

Классификация нервно-паралитических овтв в соответствии с
механизмами токсического действия на организм
1. Действующие на холинореактивные синапсы |
1.1 Ингибиторы холинэстеразы: — ФОВ, карбаматы 1.2 Пресинаптические блокаторы высвобождения ацетилхолина: — ботулотоксин |
2. Действующие на ГАМК – реактивные синапсы |
2.1 Ингибиторы синтеза ГАМК: — производные гидразина 2.2 Антагонисты ГАМК (ГАМК-литики): — бициклофосфаты, норборнан 2.3 Пресинаптические блокаторы высвобождения ГАМК: — тетанотоксин |
3. Блокаторы Na+-ионных каналов возбудимых мембран |
— тетродотоксин, сакситоксин |
Ингибиторы холинэстеразы (непрямые холиномиметики), ГАМК-литики и вещества, блокирующие синтез ГАМК и ее высвобождение в синаптическую щель, инициируют при тяжелых интоксикациях судорожный синдром. Вещества, угнетающие высвобождение ацетилхолина и блокирующие Na+-ионные каналы возбудимых мембран, вызывают паралич произвольной мускулатуры.
4.1. Отравляющие вещества судорожного действия
4.1.1. Овтв, действующие на холинергические синапсы
Синаптические структуры, в которых медиаторную функцию выполняет ацетилхолин, обнаружены как в центральной нервной системе, так и на периферии.
Процесс передачи сигнала в холинергических синапсах может быть представлен несколькими этапами:
— синтез медиатора из холина и ацетата при участии энзима холинацетилтрансферазы (ХАТ) и его депонирование в пресинаптических везикулах;
— выделение ацетилхолина в синаптическую щель (спонтанно и при поступлении в нервное окончание возбуждающего сигнала);
— взаимодействие
ацетилхолина с холинорецепторами
постсинаптической (по некоторым данным,
и пресинаптической) мембраны и инициация
стимула в иннервируемой клетке;
— разрушение выделившегося в синаптическую щель нейромедиатора энзимом ацетилхолинэстеразой (АХЭ) и прекращение процесса передачи сигнала;
— захват пресинаптическими структурами высвободившегося холина.
Наиболее уязвимыми для действия высокотоксичных веществ являются этапы выделения ацетилхолина в синаптическую щель, взаимодействия медиатора с холинорецепторами и разрушения его энзимом АХЭ (схема 10).
Установлено, что нейромедиатор ацетилхолин и в ЦНС и в периферической нервной системе (ПНС) взаимодействует с двумя видами рецепторов — мускариновыми и никотиновыми, отличающимися способностью отвечать на воздействие ряда агонистов (М-холинорецепторы избирательно возбуждаются мускарином; Н-холинорецепторы — никотином), а также механизмом рецепции сигнала и его передачи на эффекторную клетку (нейрон, миоцит, железистую клетку).
Схема 10
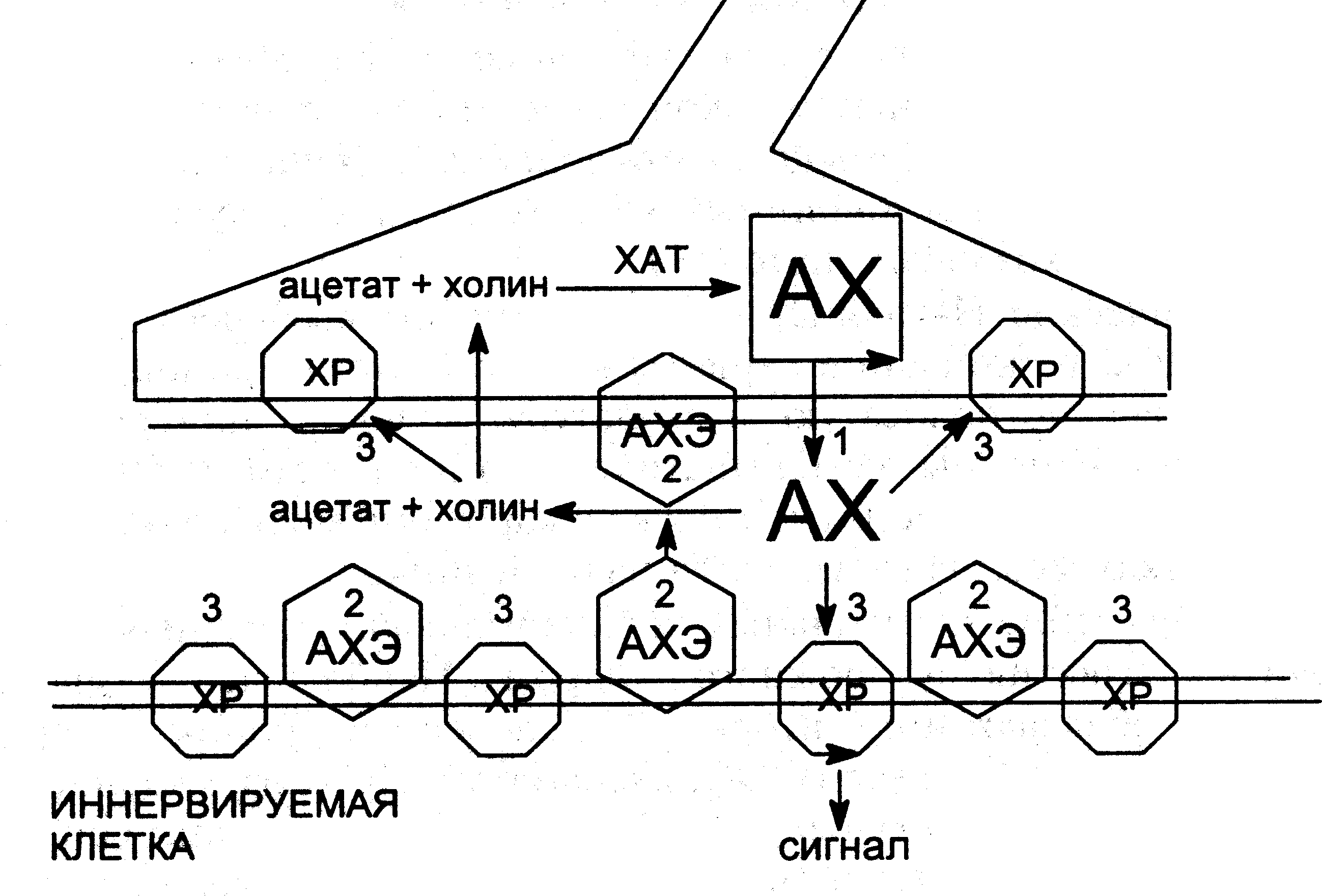
Схема функционирования холинергического синапса
АХ – ацетилхолин; ХР – холинорецептор; АХЭ - ацетилхолинэстераза;
1-3 – способы воздействия ядов на синапс: 1 – ботулотоксин; 2 – ингибиторы АХЭ;
3 – холинергические и антихолинергические средства
Н-холинорецепторы непосредственно связаны с ионными каналами постсинаптической мембраны для Na+, K+, Са2+. М-холинорецепторы также влияют на проницаемость ионных каналов для Na+ и К+. Однако это влияние опосредовано изменением активности сопряженных с рецептором ферментов, регулирующих содержание в иннервируемой клетке вторичных мессенджеров восприятия сигнала (цАМФ, цГМФ, фосфоинозитола и т.д.). Действие ацетилхолина на рецепторы приводит к усилению проницаемости постсинаптических мембран для ионов натрия, что сопровождается деполяризацией мембраны и ее возбуждением. Различается скорость генерации постсинаптического сигнала: в Н-холинергических синапсах изменение конформации ионных каналов развивается практически мгновенно, а в М-холинергических — постепенно.
Прерывание нервного импульса осуществляется путем гидролиза ацетилхолина ферментом ацетилхолинэстеразой.
В ЦНС плотность М-холинергических синапсов между нейронами существенно выше, чем Н-холинергических. Наивысшая — в хвостатом, прилежащем, чечевичном ядрах, гиппокампе, гипоталамусе, коре головного мозга, мозжечке млекопитающих и человека.
На периферии М-холинергические синапсы связывают окончания постганглионарных нервных волокон парасимпатической системы с иннервируемыми ими гладкомышечными волокнами (кишечника, бронхов, миокарда) и клетками экзокринных желез (слюнных, желудочно-кишечного тракта, бронхиальных, потовых).
Периферические Н-холинергические синапсы выявляются, главным образом, в области окончаний нервных волокон мотонейронов, иннервирующих поперечно-полосатую мускулатуру, волокон, иннервирующих ганглионарные нейроны парасимпатического и симпатического отделов вегетативной нервной системы, а также в каротидном клубочке, хромаффинных клетках мозгового вещества надпочечников.
Перевозбуждение центральных холинергических механизмов может приводить к развитию судорожного синдрома. Наибольшей активностью в этом плане обладают ингибиторы АХЭ, способные проникать через гематоэнцефалический барьер (ФОВ, карбаматы). Перевозбуждение периферических холинореактивных структур этими токсикантами сопровождается существенным, порой не совместимым с жизнью, нарушением функций дыхательной и сердечно-сосудистой систем.