяться по постсинаптическому нейрону. Молекулы трансмиттера немедленно удаляются из синаптической щели и вновь собираются в везикулы преси-наптического нейрона.
Необходимо заметить, что выделение молекул медиатора (под влиянием пресинаптических импульсов) в синаптическую щель происходит отдельными порциями, поэтому постсинаптичес-
7 Во время прохождения волны деполяризации наступает время абсолютной рефрактерности, около 1 мс, когда клетка в ответ на раздражение не будет производить нервный импульс. За этим периодом следует период относительной рефрактерности, около 5—10 мс, когда нейрон будет генерировать потенциал действия только в ответ на сильный импульс.
8 Такие рецепторы связывают молекулы только одного типа медиаторов, т. е. работают по принципу «ключ к замку».
69
Аксоны
Дендриты
Синаптические
окончания
Тело
нейрона
Аксон
Направление
нервного импульса
Пресинаптическое
окончание
Синаптические
везикулы
Аксон
Митохондрия
Дендрит
Синаптическая
щель
Синаптическая
щель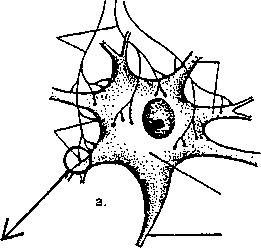

Рис. 4.9. Схема синаптической передачи
<я потенциал возникает не одномоментно, а только в случае суммации последовательных депо-гркзаций постсинаптической мембраны, когда постсинаптический потенциал достигает критической величины.
На мембране одного и того же нейрона могут находится как возбудительные, так тормозные
ззлсы. В возбудительных синапсах процесс деполяризации постсинаптической мембраны обус-
ален активацией натриевых ионных каналов, а гиперполяризация (увеличение отрицательного
ipaua на внутренней стороне мембраны) постсинаптической мембраны вызывается увеличением
•рицинин I и ионов хлора и калия.
Таким образом, нейрон через соответствующие синапсы может возбудиться или затормо-тъся.
Рассмотренный нами химический механизм синаптической передачи говорит о том, какую •ичнтельную роль в деятельности нервной системы играют медиаторы или нейротрансмитте-уи В ЦНС функцию медиаторов выполняют около 30 биологически активных веществ. Основ-ив»е медиаторы — это ацетилхолин и норадреналин. Нейроны, выделяющие норадреналин, называет адренэргическими, а ацетилхолин — холинэргическими. К медиаторам, выполняющим в " НС важные функции, относятся также адреналин, серотонин, дофамин, эндогенные опиоиды зга, ГАМК (гамма-аминомасляная кислота) и др.
В качестве примеров, подчеркивающих значимость роли медиаторов, можно указать на то, т: ~ри недостатке дофамина (синтезируется в среднем мозге) у человека развивается такая тя-жа_гая болезнь, как болезнь Паркинсона (дрожательный паралич), а истощение в головном мозге веротонина приводит к хронической бессоннице. Что касается эндорфинов (относящихся к группе эшюгенных опиоидов), то их функции связаны с механизмом снижения ноцицептивной (болевой) тэствительности.
Итак, мы коротко изложили особенности функционирования основной единицы нервной овстемы — нейрона. Рассмотрим теперь схематично общее анатомическое строение нервной системы, ее основные структуры и их функции.
Важнейшие составные части нервной системы приведены на рис. 4.10.
Нервная система
Центральная нервная система
Периферическая нервная система
1 |
|
Вегетативная Сомати нервная, система нервная |
ческая система |
! Головной мозг |
1 Спинной мозг Симпат нервная Тол СПИ! мозп нер |
||
ическая Парасим-система патическая нервная система |
|||
Спинно- Черепно-ъко мозговые мозговые шо- нервы нервы |
|||
жые I """' | Гпиннп- Чеоепно- |
|||
мозговые мозговые нервы нервы
Рис. 4.10. Важнейшие составные части нервной системы позвоночных
Из схемы видно, что ЦНС состоит из головного и спинного мозга. Спинной мозг (рис. 4.11) находится в позвоночном канале и имеет вид цилиндрического тяжа, который сверху переходит в продолговатый мозг.
Продольные борозды, которые проходят по передней и задней поверхностям спинного мозга, делят его на две половины — правую и левую. В центре спинного мозга проходит канал, заполненный спинномозговой жидкостью. Вокруг канала располагается серое вещество (тела нейронов), имеющее на поперечном разрезе форму бабочки. Передние выступы серого вещества называют передними рогами, а задние выступы — задними рогами. Серое вещество окружено белым веще-

3 12
Рис. 4.11. Поперечный разрез спинного мозга:
1 — белое вещество; 2 — спинномозговой канал; 3 — задняя продольная борозда; 4 — задний корешок; 5 — спинномозговой узел; 6 — спинномозговой нерв; 7 — серое вещество мозга; 8 — передний корешок; 9 — передняя продольная борозда; 10 — задний столб; 11 — боковой столб; 12 — передний столб:;глинизированные отростки нейронов). Белое вещество делится на три столба (задний, .:: и передний). Отростки нейронов образуют проводящие пути, соединяющие нервные цен-. -: энного мозга друг с другом и с нервными центрами головного мозга.
От спинного мозга отходят 31—32 пары спинномозговых нервов (спинномозговые нервы ,-с 12 парами черепномозговых нервов, вегетативными узлами и нервными окончаниями йоазуют периферическую нервную систему).
Гпннномозговые нервы начинаются двумя корешками — передним и задним. В задних ко-: ..о: расположены спинномозговые узлы, в которых находятся тела сенсорных (чувствитель-I- ^_..„ • нейронов. Один отросток сенсорного нейрона идет на периферию (мышцу, кожу и т. д.) и .«гч'-гчнвается там рецептором. Другой отросток в составе заднего корешка либо вступает в спин-нж мозг, а затем либо заканчивается в сером веществе (в задних рогах) спинного мозга, либо лостигает продолговатого мозга в составе проводящих путей (белого вещества). В передних ко-проходят отростки моторных (двигательных) нейронов, направляющиеся в мышцы. Че-•. г~дние корешки проходят также эфферентные (центробежные) волокна вегетативной не-; нстемы, о которой будет сказано ниже. Тела мотонейронов расположены в передних рогах - : го мозга.
. :.-;-. же как и головной, спинной мозг окружен оболочками. Между спинным мозгом и оболоч-находится спинномозговая жидкость (ликвор), которая предохраняет мозг от сотрясений и 1гж"г~.{х неблагоприятных механических воздействий.
.' плнной мозг выполняет две функции — рефлекторную и проводниковую. Спинной мозг
«;«с^еч11вает простые рефлексы, которые осуществляются без участия головного мозга, такие как
-т"-. -ельные и разгибательные (например, хорошо всем известный коленный рефлекс, схема
: г э изображена на рис. 4.1. А), а также тонические рефлексы, обеспечивающие поддержание
г.тенного напряжения (тонуса) мышцы. Спинномозговые рефлексы являются безусловны-
- - лняются (разумеется, исключая патологию) в течение всей жизни индивида. К сложным
: спинного мозга относится шагательный рефлекс — согласованная двигательная ак-
. верхних и нижних конечностей. Для осуществления шагательного рефлекса требуется
- :-;тарное взаимодействие между мышцами рук и ног, а также между мышцами туловища.
. -.-ксы спинного мозга не исчерпываются рефлексами, ведущими к сокращению попереч-
?ой мускулатуры (скелетной мускулатуры). В рогах спинного мозга залегают первые
j :м. ниже) симпатической и парасимпатической нервных систем, поэтому сосудистые
: рефлексы мочеполовой системы, рефлексы, обеспечивающие деятельность потовых
:-:е осуществляются при участии спинного мозга.
м веществе спинного мозга большинство нейронов относятся к вставочным нейронам ::: онам), обеспечивающим пластичное взаимодействие афферентных и моторных ней-годаря этому между сегментами спинного мозга достигается взаимодействие, способ-: существлению простых двигательных программ.
.-:-:торная деятельность спинного мозга находится под контролем вышележащих отде-
. .:ьной нервной системы. Это наглядно демонстрируется тем фактом, что при лишении
:--::-:ого мозгас головным (например, в результате травмы) наступает «спинальный шок»,
:ное нарушение рефлекторных функций спинного мозга. «Спинальный шок» вызыва-
г. эляризацией мембран мотонейронов спинного мозга (после прекращения активиру-
1 :.^ния вышележащих отделов головного мозга), вследствие чего афферентация, прихо-
;цепторов, не может вызвать возбуждение мотонейронов, и рефлексы спинного мозга
:заются.
:: с этим следует особо подчеркнуть, что нервная регуляция всех без исключения функ-ческого организма всегда обеспечивается целым рядом динамически связанных мозго-р. находящихся на разных уровнях нервной системы (рис. 4.12). Возьмем для приме-_ дыхание. На уровне спинного мозга обеспечиваются движения, расширяющие грудку, без чего невозможен вдох; на уровне продолговатого мозга расположен вегетативный
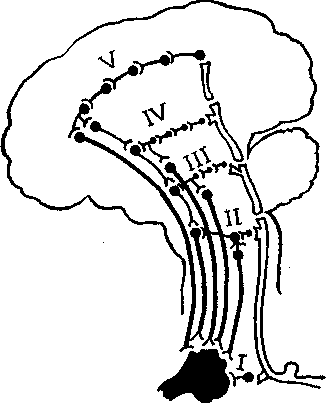
центр дыхательной деятельности; на уровне среднего мозга обеспечивается ритм, адекватность дыхательных движений вегетативной настроенности; примерно то же с участием аппарата эмоций осуществляется на уровне подкорковых образований (лимбической системы); на уровне коры головного мозга процесс дыхания увязывается со всей остальной деятельностью. Поражение каждого из отделов ведет к тем или иным расстройствам дыхания, вплоть до летального исхода вследствие паралича дыхания (при повреждении стволовых звеньев этой многоуровневой системы).
Рис. 4.12. Многоэтажное
(по вертикали) обеспечение функций
организма:
I — спинной мозг; II — ствол мозга; III —
гипоталамус; IV — подкорка; V — кора
головного мозга.
Проводниковая функция спинного мозга обеспечивается тем, что пучки нервных волокон (белое вещество) соединяют между собой рецепторы с эффекторами (исполнительными органами), головной мозг со спинным, а также сегменты спинного мозга между собой. В спинном мозге проходят восходящие пути, которые несут импульсы к головному мозгу, и ни-ходящие пути, по которым импульсация идет в обратном направлении, т. е. от головного мозга к спинному.
По восходящим проводящим путям возбуждение, возникшее в рецепторах конечностей, туловища, шеи, внутренних органов, идет по спинномозговым нервам в задние корешки, затем проводится в продолговатый мозг, а оттуда достигает коры больших полушарий.
Нисходящие проводящие пути несут импульсацию от головного мозга к мотонейронам спинного мозга, а от них через спинномозговые нервы передают импульсацию на эффекторы.
Итак, мы уже знаем, что функции нервной системы в целом и спинного мозга в частности координируются высшим отделом ЦНС — головным мозгом.
Головной мозг (рис. 4.13) располагается в полости черепа. В нем различают пять отделов, а именно: продолговатый мозг, задний мозг, средний мозг, промежуточный мозг и конечный мозг. Средний вес головного мозга человека составляет 1300—1600 г.
Так же как и спинной мозг, головной мозг покрыт оболочками (твердой, мягкой и паутинной). К ткани мозга непосредственно примыкает мягкая оболочка, в которой много кровеносных
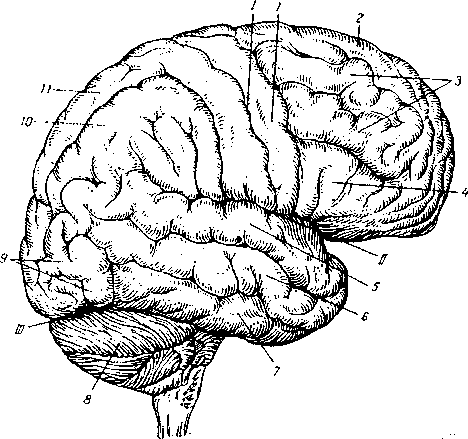
I — центральная борозда;
II — латеральная борозда;
III — поперечная щель мозга;
— предцентральная извилина;
— верхняя лобная извилина;
— средняя лобная извилина;
— нижняя лобная извилина;
— верхняя височная извилина;
— средняя височная извилина;
— нижняя височная извилина;
— мозжечок;
— затылочная доля;
10 — нижняя теменная долька; 11- верхняя теменная долька.
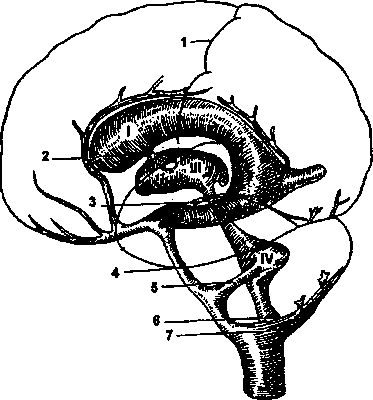
— парные боковые желудочки; III — третий желудочек; IV — четвертый желудочек; 1 — центральная (роландова) борозда;
— соединение между боковыми желудочками и III желудоч ком;.? — сильвиев водопровод, соединяющий III и IV желудоч ки: 4 — цистерна мозга; 5 — отверстие, соединяющее IV желудо чек с субарахноидальным пространством; 6 — отверстие, соеди няющее IV желудочек с мозжечково-мозговой цистерной; 7 —
мозжечково-мозговая цистерна.
с осудов,
питающих мозг. Между паутинной и мягкой
оболочками находится
пространство (субарахноидальное),
заполненное спинномозговой
жидкостью, которая, как уже было отмечено
выше, предохраняет мозг
от сотрясений, а также способствует
обмену между кровью и мозго-зой
тканью. Помимо субарахноидального
пространства, спинномозговая
жидкость содержится в четырех желудочках
головного мозга — двух боковых,
III
и IV
(рис 4.14). Мозговая жидкость продуцируется
сосудистыми сплетениями желудочков.
осудов,
питающих мозг. Между паутинной и мягкой
оболочками находится
пространство (субарахноидальное),
заполненное спинномозговой
жидкостью, которая, как уже было отмечено
выше, предохраняет мозг
от сотрясений, а также способствует
обмену между кровью и мозго-зой
тканью. Помимо субарахноидального
пространства, спинномозговая
жидкость содержится в четырех желудочках
головного мозга — двух боковых,
III
и IV
(рис 4.14). Мозговая жидкость продуцируется
сосудистыми сплетениями желудочков.
-4
Рис. 4.15. Продолговатый мозг (вид сзади):
1 — мост; 2 — передняя срединная щель; 3 — передние наружные дугообразные волокна (часть мостомоз-жечкового пути); 4 — перекрест пирамид; 5 — боковой канатик; 6 — олива
Нижним отделом головного мозга является продолговатый мозг, который рассматривается как непосредственное продолжение спинного мозга (рис. 4.15). Продолговатый мозг осуществляет рефлекторную и "роводниковую функции. По чувствительным волокнам черепномозго-: ;:х нервов (см. ниже) он получает импульсы от кожи, слизистых оболо--.::-i п органов головы, а также от рецепторов гортани, трахеи, внутренних органов грудной клетки (сердце, легкие), пищеварительной системы. Через продолговатый мозг осуществляются многие простые и слож-1:е рефлексы. Например: 1) защитные — кашель, чиханье, рвота, слезо-~деление, мигание; 2) пищевые — сосание, глотание, отделение пище-: - •- отельного сока; 3) сердечно-сосудистые, регулирующие деятельность :;ца и кровеносных сосудов. В продолговатом мозге находятся также .:. матически регулируемый дыхательный центр, обеспечивающий вен-•::~яцию легких и вестибулярные ядра, участвующие в осуществлении : лексов позы и в перераспределении тонуса мышц. Мас-
: :-:ое повреждение продолговатого мозга приводит к летальному исходу вследствие прекраще-г_ :ердечной деятельности и дыхания.
Кроме того, через продолговатый мозг проходят пути, соединяющие двусторонней связью tv головного мозга, промежуточный и средний мозг, мозжечок и спинной мозг. Задний мозг включает в себя мост и мозжечок (рис. 4.16).
Задний мозг является жизненно важным отделом нервной системы, где происходит замыка-
-:не дуг целого ряда соматических и вегетативных рефлексов. При участии ядер заднего мозга
; глествляются рефлексы, связанные с жеванием и глотанием. С функцией пищеварительного
: лкта связаны многие вегетативные рефлексы заднего мозга (например рефлекторная регуля-
.'.'..-. секреции слюнных желез).
Мозжечок входит в систему регуляции движений и выполняет следующие важные функции: 1) регуляцию позы и мышечного тонуса; 2) сенсомоторную (чувственно-двигательную) координацию позы и целенаправленных движений; 3) координацию быстрых целенаправленных движений, осуществляемых по команде из коры больших полушарий.
Рис. 4.16. Задний мозг:
1 — мозжечок; 2 — мостомозжеч-ковый треугольник; 3 — бульбар-номостовая борозда; 4 — базилляр-ная борозда; 5 — средняя мозжечковая ножка; б — мост
Главные функции мозжечка определяют характер патологических симптомов при нарушении его деятельности. При общем органическом поражении мозжечка наблюдаются многочисленные симптомы, связанные с расстройством координации движений и мышечного тонуса. У животных после удаления мозжечка (рис. 4.17) наблюдается начальное повышение тонуса мышц-разгибателей. Движения их плохо скоординированы, размашистые, резкие, они не способны поддерживать соответствующую позу.
Средний мозг расположен впереди (варолиева) моста. К среднему мозгу относятся четверохолмие и ножки мозга (рис. 4.18). У человека верхние холмики четверохолмия и латеральные коленчатые тела выполняют функцию подкорковых зрительных центров, а нижние холмики четверохолмия и медиальные коленчатые тела являются подкорковыми слуховыми центрами.

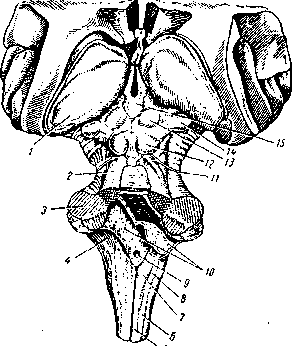
1 — таламус (зрительный бугор); 2 — верхняя мозжечковая ножка; 3 — средняя мозжечковая ножка; 4 — нижняя мозжечковая ножка; 5 — тонкий пучок (продолговатый мозг); 6 — клиновидный пучок; 7 — бугорок тонкого ядра; 8 — бугорок клиновидного ядра; 9 — срединное отверстие IV желудочка; 10 — сосудистое сплетение IV желудочка; 11 — блоковидный нерв; 12 — нижнее четверохолмие; 13 — верхнее четверохолмие; 14 — медиальное коленчатое тело; 75—эпифиз
Функциональное значение среднего мозга заключается в том, что здесь находятся: подкорковые центры зрения и слуха; ядра черепномозговых нервов, обеспечивающих иннервацию мышц глазного яблока; двигательные ядра (компактные скопления нейронов) эктрапирамидной системы, которые обеспечивают сокращения мышц тела во время автоматических движений9. Помимо этого, через средний мозг проходят восходящие (чувствительные) и нисходящие (двигательные) проводящие пути. При участии структур среднего мозга осуществляются ориентировочные слуховые рефлексы.
Средний мозг является также местом расположения вегетативных центров и ретикулярной (сетевидной) формации. Ретикулярная формация (рис. 4.19) находится во внутренних областях продолговатого мозга, моста и, как уже было отмечено, в области среднего мозга. Ретикулярная формация состоит из огромного числа нейронов различной формы и размеров. Отростки нервных клеток ретикулярной формации идут как в восходящем, так и в нисходящем направлениях.
Талимус
•Зрительный
рецептор
Слуховой
рецептар
СпшнамозгоЙой
путь Стваловой
пути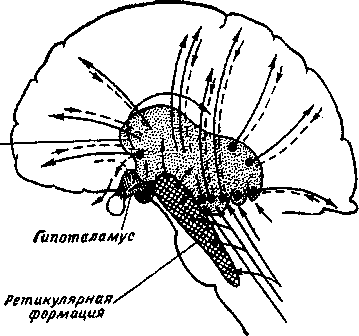
В свою очередь ее нервные клетки получают импульсацию также как от восходящих, так и от нисходящих путей, т. е. от самых разных мозговых структур. Кроме того, нейроны ретикулярной формации образуют более четырех десятков ядер. Нисходящие ретикулоспинальные воздействия оказывают тоническое влияние на рефлексы спинного мозга, регулируя движения, позу и вегетативные рефлексы. Так, повреждение среднего мозга у животных вызывает нарушение тонуса мышц, выражающееся в резком повышении тонуса мышц-разгибателей (так называемая «деце-ребрационная ригидность»). Внешне этот эффект проявляется в том, что животное, поставленное на лапы, так и остается в стоячем положении, будучи не в силах согнуть их, т. к. не происходит сгибания лап в суставах. Децеребрационная ригидность обусловлена тем, что в результате перерезки ствола от продолговатого и спинного мозга отделяются ретикулярная формация и красные ядра.
Восходящие ретикулокортикальные воздействия поддерживают тонус коры, регулируют уровень бодрствования, определяют уровень концентрации внимания, а также поддерживают проявление ориентировочных рефлексов. Нейроны ретикулярной формации являются крайне чувствительными к различного рода гуморальным агентам (от греч. humor — жидкость), что является важным в плане объяснения механизма влияния гуморальных факторов и эндокринной гнстемы на высший отдел нервной системы — кору головного мозга.
Самым крупным из ядер, которые видны на разрезе среднего мозга, являются красное ядро и черная Ч тянция. Эти ядра относятся к экстрапирамидной системе.
С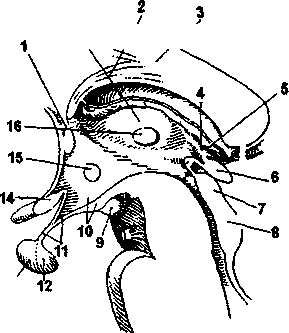 ледующим
вышележащим отделом головного мозга
является промежуточный мозг (рис. 4.20).
В состав промежуточного
ледующим
вышележащим отделом головного мозга
является промежуточный мозг (рис. 4.20).
В состав промежуточного
мозга входят таламус, гипоталамус и эпиталамус.
Таламус (зрительный бугор) представляет собой парное образование, расположенное по сторонам III желудочка. Таламус состоит из серого вещества (тел нейронов), в котором насчитывается большое количество ядер.
13
Рис.
4.20. Продольный разрез мозгового
ствола10:
1
— передняя
спайка; 2 — свод; 3
— мозолистое
тело; 4
— борозда
поводка; 5 — спайка поводка; 6
—
эпифиз; 7
— верхнее четверохолмие; 8
— нижнее
четверохолмие; 9
— мамиллярное
тело;
10
— серый
бугор; 11
— воронка;
12
— нейрогипофиз;
13
— адено-гипофиз;
14
— зрительный
перекрест; 15
— гипоталамус;
16
— межталами-ческое
сращение; 17
— таламус
К гипоталамусу относят зрительный перекрест, зрительный тракт, мамиллярные (сосцевидные тела), серый бугор, переходящий внизу в воронку, которая далее соединяется с гипофизом.
Гипоталамус с гипофизом образуют единый функциональный комплекс, в котором гипоталамус играет регулирующую роль, а гипофиз эффекторную (исполнительную). В гипоталамусе осуществляется взаимодействие нервной и эндокринной11 систем организма человека.
Гипоталамус является высшим подкорковым центром нервной (вегетативной — см. ниже) системы. За счет мобилизации вегетативных и эндокринных механизмов он отвечает за регуляцию внутренней среды организма (поддержание гомеостаза). Некоторые структуры гипоталамуса входят в лимбическую систему (высший подкорковый центр, имеющий отношение к эмоци-онально-мотивационной сфере). Поэтому активность гипоталамуса коррелирует с проявлениями эмоций и мотивации. Перечислим основные функции гипоталамуса:
1) Гипоталамус участвует в регуляции температуры тела. В гипоталамусе имеются нейроны, чувствительные к изменению температуры в этой области мозга. Если специально поднять температуру в этой области, то у экспериментального животного наблюдаются учащение дыхания и расширение периферических кровеносных сосудов, что приводит к повышенной теплоотдаче. При охлаждении определенной области гипоталамуса наблюдается противоположный эффект — урежение дыхания, дрожь и сужение сосудистой периферии. Повреждение определенных областей гипоталамуса приводит к нарушению процессов терморегуляции организма.
Гипоталамус участвует в регуляции дыхания.
Гипоталамус участвует в регуляции деятельности сердечно-сосудистой системы.
Гипоталамус участвует в контроле водного баланса в организме. Так, стимуляция опреде ленной области («центра жажды») гипоталамуса вызывает увеличенное поглощение воды, а по вреждение соответствующей области приводит к отказу от потребления воды.
Гипоталамус участвует в регуляции пищевого поведения. В гипоталамусе находятся два взаимодействующих центра, определяющих уровень пищевой мотивации, — «центр голода» и «центр насыщения». Повреждение этих центров (рис. 4.21) приводит к нарушению регуляции пищевого поведения.
Таламус
Разрушение
латерального
ядра вызывает
потерю аппетита
Разрушение
венгро-медиального ядра вызывает
ярость и
чрезмерный аппетит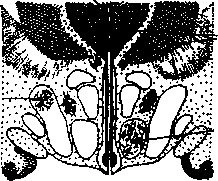
Рис. 4.21. Влияние повреждения «центра голода» и «центра насыщения»
Гипоталамус имеет отношение к регуляции половой активности. Стимуляция определен ных областей гипоталамуса вызывает у экспериментальных животных внешние поведенческие признаки «готовности к спариванию».
Гипоталамус является одной из структур, участвующих в генерации отрицательных аф фектов (страха и ярости), поскольку стимуляция определенных областей гипоталамуса вызыва ет у подопытных животных реакции, соответствующие по своим внешним проявлениям указан ным аффектам.
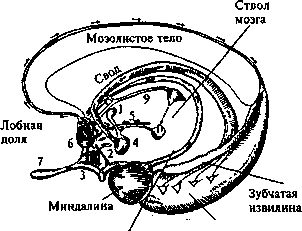
Как уже было отмечено выше, некоторые структуры гипоталамуса относятся к лимбической системе (рис. 4.22), высшему подкорковому центру, активность отделов которого коррелирует с внешними проявлениями аффективно-мотивационных процессов, а также с процессами памяти и (предположительно) ориентировочного рефлекса.
В определенных областях лимбической системы (например медиальный пучок переднего мозга) имеются также «центры удовольствия».
Опыты Джеймса Олдса12, проведенные на крысах при помощи вживленных в лимбические структуры мозга микроэлектродов, показали связь этих отделов с эмоциональными переживаниями. Крысы, научившись передними лапами нажимать на педали, от которых провода шли к вживленным в их мозг электродам, могли раздражать различные участки своей лимбической системы. Оказалось, что при раздражении определенных участков крысы продолжали безудержно (до 4000—5000 раз в час) нажимать на педаль.
Связь лимбических структур головного мозга с эмоциональной сферой подтвердили также опыты испанца Хосе Дельгадо13 с электрическим раздражением на расстоянии все тех же подкор-
1
Поясная извилина
Свод
Гипоталамус;
дугообразное
ядро
паравентрикулярное
ядро
/
Гипофи /АКТГ
Кортизол Надпочечник
Надпочечник
Гиппокамп
Гиппокамп Терминальные
'олубое пятно волокна
Миндалина Схема связей лимбической системы.
1 — переднее таламическое ядро; 2 — диагональная связка (Брока); 3 — латеральная обонятельная полоска; 4 — мамил-лярное тело; 5 — медиальный пучок переднего мозга; 6 — медиальная обонятельная полоска; 7 — обонятельная луковица; 8 — перегородка; 9 — медуллярные волокна; 10 — обонятельный бугорок.
Рис. 4.22. Лимбическая система
головного мозга
ковых структур, когда разъяренный бык, мчащийся к тореадору, превращался в испуганное, ретирующееся животное.
К структурам лимбической системы, имеющим какое-то отношение к сфере аффектов, принадлежит, по-видимому, и миндалина. Об этом свидетельствует тот факт, что после ее двустороннего разрушения у обезьян пропадает чувство страха и ярости. Так, обезьяны начинают безбоязненно брать в рот змей, которых они обычно панически боятся, а доминантные самцы, занимавшие до разрушения миндалины высокое положение в стаде, начинают занимать подчиненное положение.
Еще одна структура лимбической системы — гиппокамп каким-то образом связана с процессами памяти и, предположительно, с механизмом ориентировочного рефлекса.
Повреждение гиппокампа приводит к характерным грубым расстройствам мнемической функции и утрате способности к обучению. Впервые это расстройство алкогольного генеза (синдром Корсакова) было описано русским психиатром С. С. Корсаковым еще в 19 веке. Синдром Корсакова выражается в том, что больной сохраняет воспоминания об отдаленных периодах своей жизни, но совершенно не помнит о том, что произошло с ним несколько минут назад. Если, например, лечащий врач выходит из палаты на пять минут, то при повторном посещении палаты больной не узнает его. Патологоанатомическое обследование таких больных обнаруживало у них дегенеративные повреждения гиппокампа14.
Нейроны гиппокампа вместе с нейронами коры головного мозга, согласно гипотезе академика Е. Н. Соколова15, обеспечивает механизм ориентировочного рефлекса, являющегося основой ориентировочно-исследовательской деятельности в целом.
Ориентировочный рефлекс был определен великим русским физиологом И. П. Павловым как рефлекс «Что такое?». Главная функция ориентировочного рефлекса заключается в настройке (повышении чувствительности) анализаторов16 для наилучшего восприятия действующих раздражителей и выделения из них наиболее значимых в данный момент времени17.
для восприятия сигналов из внешней и внутренней среды, преобразования их в нервный импульс и проведения его (нервного импульса, в котором закодирован стимул) к высшим отделам мозга для последующего анализа. В состав анализатора входят периферический прибор, преобразующий внешнее воздействие в нервный код;В плане внешних поведенческих проявлений начальная фаза ориентировочного рефлекса на новый стимул выражается в прекращении текущей деятельности и в фиксации позы (стоп-реак-ция).
Затем на стадии общей активации организм приводится в состояние готовности к встрече с возможно значимым для него стимулом (воздействием). Внешне это проявляется в повышении тонуса скелетных мышц, повороте головы и глаз в направлении раздражителя, а также в целом ряде вегетативных сдвигов, таких, как изменение сердечного и дыхательного ритмов, периферические сосудистые реакции и т. д. Имеет место и изменение кожно-гальванической реакции.
Наконец, организм переходит в режим (фазу) дифференцированного анализа стимула, для которой характерно быстрое угашение вегетативных компонентов рефлекса и снижение интенсивности (исчезновение) кожно-гальванической реакции при многократном повторении одного и того же константного стимула. Однако при изменении любого параметра стимула (его интенсивности, локализации в пространстве, длительности и т. д.) ориентировочный рефлекс восстанавливается, т. е. наблюдается эффект «растормаживания».
На основании экспериментального исследования динамики «угашения — растормаживания» Е. Н. Соколов предположил, что повторение константного стимула приводит к построению в гиппокампе «нервной модели стимула», точно отражающей параметры повторяющегося стимула. Согласно представлениям Е. Н. Соколова, при предъявлении того или иного стимула впервые нейроны-детекторы специфических физических признаков стимула возбуждают огромное количество нейронов «новизны» гиппокампа. Нейроны «новизны» связаны, в свою очередь, с нервным центром ориентировочного рефлекса, расположенным в области ретикулярной формации, который и приводит к запуску сложного рефлекторного процесса. При повторных предъявлениях того же самого стимула ответ нейронов «новизны» постепенно становится слабее и наконец угасает совсем. Это обусловлено тем, что происходит активация нейронов «тождества», которые оказывают тормозное влияние на нейроны «новизны». При предъявлении нового (по какому-либо физическому параметру) стимула активируется новая «матрица» нейронов «новизны» и ориентировочный рефлекс возникает снова. При этом выраженность компонентов ориентировочно увеличивается пропорционально величине рассогласования (различия) между параметрами нового стимула и характеристиками стимула, который соответствует «нервной модели стимула».
Модель Е. Н. Соколова является одной из самых известных теорий ориентировочного рефлекса (внимания), однако она не в состоянии объяснить возникновение ориентировочных реакций на стимул, который не является ни новым, ни отличным (например на вторую цифру «8» в цифровом ряду 123456788). Помимо этого ориентировочная реакция может возникать и в сложном ряду стимулов, свойства которых постоянно изменяются в сравнении с предшествующими так, что эту ситуацию никак нельзя отнести к случаям появления неожиданного стимула с точки зрения его физических параметров. Например ориентировочная реакция, которая возникает на цифру «5» в ряду РКЛОБНЖ5. Указанные феномены могут быть объяснены корковыми влияниями на ориентировочный центр ретикулярной формации, осуществляемыми по эфферентным кортикоретикулярным проводящим путям. Таким образом, под сомнением оказывается базовый
постулат гипотезы Е. Н. Соколова о том, что именно физические изменения параметров стимуляции всегда приводят к возникновению ориентировочного рефлекса. В связи с этим А. Бернштейн высказал идею о том, что «хотя ориентировочная реакция и следует за изменением стимула, однако это скорее результат значения стимула на основании прошлого опыта, а не свойства стимула как такового. Согласно гипотезе «значимости» Бернштейна, корковые процессы интерпретации и усиления являются решающими...»18.
На наш взгляд, и тот и другой предложенные объяснительные принципы механизма инициации ориентировочной реакции, по-видимому, существуют в действительности. При этом механизм «значимости» стимула является в реальной (а не лабораторной) обстановке доминирующим, в то время как «нервная модель стимула» Е. Н. Соколова может быть использована как нейрофизиологический коррелят непроизвольного (пассивного) внимания.
Итак, мы рассмотрели основные функциональные особенности структур промежуточного мозга. Перейдем теперь к описанию структуры и функций конечного мозга.
ю
Рис. 4.23. Головной мозг на разрезе:
К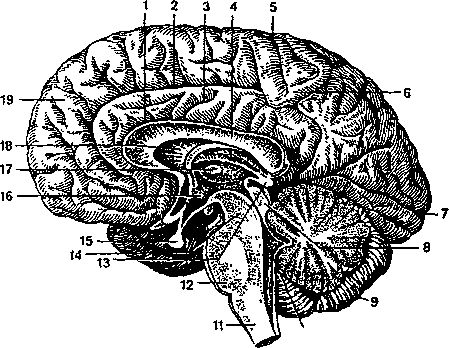 онечный
мозг состоит из двух больших полушарий,
разделенных продольной щелью и
соединенных в ней с помощью мозолистого
тела,
а также передней и задней комиссур (вид
сбоку
на рис. 4.13, вид на поперечном разрезе
на
рис. 4.23). Общая поверхность коры полушарий
(серого вещества) составляет 2000— 2500
см2,
толщина ее — 2,5—3 мм. Кора включает
более 14 млрд нервных клеток. Кора
образует
складки (извилины), разделенные
бороздами.
Борозды делят поверхность полушарий
на доли; в каждом полушарии различают
четыре доли: лобную, теменную, височную
и затылочную.
Новая кора состоит из 6 слоев нервных
клеток, объединенных в колонки.
онечный
мозг состоит из двух больших полушарий,
разделенных продольной щелью и
соединенных в ней с помощью мозолистого
тела,
а также передней и задней комиссур (вид
сбоку
на рис. 4.13, вид на поперечном разрезе
на
рис. 4.23). Общая поверхность коры полушарий
(серого вещества) составляет 2000— 2500
см2,
толщина ее — 2,5—3 мм. Кора включает
более 14 млрд нервных клеток. Кора
образует
складки (извилины), разделенные
бороздами.
Борозды делят поверхность полушарий
на доли; в каждом полушарии различают
четыре доли: лобную, теменную, височную
и затылочную.
Новая кора состоит из 6 слоев нервных
клеток, объединенных в колонки.
1 — борозда мозолистого тела; 2 — поясная борозда; 3 — поясная извилина; 4 — мозолистое тело; 5 — центральная борозда; 6 — парацентральная долька; 7 — шпорная борозда; 8 — средний мозг; 9 — мозжечок; 10 — IV желудочек; 11 — продолговатый мозг; 12 — мост; 13 — шишковидное тело (эпифиз); 14 — ножки мозга; 15 — гипофиз; 16, 17 — промежуточный мозг; 18 — прозрачная перегородка; 19 — верхняя лобная извилина
Комиссуральные пути соединяют между собой идентичные участки правого и левого полушарий.
Кора является высшим отделом головного мозга. В кору (корковые концы анализаторов) приходит закодированная в нервных кодах информация, воспринятая органами чувств из внешней и внутренней среды (рис 4.24). При участии коры осуществляются все высшие психические функции человека, такие как восприятие, память, внимание, мышление, формирование мотивации и т. д.
Активность коры головного мозга неразрывно (однако неизвестно каким образом) связана с сознанием (картиной мира) — высшим уровнем психического отражения. Речь, как специфически человеческий феномен, также неразрывно связана с определенными областями коры.
Вполне справедливо будет сравнить нервную систему с совершенным компьютером (кибернетической машинной установкой), не забывая, понятно, о том, что мозг человека совершен-
Помимо серого вещества, в коре головного мозга имеется белое вещество, состоящее из проводящих путей, которые делятся на ассоциативные (объединяют разные участки коры внутри одного полушария) и проекционные (связывают полушария с нижележащими отделами мозга).
ж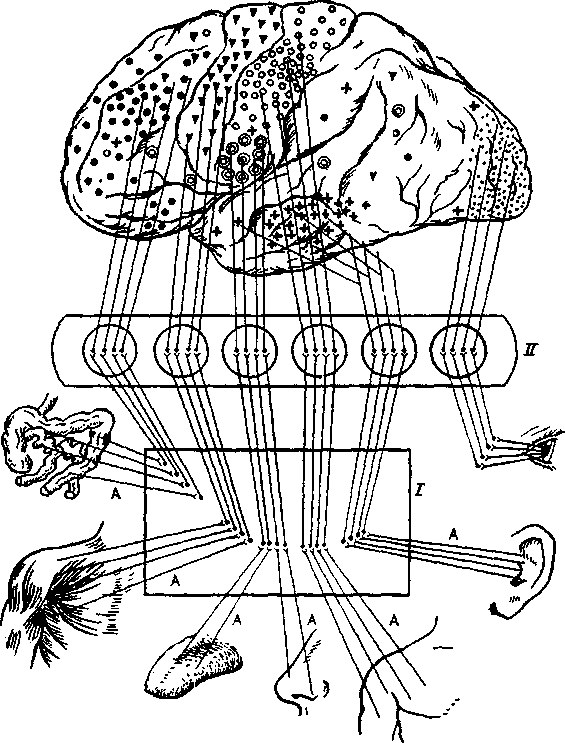
W.V.V ЗРИТЕЛЬНЫЙ АНАЛИЗАТОР (КОРКОВЫЙ ОТДЕЛ) 4.НЧ. + ++ СЛУХОВОЙ >' »>
0°о°о°о КОЖНЫЙ 1> П
ВКУСОВОЙ И ОБОНЯТЕЛЬНЫЙ и I»
ДВИГАТЕЛЬНЫЙ » »»
•••••• ИНТЕРОЦЕПТИВНЫЙ ч П
Рис. 4.24. Корковые анализаторы:
I — органы чувств (рецептивные поля); II — проводящие пути и промежуточные подкорковые центры;
III — корковые концы анализаторов.
нее любой кибернетической машины, поскольку нервная система, да и весь организм человека, как система биологическая, обладает огромными приспособительными и компенсаторными возможностями с многочисленными механизмами саморегуляции и активной деятельности. Условиями кибернетического устройства являются передача, переработка и хранение информации с последующим ее воспроизведением. Передача информации идет в закодированном виде.
Раскодирование
информации каким-то образом происходит
в коре мозга, что и дало право 11.
П. Павлову образно сравнить кору
головного мозга с сигнализационной
доской, на которой то там,
то
здесь вспыхивают электрические
лампочки.е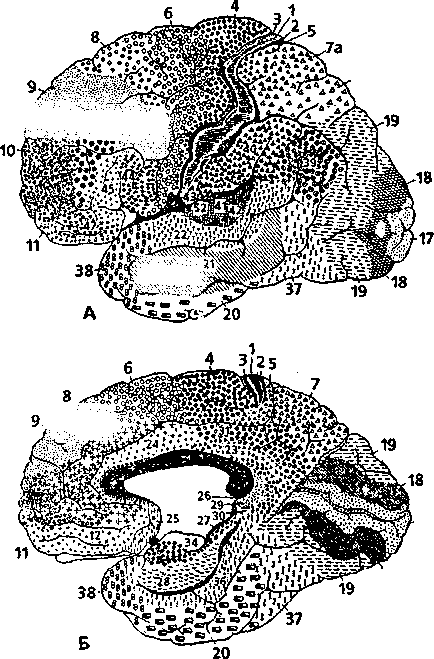 равнозначность
различных отделов коры мозга,
наличие в ней большого числа высших
отделов
анализаторов дали основание для
анато-мо-функционального
подразделения коры на отдельные
цитоархитектонические поля, в которых
структура
определяет в какой-то мере функцию.
Так,
широкую известность получила
цитоархи-тектоническая карта Бродмана,
опубликованная в
1925 году (рис. 4.25).
равнозначность
различных отделов коры мозга,
наличие в ней большого числа высших
отделов
анализаторов дали основание для
анато-мо-функционального
подразделения коры на отдельные
цитоархитектонические поля, в которых
структура
определяет в какой-то мере функцию.
Так,
широкую известность получила
цитоархи-тектоническая карта Бродмана,
опубликованная в
1925 году (рис. 4.25).
Достаточно хорошо известно, в каких корковых полях происходит анализ и синтез той или иной информации (представленной в кодах нейронной импульсации) с трансформацией ее (абсолютно неясным образом) в ощущения.
10
17
18
Первичная зона сенсорного (чувствительного) анализатора занимает постцентральную извилину (3-е поле). Проекции тела расположены в постцентральной извилине в следующем порядке: в верхних частях расположены проекции от нижних конечностей, в средней области — от верхних, а в нижней области находятся проекции от лица и внутренних органов (рис. 4.26).
Величина частей тела на рисунке соответствует размерам сенсорного представительства в коре.
Рис. 4.25. Цитоархитектонические поля по Бродману:
А — вид коры больших полушарий сбоку; Б — вид коры больших полушарии на срединном разрезе.
При локальных повреждениях постцентральной извилины наблюдается потеря чувствительности в соответствующей области тела.
Первичный корковый конец двигательного анализатора (рис. 4.27) расположен в прецент-ральной извилине (4-е поле).
Величина частей тела на рисунке соответствует размерам двигательного представительства в коре.

Рис. 4.26. Расположение в сенсорной зоне коры больших полушарий проекций частей тела (по У. Пенфильду):
1 — половые органы; 2 — пальцы; 3 — ступня; 4 — голень; 5 — бедро; 6 — туловище; 7 — шея; 8 — голова; 9 — плечо; 10 — локтевой су став; 77— локоть; 72 — предплечье; 13 — запястье; 14 — кисть; 15 — мизинец; 76 — безымянный палец; 77 — средний палец; 18 — указательный палец; 19 — большой палец; 20 — глаз; 21 — нос; 22 — лицо; 23 — верхняя губа; 24 — зубы; 25 — нижняя губа; 26 — зубы, десны, челюсть; 27 — язык; 28 — глотка; 29 — внутренние
органыГлава 4
П ервичные
проекционные зоны зрительного анализатора
находятся в затылочной доле (17-е поле).
Нейроны-детекторы этой
области реагируют лишь на отдельные
специфические параметры
стимулов (цвет, характер линий, направление
движения
и т. д.).
ервичные
проекционные зоны зрительного анализатора
находятся в затылочной доле (17-е поле).
Нейроны-детекторы этой
области реагируют лишь на отдельные
специфические параметры
стимулов (цвет, характер линий, направление
движения
и т. д.).
Первичный слуховой анализатор расположен в височной доле (41-е поле).
L
Рис. 4.27. Расположение двигательных участков в моторной зоне больших полушарий (по У. Пенфильду):
1 — пальцы; 2 — лодыжка; 3 — колено; 4 — бедро; 5 — туловище; б — бедро; 7 — локоть; 8 — запястье; 9 — кисть; 10 — мизинец; 11 — безымянный палец; 12 — средний палец; 13 — указательный палец; 14 — большой палец; 15 — шея; 16 — бровь; 17 — веко и глазное яблоко; 18 — лицо; 19 — губы; 20 — челюсть; 21 — язык; 22 — гортань
Для всех анализаторов характерно существование так называемых вторичных зон, при органическом поражении которых наступают более выраженные и комплексные нарушения в работе соответствующего анализатора. Так, повреждение вторичной зоны сенсорного анализатора (1, 2, 5, 7-е поля и частично 40-е поле) человек не в состоянии определить наощупь предъявляемые ему предметы, не смотря на то, что чувственная основа ощущений (первичная зона) остается сохранной. При этом нарушается также «схема тела», т. е. возникает расстройство в восприятии расположения частей собственного тела по отношению друг к другу и т. д. При повреждении вторичных зон моторной области (6-е и 8-е поля) не возникают параличи, а имеет место расстройство сложных двигательных навыков, таких, например, как почерк или автоматизированные движения спортсмена.
При органическом поражении вторичных зон зрительного анализатора (18-е и 19-е поля) слепота не наступает, однако чело-зек утрачивает способность объединять отдельные видимые признаки в единый образ.
При массивном поражении вторичных проекционных зон слухового анализатора (41,42-е и 22-е поля) больной утрачивает способность определять значение предметных звуков (скрип двери, шум воды), а также теряет способность понимать обращенную к нему устную речь.
Итак, в строении любого анализатора выделяют первичные и вторичные проекционные зоны. В них осуществляется прием, переработка и хранение информации. При этом вторичные зоны надстраиваются над первичными.
В осуществлении процессов высшей психической регуляции принимают участие19 так назы-
; ;-гМые третичные или ассоциативные зоны коры. Эти зоны обеспечивают скоординированную
:.. :оту отдельных анализаторов, обеспечивая тем самым предпосылки для создания всесторонне-
": психического отражения действительности. Различают две ассоциативные зоны. Первая об-
. _ ~ъ расположена на стыке корковых представительств зрительного (затылочного), слухового
. ;;:очного) и сенсорного (теменного) анализаторов, а именно в 7, 39-е и 40-м полях теменной
"лети, в 21-м поле височной области и в 37-м поле височно-затылочной области. Эта область
:; ~лется специфически человеческим образованием. Она имеет отношение к трансформации чув-
~: гнного восприятия предметов и явлений в их отражение в рамках словесно-логического мыш-
'" При органическом повреждении этих областей человек утрачивает способность к узнава-: : дметов и явлений, нарушаются процессы интеллектуальной переработки, хранения и вос-
: введения информации. При этом повреждение теменной ассоциативной зоны никак не сказы-
;.~. на работе отдельно взятых анализаторов, однако больные утрачивают способность к ориен-
::: в пространстве (например, одеваясь, они не могут попасть рукой в нужный рукав и т. д.).
имеем в виду, что при поражении этой структуры или области наблюдаются более или менее выраженные В расстройства соответствующей психической функции.
Другая ассоциативная зона коры больших полушарий головного мозга расположена в лобных долях спереди от моторной коры. При поражении корковых лобных отделов у человека нарушается целенаправленная произвольная (сознательно-волевая) регуляция высших психических функций (мышления и памяти) и деятельности. Больные не в состоянии следовать какой-либо линии поведения, они чрезвычайно отвлекаемы, постоянно перескакивают от одного действия к другому и т. д. (например, больной не способен самостоятельно зашнуровать ботинок, хотя прекрасно понимает данную ему инструкцию). При попытках решить какую-либо задачу больные с поражением премоторной коры осуществляют лишь случайные действия и при этом совершенно не замечают их бессмысленности. Также у таких больных утрачивается способность к целостному восприятию действительности. Так, при предъявлении сюжетной картинки они не способны сопоставить детали картинки друг с другом. Например, больному показывают картинку, на которой изображены мальчик, провалившийся в прорубь, и люди, бегущие к нему на выручку. На берегу реки стоит предупреждающий знак с надписью «Осторожно». Больной не способен проанализировать сюжет картинки. Увидев табличку с надписью «Осторожно», он говорит — «Ток высокого напряжения», а увидев милиционера, бегущего к мальчику на выручку, больной заключает — «Война»20.
После поражения лобной коры имеют место и эмоциональные расстройства. Больные становятся апатичными, безразличными к происходящему вокруг них, эмоции приобретают стертый характер. В то же время больные могут неожиданно проявлять вспышки немотивированной агрессии.
Рис. 4.28. Локализация центров речи
в левом доминантном (у правшей)
полушарии:
1 — область лицевой мускулатуры; 2 — дугообразный пучок; 3 — зрительная кора; 4 — угловая извилина; 5 — зона Вернике; 6 — сильвиева щель; 7 — зона Брока
Кора больших полушарий головного мозга также связана с процессами понимания и продуцирования речи (рис. 4.25 и 4.28). В задней трети нижней лобной извилины левого полушария (у правшей) находится так называемая (по имени французского врача, открывшего ее в 1861 году) зона Брока (поля 44-е и 45-е). При полном разрушении этой зоны больные не могут произнести ни одного слова. При поражении областей, расположенных рядом с зоной Брока (9,10,46-е поля), у больного возникают затруднения, связанные со сложным (развернутым) речевым высказыванием. В задних отделах височной доли расположена так называемая зона Вернике (названа по имени немецкого психиатра К. Вернике, открывшего ее в 1874 г.), относящаяся к вторичным зонам слухового анализатора (41,42,22-е поля). При повреждении этой зоны человек не понимает обращенной к нему речи.
Итак, мы описали основные структуры и функции центральной нервной системы. Обратимся теперь к описанию периферической нервной системы.
Периферическая нервная система подразделяется на соматическую нервную систему и на вегетативную21, строение и функции которой мы рассмотрим сейчас более подробно, поскольку
Гипоталамическая
(диэнцефальная
область)
долокна
Глаз
Слюнные
железы
Сосуды голова
Щитовидная
железа
Лёгкие
Сердце
Желудок
Печень
Поджелудочная
железа.
Надпочечник
По
ч к и
Тонкие
нитки Толстые
ниш ни
Мочевой
пузырь Половые
органы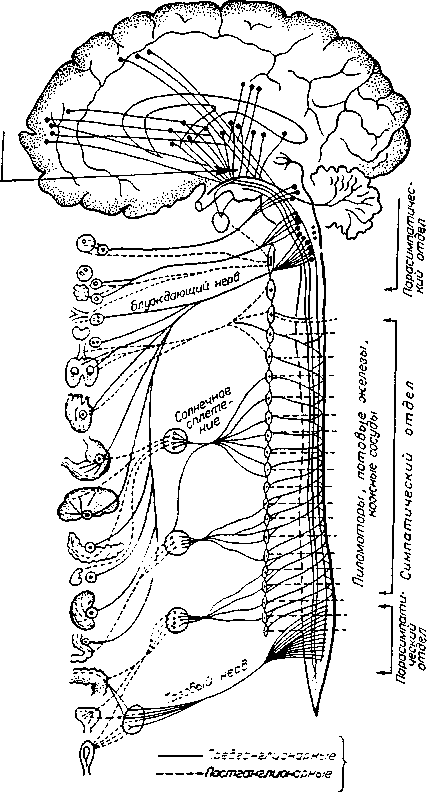
именно вегетативная (автономная) нервная система определяет (под воздействием психических факторов) динамику физиологических процессов, регистрируемых в ходе полиграфных проверок.
Вегетативная нервная система, обеспечивающая все внутреннее хозяйство организма, функционально тесно связана с центральной нервной системой (ствол мозга), в том числе и с корой головного мозга (лобная и теменная доли).
Вегетативная нервная система берет свое начало (рис. 4.29) в среднем, продолговатом мозгу, грудино-поясничнои и крестцовой части спинного мозга и делится на симпатическую и парасимпатическую нервную систему. Волокна, отходящие в центральной нервной системе от среднего и продолговатого мозга и из крестцового отдела спинного мозга, образуют парасимпатическую нервную систему. Та же часть вегетативной нервной системы, которая начинается в грудино-поясничном отделе, называется симпатической нервной системой.
Особенностью строения вегетативной нервной системы является то, что нервные волокна после выхода из центральной нервной системы оканчиваются, не доходя до иннервируемого ими органа. Однако они тут же вступают в связь с другой нервной клеткой, отросток которой уже доходит до органа. Таким образом, вегетативные нервы состоят из двух нейронов, между которыми находится синапс. В местах перерыва имеется большое скопление нейронов, которые образу-ют ганглии (узлы) вегетативной нервной системы. Нервные волокна, которые выходят из центральной нервной системы и оканчиваются в ганглиях, называются преганглионарными. Те же волокна, которые отходят от ганглия и доходят до определенного органа, получили название постганглионарных. Синаптические окончания преганглионарных волокон как симпатической, так и парасимпатической нервных систем выделяют трансмиттер ацетилхолин. В синаптических окончаниях постганглионарных волокон симпатической нервной системы выделяется медиатор норадреналин (оказывает возбуждающее воздействие), а постганлионарные волокна парасимпатической нервной системы выделяют в своих синаптических окончаниях медиатор ацетилхолин. Ганглии вегетативной нервной системы располагаются на разных расстояниях от центральной нервной системы. Имеются ганглии, находящиеся непосредственно у позвоночного столба; другая группа ганглиев располагается сравнительно далеко от позвоночника, примерно на одинаковом расстоянии между органом и позвоночным столбом; третья группа ганглиев находится непосредственно в самих органах. Ганглии симпатической системы располагаются около позвоночного столба и образуют цепочку справа и слева от него. Ганглии парасимпатической нервной системы располагаются внутри органов или рядом с ними. Волокна вегетативной нервной системы менее возбудимы, чем соматические, возбуждение по вегетативным волокнам проводится медленнее, а рефрактерный период более длительный. Преганглионарные волокна вегетативной нервной системы миелинизированные, в то время как постганглионарные лишены миелиновой оболочки. Особенности строения симпатической и парасимпатической нервной системы отражены в таблице 4.1.
Таблица 4.1 Различия между симпатической и парасимпатической нервными системами
Особенность или свойство |
Симпатическая система |
Парасимпатическая система |
Происхождение нервных волокон |
Выходят из грудного и поясничного отделов спинного мозга |
Выходят из ствола головного мозга и крестцового отдела спинного мозга |
Расположение |
Рядом со спинным мозгом |
В эффекторе (органе) или рядом с ним |
Длина волокон |
Короткие преганглионарные и длинные постганглионарные волокна |
Длинные преганглионарные и короткие постганглионарные волокна |
Число волокон |
Многочисленные постганглионарные волокна |
Немногочисленные постганглионарные волокна |
Распределение волокон |
Постганглионарные волокна иннервируют обширные области |
Постганглионарные волокна инвертируют ограниченные области |
Зона влияния |
Действие генерализованное |
Действие местное |
Медиатор |
В эффекторе освобождается норадреналин |
В эффекторе освобождается ацетилхолин |
Общие эффекты |
Усиливает ритмические формы активности. Снижает пороги чувствительности |
Снижается интенсивность обмена или не влияет на нее. Снижает ритмические формы активности. Восстанавливает пороги чувствительности до нормального уровня |
Суммарный эффект |
Возбуждающий |
Тормозящий |
В каких условиях активируется |
Доминирует во время опасности, стресса и активности. Контролирует реакции на стресс. |
Доминирует в покое. Контролирует обычные «повседневные» физиологические функции |
Парасимпатическая нервная система берет свое начало от среднего и продолговатого мозга и из крестцового отдела спинного мозга (рис. 4.29). От среднего мозга отходят парасимпатические волокна, входящие в состав глазодвигательного22 нерва (одного из 12 пар черепномозговых нервов). Эти волокна иннервируют гладкие мышцы глаза. Импульсы, поступающие по этим нервам, суживают зрачок. От группы клеток, заложенных в продолговатом мозгу, берут начало парасимпатические волокна, которые идут в составе лицевого и блуждающего нервов. В составе лицевого нерва (один из черепномозговых нервов) проходят волокна, которые образуют нерв, получивший название барабанной струны. Он является секреторным нервом подчелюстной слюнной железы, и его возбуждение вызывает слюноотделение23. Другая часть клетки продолговатого мозга дает начало блуждающему нерву (вагусу), который сильно разветвляется; образовавшиеся многочисленные веточки иннервируют сердце, пищевод, бронхи, желудок, кишечник, поджелудочную железу, надпочечники, почки, печень, селезенку. Из крестцового отдела спинного мозга берут начало парасимпатические волокна, которые идут в составе тазового нерва. Эти волокна иннервируют кишечник, половые органы и мочевой пузырь.
Эффекты, которые оказывает парасимпатическая нервная система на иннервируемые ею органы, представлены в таблице 4.2.
Симпатическая нервная система берет начало в грудинно-поясничных отделах спинного мозга. Значительная часть волокон, выйдя из спинного мозга, входит в узлы, находящиеся по обе стороны
позвоночника, и прерывается там, другие же преганглионарные волокна прерываются в ганглиях, которые находятся дальше от позвоночного столба. К таким узлам относятся шейные узлы, солнечное сплетение, брыжеечные узлы. Симпатическая нервная система иннервирует все ткани и органы. Она влияет на деятельность пищеварительных органов, гладкой сердечной мышцы, сосудов, потовых желез, почек, органов чувств, половых органов, желез внутренней секреции и т. д.
Академик Л. А. Орбели в первой половине прошлого столетия создал теорию об адаптационно-трофической функции симпатической нервной системы. Согласно этой теории, симпатическая нервная система играет в организме двойную роль. Она влияет на обмен веществ, протекающий в мышцах, что отражается на работе мышечной ткани. Эта функция и была названа трофической (от греч. трофос — питающий).
Адаптационное, т. е. приспосабливающее влияние симпатической нервной системы заключается в том, что под ее действием в органах и тканях происходит ряд преобразований, подготавливающих орган к работе в новых измененных условиях.
Существует определенная взаимосвязь между эмоциональным состоянием и деятельностью вегетативной нервной системы. Состояние страха, гнева, ярости и т. д. вызывает в организме комплекс изменений (так называемый «вегетативный криз»): повышение артериального давления, расширение или сужение сосудов, увеличение (уменьшение) силы и частоты сердечных сокращений, увеличение (уменьшение) глубины и частоты дыхания, интенсификация процесса потоотделения и т. д.
Все эти внешние вегетативные выражения тех или иных эмоций обусловлены влиянием коры больших полушарий и лимбической системы головного мозга на системы и органы, в первую очередь, через симпатическую нервную систему. Многочисленными экспериментами по выработке условных рефлексов было показано, что именно кора больших полушарий держит в своем ведении деятельность вегетативной нервной системы. Так, в одном из экспериментов бег, который вызывает изменения в деятельности сердца (увеличение силы и частоты сер-дечных сокращений), сочетался со звуком метронома. После нескольких опытов один звук метронома стал вызывать симпатическую активацию и, следовательно, указанные изменения в работе сердца.
Несмотря на то, что влияние парасимпатической и симпатической нервных систем часто противоположно (см. таблицу 4.2), их активность носит симбиотический характер. Только в результате согласованной деятельности обеих систем возможно достижение адаптации организма, адекватной той или иной ситуации.
Таблица 4.2