

Биохимия_Ответы
.pdfфлавопротеин. На долю двух последних приходится до 80% митохондриальных флавопротеинов, выполняющих важную роль в биоэнергетике клетки. Негемовое железо связывается с белковым компонентом, отличающимся от гемсодержащих хромопротеинов. Железо ковалентно связано с атомом серы остатка цистеина в белке. При кислотном гидролизе такого белка освобождается железо и H2S. Несмотря на структурные отличия от цитохромов, негемовые флавопротеины выполняют аналогичную функцию в транспорте электронов благодаря способности переходить из окисленного в восстановленное состояние.
Нуклеопротеины состоят из белков и нуклеиновых кислот. Последние рассматриваются как простетические группы. В природе обнаружено 2 типа нуклеопротеинов, отличающихся друг от друга по составу, размерам и физико-химическим свойствам,– дезоксирибонуклеопротеины (ДНП) и рибонуклеопротеины (РНП). Названия нуклеопротеинов отражают только природу углеводного компонента (пентозы), входящего в состав нуклеиновых кислот. У РНП углевод представлен рибозой, у ДНП – дезоксирибозой.
Методы выделения и очистки белков
Для подробного исследования физико-химических и биологических свойств белков, а также для изучения их химического состава и структуры непременным условием является получение белков из природных источников в химически чистом, гомогенном состоянии. Последовательность операций по выделению белков обычно состоит в следующем: измельчение биологического материала (гомогенизация); извлечение белков, точнее, перевод белков в растворенное состояние (экстракция); выделение исследуемого белка из смеси других белков, т.е. очистка и получение индивидуального белка.
Белковые вещества весьма чувствительны к повышению температуры и действию многих химических реагентов (органические растворители, кислоты, щелочи). Поэтому обычные методы органической химии, применяемые для выделения того или иного вещества из смеси (нагревание, перегонка, возгонка, кристаллизация и др.), в данном случае неприемлемы.
Белки в этих условиях подвергаются денатурации, т.е. теряют некоторые существенные природные (нативные) свойства, в частности растворимость, биологическую активность. Разработаны эффективные методы выделения белков в мягких условиях, при низкой температуре (не выше 4°С), с применением щадящих нативную структуру химических реагентов.
8.Обмін вуглеводів. Розпад та біосинтез полісахаридів. Взаємоперетворення вуглеводів.
Метаболизм (обмен) углеводов в организме человека состоит в основном из следующих процессов:
1.Расщепление в пищеварительном тракте поступающих с пищей полисахаридов и дисахаридов до моносахаридов. Всасывание моносахаридов из кишечника в кровь.
2.Синтез и распад гликогена в тканях, прежде всего в печени.
3.Гликолиз. Понятие гликолиз означает расщепление глюкозы. Первоначально этим термином обозначали только анаэробное брожение, завершающееся образованием молочной
кислоты (лактата) или этанола и СО2. В настоящее время понятие гликолиз используется более широко для описания распада глюкозы, проходящего через образование глюкозо-6-фосфата, фруктозобисфосфата и пирувата как в отсутствие, так и в присутствии кислорода. В последнем случае употребляют термин аэробный гликолиз в отличие от анаэробного гликолиза , завершающегося образованием молочной кислоты (лактата).
4.Аэробный путь прямого окисления глюкозы или, как его называют,пентозофосфатный путь (пентозный цикл).
5.Взаимопревращение гексоз.
6.Аэробный метаболизм пирувата. Этот процесс выходит за рамки углеводного обмена, однако может рассматриваться как завершающая его стадия: окисление продукта гликолиза – пирувата.
7.Наконец, важным является процесс глюконеогенеза, или образование углеводов из неуглеводных продуктов. Такими продуктами являются в первую очередь пировиноградная и молочная кислоты, глицерин, аминокислоты и ряд других соединений.
Биосинтез полисахаридов протекает главным образом с участием нуклеозиддифосфатсахаров, служащих донорами моносахаридных (реже — дисахаридных) остатков, которые переносятся на
соответствующие олигосахаридные фрагменты строящегося полисахаридного биосинтеза гетерополисахаридов происходит путѐм последовательного включения М из соответствующих нуклеозиддифосфатсахаров в полисахаридную цепь. Известен и др. механизм, реализующийся при построенииПолисахариды бактериальных антигенов; вначале с участием липидных и нуклеотидных переносчиков сахаров синтезируются специфические, т. н. повторяющиеся звенья, из которых под
действием |
фермента |
полимеразы |
|
происходит |
синтез Полисахариды |
|
Разветвленные Полисахариды типа |
гликогена |
и |
амилопектина |
образуются |
путѐм |
|
внутримолекулярной ферментативной перестройки |
линейного Полисахариды Разрабатываются |
|||||
подходы к направленному химическому синтезу Полисахариды |
|
|
||||
В живых организмах Полисахариды, служащие основными резервами энергии, расщепляются внутри- и внеклеточными ферментами с образованием М и их производных, распадающихся далее с высвобождением энергии. Накопление и распад гликогена в печени человека и высших животных —
способ регулирования |
уровня глюкозы в |
крови. Мономерные продукты образуются или |
непосредственно путѐм |
последовательного |
отщепления от молекулы Полисахариды, или в |
результате ступенчатого распадаПолисахариды с промежуточным образованием олигосахаридов. Многие Полисахариды (крахмал, целлюлоза, пектиновые вещества и др.) применяют в пищевой, химической и др. отраслях промышленности, в медицине.
Взаимопревращение моносахаридов.
Происходит под действием ферментов эпимераз, преимущественно в печени. Все моносахариды способны превращаться друг в друга, но главное могут превращаться в глюкозу. Например, галактоза, фруктоза сначала активируются, а потом под действием ферментов превращаются в глюкозу.
9. Ліпіди, їхня біологічна роль. Загальні властивості, класифікація та номенклатура ліпідів.
Липиды играют важнейшую роль в процессах жизнедеятельности. Будучи одним из основных компонентов биологических мембран, липиды влияют на их проницаемость, участвуют в передаче нервного импульса, создании межклеточных контактов. Жир служит в организме весьма эффективным источником энергии либо при непосредственном использовании, либо потенциально – в форме запасов жировой ткани. В натуральных пищевых жирах содержатся жирорастворимые витамины и незаменимые жирные кислоты. Важная функция липидов – создание термоизоляционных покровов у животных и растений, защита органов и тканей от механических воздействий.
Классификация липидов
Существует несколько классификаций липидов. Наибольшее распространение получила классификация, основанная на структурных особенностях липидов. По этой классификации различают следующие основные классы липидов.
Простые липиды: сложные эфиры жирных кислот с различными спиртами.
1.Глицериды (ацилглицерины, или ацилглицеролы – по международной номенклатуре) представляют собой сложные эфиры трехатомного спирта глицерина и высших жирных кислот.
2.Воска: сложные эфиры высших жирных кислот и одноатомных или двухатомных спиртов. Сложные липиды: сложные эфиры жирных кислот со спиртами, дополнительно содержащие и другие группы.
1.Фосфолипиды: липиды, содержащие, помимо жирных кислот и спирта, остаток фосфорной кислоты. В их состав часто входят азотистые основания и другие компоненты:
а) глицерофосфолипиды (в роли спирта выступает глицерол); б) сфинголипиды (в роли спирта – сфингозин).
2.Гликолипиды (гликосфинголипиды).
3.Стероиды.
4.Другие сложные липиды: сульфолипиды, аминолипиды. К этому классу можно отнести и липопротеины.
Предшественники и производные липидов: жирные кислоты, глицерол, стеролы и прочие спирты (помимо глицерола и стеролов), альдегиды жирных кислот, углеводороды, жирорастворимые витамины и гормоны.
Взависимости от функций в организме различают две группы липидов:
1.Резервные липиды (жиры жировых депо), их количество и состав непостоянны, зависят от режима питания и физического состояния организма.
2.Структурные липиды — их количество и состав в организме строго постоянны, генетически обусловлены и в норме не зависят от режима питания, функционального состояния организма. Жирные кислоты служат своеобразными строительными блоками для большинства липидов. Они являются алифатическими карбоновыми кислотами. В настоящее время из живых организмов выделено свыше 70 жирных кислот. Всех их можно разделить на две большие группы: 1) насыщенные жирные кислоты; 2) ненасыщенные жирные кислоты (содержат в составе углеродного скелета ацила одну или несколько двойных связей). Cмесь жирных кислот, получаемая при гидролизе липидов из различныхприродных источников, обычно содержит как насыщенные, так и ненасыщенные жирные кислоты.
В зависимости от числа углеродных атомов в составе жирной кислоты различают короткоцепочечные жирные кислоты (радикал содержит от 2 до 4 углеродных атомов), среднецепочечные жирные кислоты (радикал содержит от 6 до 12 углеродных атомов), длинноцепочечные жирные кислоты (радикал содержит от 14 до 22 углеродных атомов) и жирные кислоты с очень длинной цепью (от 24 до 26 углеродных атомов). В типичных липидах животного происхождения преобладают жирные кислоты с длинной углеродной цепью, содержащей обычно четное число атомов углерода (от 14 до 24), для которых также употребим термин "высшие жирные кислоты". Среднецепочечные жирные кислоты, также как, и кислоты с очень длинной цепью, встречаются в организме лишь в небольших количествах. Что касается короткоцепочечных жирных кислот, то они вообще редко встречаются в составе липидов животных.
10. Амінокислоти, їхні властивості та класифікація. Методи визначення.
Аминокислоты - класс органических соединений, содержащих карбоксильные (-COOH) и
аминогруппы (-NH2); обладают свойствами и кислот, и оснований. Участвуют в обмене азотистых веществ всех организмов (исходное соединение при биосинтезе гормонов, витаминов, медиаторов,
пигментов, пуриновых и пиримидиновых оснований, алкалоидов и др.). Природных аминокислот св. 150. Около 20 важнейших аминикислот служат мономерными звеньями, из которых построены все белки (порядок включения аминокислот в них определяется генетическим кодом). Большинство микроорганизмов и растения синтезируют необходимые им аминокислоты; животные и человек не способны к образованию т. н. незаменимых аминокислот, получаемых с пищей.
Аминокислоты находят широкое применение в качестве пищевых добавок. Например,
лизином, триптофаном, треонином и метионином обогащают корма сельскохозяйственных животных, добавление натриевой соли глутаминовой кислоты (глутамата натрия) придает ряду продуктов мясной вкус. В смеси или отдельно аминокислоты применяют в медицине, в том числе при нарушениях обмена веществ и заболеваниях органов пищеварения, при некоторых заболеваниях центральной нервной системы (g-аминомасляная и глутаминовая кислоты, ДОФА). Аминокислоты используются при изготовлении лекарственных препаратов, красителей, в парфюмерной промышленности, в производстве моющих средств, синтетических волокон и пленки и т. д. Для хозяйственных и медицинских нужд аминокислоты получают с помощью микроорганизмов путем так называемого микробиологического синтеза (лизин, триптофан, треонин); их выделяют также из гидролизатов природных белков (пролин, цистеин, аргинин, гистидин). Но наиболее перспективны смешанные способы получения, совмещающие методы химического синтеза и использование ферментов.
Классификация аминокислот
Аминокислоты классифицируют на основе химического строения радикалов, хотя были предложены и другие принципы. Различают ароматические и алифатические аминокислоты, а также аминокислоты, содержащие серу или гидроксильные группы. Часто классификация основана на природе заряда аминокислоты. Если радикал нейтральный (такие аминокислоты содержат только одну амино- и одну карбоксильную группу), то они называются нейтральными аминокислотами.
Если же аминокислота содержит избыток аминоили карбоксильных групп, то она называется соответственно основной или кислой аминокислотой.
Современная рациональная классификация аминокислот основана на полярности радикалов,
т.е. способности их к взаимодействию с водой. Она включает четыре класса аминокислот:
1)неполярные (гидрофобные)
2)полярные (гидрофильные) незаряженные
3)отрицательно заряженные
4)положительно заряженные при физиологических значениях pH
11. Анаеробний розпад вуглеводів.
Анаэробный распад углеводов (анаэробный гликолиз) – это распад при котором разложение углеводов происходит без доступа кислорода, в начале образуется пировиноградная кислота, которая восстанавливается в молочную, предшествует аэробному гликолизу. В основном углеводы выполняют энергодативную функцию. Главными источниками энергии являются глюкоза и гликоген. Кроме того, из углеводов могут синтезироваться липиды, некоторые аминокислоты, пентозы. Углеводы входят как составная часть в структурно-функциональные компоненты клетки гликолипиды и гликопротеины.
Расщепление углеводов
Суточная норма углеводов в пище составляет 400-500 г. Основными углеводами пищи являются:
крахмалразветвленный гомополисахарид из глюкозы. Мономеры линейных участков соединены -1,4-гликозидными связями, а в местах разветвления -1,6 связями;
дисахариды - сахароза (глк-(-1,2)-фру), лактоза (гал-(бета-1,4)-глк), мальтоза (глк-(-1,4)-глк). При переваривании углеводов в желудочно-кишечном тракте происходит ферментативный
гидролиз гликозидных связей и образование моносахаридов, главным из которых является глюкоза. Гидролиз крахмала начинается в полости рта при участии амилазы слюны, которая частично расщепляет внутренние -1,4-гликозидные связи, образуя менее крупные, чем крахмал молекулы - декстрины. Далее гидролиз крахмала продолжается в верхнем отделе кишечника под действием панкреатической амилазы, также расщепляющей -1,4-гликозидные связи. В результате из крахмала образуются дисахаридные остатки мальтозы и изомальтозы (глк-(-1,6)-глк). Гидролиз всех дисахаридов происходит на поверхности клеток кишечника и катализируется специфическими ферментами: сахаразой, лактазой, мальтазой и изомальтазой. Эти гликозидазы синтезируются в клетках кишечника.
Всасывание моносахаридов из кишечника в кровь осуществляется путем облегченной диффузии. Если концентрация глюкозы в кишечнике невелика, то ее транспорт может происходить за счет градиента концентрации ионов натрия, создаваемого Na+, K+-AТФ-азой.
Глюкоза играет главную роль в метаболизме, так как именно она является основным источником энергии. Глюкоза может превращаться практически во все моносахариды, в то же время возможно и обратное превращение.
катаболизм глюкозы - гликолиз;
синтез глюкозы - глюконеогенез;
депонирование и распад гликогена;
синтез пентоз - пентозофосфатные пути.
Транспорт глюкозы в клетки С кровью воротной вены большая часть глюкозы (около половины) из кишечника поступает в печень, остальная глюкоза через общий кровоток транспортируется в другие ткани. Концентрация глюкозы в крови в норме поддерживается на постоянном уровне и составляет 3,33-5,55 мкмоль/л, что соответствует 80-100 мг в 100 мл крови. Транспорт глюкозы в клетки носит характер облегченной диффузии, но регулируется во многих клетках гормоном поджелудочной железы - инсулином, действие которого приводит к перемещению белков-переносчиков из цитозоля в плазматическую мембрану.
Затем с помощью этих белков глюкоза транспортируется в клетку по градиенту концентрации. Скорость поступления глюкозы в мозг и печень не зависит от инсулина и определяется только концентрацией ее в крови. Эти ткани называются инсулинонезависимыми.
Катаболизм глюкозы Гликолиз - это серия реакций, в результате которых глюкоза распадается на две молекулы
пирувата (аэробный гликолиз) или две молекулы лактата (анаэробный гликолиз). Все десять реакций гликолиза протекают в цитозоле и характерны для всех органов и тканей. Аэробный распад глюкозы включает реакции аэробного гликолиза и последующее окисление пирувата в реакциях катаболизма. Таким образом, аэробный распад глюкозыэто предельное ее окисление до СО2 и Н2О, а анаэробный гликолиз - это специфический путь катаболизма, т. е. часть аэробного распада глюкозы. Анаэробный распад включает те же реакции специфического пути распада глюкозы до пирувата, но с последующим превращением пирувата в лактат (т. е. термины анаэробный распад и анаэробный гликолиз совпадают).
Три основных типа гликолиза основных этапов.
На первом этапе превращениям подвергаются гексозы, на втором - триозы, на третьем - карбоновые кислоты.
Характеристика гликолиза:
большинство реакций обратимо, за исключением трех (реакций 1, 3, 10);
все метаболиты находятся в фосфорилированной форме;
источником фосфатной группы в реакциях фосфорилирования являются АТФ (реакции 1, 3) или неорганический фосфат (реакция 6);
регенерация NAD+, являющаяся необходимым условием протекания гликолиза, происходит при аэробном гликолизе посредством дыхательной цепи. В этом случае водород транспортируется в митохондрии с помощью челночного механизма при участии переносчиков. Это происходит потому, что мембрана митоходрий непроницаема для протонов. При анаэробном гликолизе регенерации NAD+ осуществляется независимо от дыхательной цепи. В этом случае акцептором водорода от NADH является пируват, который восстанавливается в лактат;
образование АТФ при гликолизе может идти двумя путями: либо субстратным фосфорилированием, когда для фосфорилирования AДФ используется энергия макроэргической связи субстрата (реакции 7, 9), либо путем окислительного
фосфорилирования AДФ, сопряженного с дыхательной цепью (реакция 6). Значение анаэробного гликолиза
Анаэробный и аэробный гликолиз энергетически неравноценны. Образование двух моль лактата из глюкозы сопровождается синтезом всего двух моль АТФ, потому что NADH, полученный при окислении глицероальдегидфосфата, не используется дыхательной цепью, а акцептируется пируватом.
Анаэробный распад глюкозы.
Анаэробный гликолиз, несмотря на небольшой энергетический эффект, является основным источником энергии для скелетных мышц в начальном периоде интенсивной работы, т. е. в условиях, когда снабжение кислородом ограничено. Кроме того, зрелые эритроциты извлекают энергию за счет анаэробного окисления глюкозы, потому что не имеют митохондрий.
Депонирование и распад гликогена Гликоген - основная форма депонирования глюкозы в клетках животных. У растений эту же
функцию выполняет крахмал. В структурном отношении гликоген, как и крахмал, представляет собой разветвленный полимер из глюкозы.
Однако гликоген более разветвлен и компактен. Ветвление обеспечивает быстрое освобождение при распаде гликогена большого количества концевых мономеров. Синтез и распад гликогена не являются обращением друг в друга, эти процессы происходят разными путями. Биосинтез/гликогена.
Гликоген синтезируется в период пищеварения (в течение 1-2 ч после приема углеводной пищи). Гликогенез особенно интенсивно протекает в печени и скелетных мышцах. В начальных реакциях образуется UDF-глюкоза (реакция 3), которая является активированной формой глюкозы, непосредственно включающейся в реакцию полимеризации (реакция 4). Эта последняя реакция катализируется гликогенсинтазой, которая присоединяет глюкозу к олигосахариду или к уже имеющейся в клетке молекуле гликогена, наращивая цепь новыми мономерами. Для подготовки и включения в растущую полисахаридную цепь требуется энергия 1 моль АТР и 1 моль UTP. Ветвление полисахаридной цепи происходит при участии фермента амило - -1,4--1,6-гликозил- трансферазы путем разрыва одной -1,4-связи и переноса олигосахаридного остатка от конца растущей цепи к ее середине с образованием в этом месте -1,6-гликозидной связи. Молекула гликогена содержит до 1 млн остатков глюкозы, следовательно, на синтез расходуется значительное количество энергии. Необходимость превращения глюкозы в гликоген связана с тем, что накопление значительного количества глюкозы в клетке привело бы к повышению осмотического давления, так как глюкоза хорошо растворимое вещество. Напротив, гликоген содержится в клетке в виде гранул, и мало растворим. Распад гликогена - гликогенолиз - происходит в период между приемами пищи.
12. Характеристика основних ліпідів організму людини: класифікація, добова потреба та біологічна роль. Будова простих та складних ліпідів.
Липиды- нерастворимые в воде низкомолекулярные органические маслянистые или жирные вещества, являющиеся сложными эфирами жирных кислот и спиртов, которые могут быть экстронированы из клеток животных, растений и микроорганизмов неполярными растворителями, такими как хлороформ, бензол,эфир.
Биологическая роль липидов
Липиды играют важнейшую роль в процессах жизнедеятельности. Будучи одним из основных компонентов биологических мембран, липиды влияют на их проницаемость, участвуют в передаче нервного импульса, создании межклеточных контактов. Жир служит в организме весьма эффективным источником энергии либо при непосредственном использовании, либо потенциально – в форме запасов жировой ткани. В натуральных пищевых жирах содержатся жирорастворимые витамины и «незаменимые» жирные кислоты. Важная функция липидов – создание термоизоляционных покровов у животных и растений, защита органов и тканей от механических воздействий.
Функции липидов в организме:
1 – пластическая функция: они участвуют в построении мембран клеток всех органов и тканей и образовании многих биологически важных соединений (гормоны, жирорастворимые витамины).
2– энергетическая функция: липиды обеспечивают 25-30 % энергетических потребностей организма. Распад 1 г жира – 9,3 ккал.
3– жиры являются запасными питательными веществами, их депо – подкожная клетчатка, околопочечная капсула.
4– защитная функция липидов: они участвуют в терморегуляции, защищают кожу от высыхания, органы – от сотрясений.
5- выполняют функцию защитных оболочек, предохраняющих от инфекции или излишней потери или накопления воды.
6- обеспечивают всасывание жирорастворимых витаминов.
Классификация липидов
Различают следующие основные классы липидов:
Простые липиды: сложные эфиры жирных кислот с различными спиртами.
1.Глицериды (ацилглицерины, или ацилглицеролы – по международной номенклатуре) представляют собой сложные эфиры трехатомного спирта глицерина и высших жирных кислот.
2.Воска: сложные эфиры высших жирных кислот и одноатомных или двухатомных спиртов. Сложные липиды: сложные эфиры жирных кислот со спиртами, дополнительно содержащие и другие группы.
1.Фосфолипиды: липиды, содержащие, помимо жирных кислот и спирта, остаток фосфорной кислоты. В их состав часто входят азотистые основания и другие компоненты:
а) глицерофосфолипиды (в роли спирта выступает глицерол); б) сфинголипиды (в роли спирта – сфингозин).
2.Гликолипиды (гликосфинголипиды).
3.Стероиды.
4.Другие сложные липиды: сульфолипиды, аминолипиды. К этому классу можно отнести и липопротеины. Предшественники и производные липидов: жирные кислоты, глицерол, стеролы и прочие спирты (помимо глицерола и стеролов), альдегиды жирных кислот, углеводороды, жирорастворимые витамины и гормоны.
Строение простых и сложных липидов Простые липиды. Молекула простых липидов не содержит атомов азота, фосфора, серы. К ним
относят производные одноатомных (высших с 14—22 атомами углерода) карбоновых кислот и одно- и многоатомных спиртов (в первую очередь трех атомного спирта — глицерина).
Сложные липиды. Молекула их построена из остатков спиртов, высокомолекулярных жирных кисло фосфорной кислоты, азотистых оснований (чаще всего холин и этаноламина, аминокислот и некоторых других соединений. Фосфолипиды - обязательный компонент клеток.
Вместе с белками и углеводами фосфолипиды участвуют в построении мембран (перегородок) клеток и субклеточных структур (органелл), выполняя роль несущих конструкций мембран.
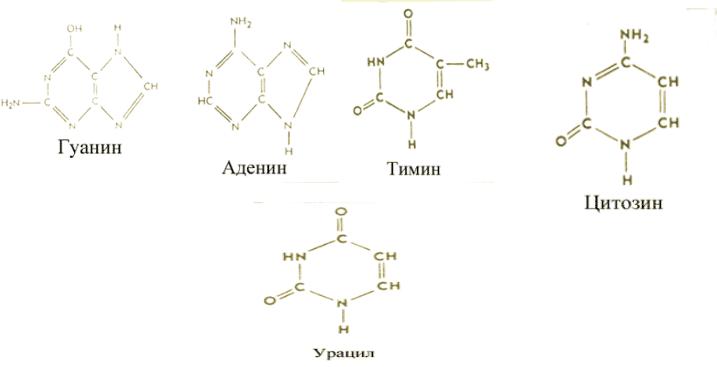
13. Нуклеїнові кислоти. Будова нуклеїнових кислот. Пуринові та піримідинові основи, мононуклеотиди, нуклеозід ди- і трифосфати, їхня фізіологічна роль.
Нуклеиновые кислоты - природные высокомолекулярные органические соединения, обеспечивающие хранение и передачу наследственной(генетической) информации в живых организмах.
В природе существуют нуклеиновые кислоты двух типов, различающиеся по составу, строению и функциям. Одна из них содержит углеводный компонент дезоксирибозу и названа дезоксирибонуклеиновой кислотой (ДНК). Другая содержит рибозу и названа рибонуклеиновой кислотой (РНК).
ДНК — представляет собой двухцепочечный биологический полимер, мономерами которого являются нуклеотиды, содержащие одно из азотистых оснований, дезоксирибозу и остаток фосфорной кислоты. Полинуклеотидные цепи молекулы ДНК антипараллельны и соединены друг с другом водородными связями по принципу комплиментарности. ДНК состоит из нуклеотидов: пуриновых оснований аденина(А) и гуанина (Г) и пиримидиновых оснований цитозина(Ц) и тимина Т).
РНК состоит из тех же оснований с различием лишь в то, что у РНК вместо тимина присутствует урацил(У). Тимин отличается от урацила наличием метильной группы (-СН3), которой нет в урациле.
Э. Чаргафф обнаружил, что количество пуринового основания аденина (А) равно количеству пиримидинового основания тимина (Т), т. е. А = Т. Сходным образом количество второго пурина — гуанина (Г) всегда равно количеству второго пиримидина—цитозина (Ц),т. е. Г = Ц. Таким образом, число пуриновых оснований в ДНК всегда равно числу пиримидиновых, количество аденина равно количеству тимина, а гуанина — количеству цитозина. Такая закономерность получила название правил Чаргаффа.
Картина полностью прояснилась в 1953 г., когда американский биохимик Дж. Уотсон и английский физик Ф. К р и к, исследуя структуру молекулы ДНК, пришли к выводу, что сахарофосфатный остов находится на периферии молекулы ДНК, а пуриновые и пиримидиновые основания — в середине. Причем последние ориентированы таким образом, что между основаниями из противоположных цепей могут образоваться водородные связи. Из построенной ими модели выявилось, что какой-либо пурин в одной цепи всегда связан водородными связями с одним из пиримидинов в другой цепи. Такие пары имеют одинаковый размер по всей длине молекулы. Не менее важно то, что аденин может спариваться лишь с тимином, а гуанин только с цитозином. При этом между аденином и тимином образуются две водородные связи, а между гуанином и цитозином три. Противоположные последовательности и соответствующие полинуклеотидные партнеры называются комплементарными. Хотя водородные связи, стабилизирующие пары оснований, относительно
слабы, каждая молекула ДНК содержит так много пар, что в физиологических условиях (температура, рН) комплементарные цепи никогда самостоятельно не разделяются.
Функция у ДНК одна - хранение генетической информации.
РНК - также полимер, мономерами которой являются нуклеотиды. РНК представляет собой однонитевую молекулу. Она построена таким же образом, как и одна из цепей ДНК. Нуклеотиды РНК очень близки, хотя и не тождественны, нуклеотидам ДНК. Их тоже четыре и они состоят из азостистого основания, пентозы и фосфорной кислоты. Три азотистых основания совершенно такие же, как в ДНК: — Аденин, Гуанин и Цитозин. Однако вместо Тимина у ДНК, в РНК присутствует близкий к нему по строению пиримидин - урацил.Различие между ДНК и РНК существует также в характере углевода: в нуклеотидах ДНК углевод — дезоксирибоза, у РНК — рибоза. В отличие от ДНК, содержание которой в клетках конкретных организмов относительно постоянно, содержание РНК сильно в них колеблется. Оно заметно повышено в клетках, в которых происходит синтез белка.
Функции РНК По выполнению функций выделяют несколько видов РНК.
Транспортная РНК(т-РНК). Молекулы т-РНК самые короткие: они состоят всего из 80—100 нуклеотидов. Молекулярная масса таких частиц равна 25—30 тыс. Транспортная РНК в основном содержится в цитоплазме клетки. Функция состоит в переносе аминокислот в рибосомы, к месту синтеза белка. Из общего содержания РНК клетки на долю т-РНК приходится около 10%.
Рибосомная РНК (р-РНК). Это самые крупные РНК в их молекулы входит 3—5 тыс. нуклеотидов, соответственно их молекулярная масса достигает 1,0—1, 5 млн. Рибосомная РНК составляет существенную часть структуры рибосомы. Из общего содержания РНК в клетке на долю р-РНК приходится около 90%.
Информационная РНК (и-РНК), или матричная (м-РНК). Содержится в ядре и цитоплазме. Функция ее состоит в переносе информации о структуре белка от ДНК к месту синтеза белка в рибосомах. На долю и-РНК приходится примерно 0,5—1% от общего содержания РНК клетки.
Все виды РНК синтезируются на ДНК, которая служит своего рода матрицей.
АТФ - аденозинтрифосфорная кислота. Аденозинмонофосфорная кислота (АМФ) входит в состав всех РНК; при присоединении еще двух молекул фосфорной кислоты (НзРО4) она превращается в АТФ и становится источником энергии, которая запасается в двух последних остатках фосфатов: Как во всякий нуклеотид, в АТФ входит остаток азотистого основания (аденин), пентоза (рибоза) и остатки фосфорной кислоты (у АТФ их три). Из состава АТФ под действием фермента АТФ-азы отщепляются остатки фосфорной кислоты.При отщеплении одной молекулы фосфорной кислоты АТФ переходит в АДФ (аденозиндифосфорная кислота), а если отщепляются две молекулы фосфорной кислоты, АТФ переходит в АМФ (аденозинмонофосфорная кислота). Реакции отщепления каждой молекулы фосфорной кислоты сопровождаются освобождением 419 кДж/моль. Для того чтобы подчеркнуть высокую энергетическую «стоимость» фосфорнокислородной связи в АТФ, ее принято обозначать знаком ~ и называть макроэргической связью. В АТФ имеются две макроэргические связи.
Значение АТФ в жизни клетки велико, она играет центральную роль в клеточных превращениях энергии. В реакциях с участием АТФ она, как правило, теряет одну молекулу фосфорной кислоты и переходит в АДФ. А далее АДФ может присоединить остаток фосфорной кислоты с поглощением 419 кДж/моль, восстановив запас энергии. Основной синтез АТФ происходит в митохондриях.
14. Аеробний розпад вуглеводів.
Аэробный распад углеводов – разложение углеводов при участии кислорода, конечными продуктами являются углекислота и вода при полном освобождении энергии АТФ.
Энергетическое значение аэробного распада глюкозы.
Ваэробном гликолизе образуется 10 моль АТФР на 1 моль глюкозы. Так, в реакциях 7, 10 образуется 4 моль АТФ путем субстратного фосфорилирования, а в реакции 6 синтезируется 6 моль АТФ (на 2 моль глицероальдегидфосфата) путем окислительного фосфорилирования.
Баланс аэробного гликолиза.
Суммарный эффект аэробного гликолиза составляет 8 моль АТФ, так как в реакциях 1 и 3 используется 2 моль АТФ. Дальнейшее окисление двух моль пируват в общих путях катаболизма сопровождается синтезом 30 моль АТФ (по 15 моль на каждую молекулу пирувата. Следовательно, суммарный энергетический эффект аэробного распада глюкозы до конечных продуктов составляет 38 моль АТФ.
Восновном углеводы выполняют энергодативную функцию. Главными источниками энергии являются глюкоза и гликоген. Кроме того, из углеводов могут синтезироваться липиды, некоторые аминокислоты, пентозы. Углеводы входят как составная часть в структурно-функциональные компоненты клетки - гликолипиды и гликопротеины.
Расщепление углеводов. Суточная норма углеводов в пище составляет 400-500 г. Основными углеводами пищи являются: крахмалразветвленный гомополисахарид из глюкозы. Мономеры линейных участков соединены -1,4-гликозидными связями, а в местах разветвления -1,6 связями;
дисахариды - сахароза (глк-(-1,2)-фру), лактоза (гал-(бета-1,4)-глк), мальтоза (глк-(-1,4)-глк). При переваривании углеводов в желудочно-кишечном тракте происходит ферментативный
гидролиз гликозидных связей и образование моносахаридов, главным из которых является глюкоза. Гидролиз крахмала начинается в полости рта при участии амилазы слюны, которая частично расщепляет внутренние -1,4-гликозидные связи, образуя менее крупные, чем крахмал молекулы - декстрины. Далее гидролиз крахмала продолжается в верхнем отделе кишечника под действием панкреатической амилазы, также расщепляющей -1,4-гликозидные связи. В результате из крахмала образуются дисахаридные остатки мальтозы и изомальтозы (глк-(-1,6)-глк). Гидролиз всех дисахаридов происходит на поверхности клеток кишечника и катализируется специфическими ферментами: сахаразой, лактазой, мальтазой и изомальтазой. Эти гликозидазы синтезируются в клетках кишечника.
Всасывание моносахаридов из кишечника в кровь осуществляется путем облегченной диффузии. Если концентрация глюкозы в кишечнике невелика, то ее транспорт может происходить за счет градиента концентрации ионов натрия, создаваемого Na+, K+-AТФ-азой.
Глюкоза играет главную роль в метаболизме, так как именно она является основным источником энергии. Глюкоза может превращаться практически во все моносахариды, в то же время возможно и обратное превращение.
Полное рассмотрение метаболизма глюкозы не входит в нашу задачу, поэтому сосредоточимся на основных путях:катаболизм глюкозы - гликолиз; интез глюкозы - глюконеогенез; депонирование и распад гликогена; синтез пентоз - пентозофосфатные пути.
Транспорт глюкозы в клетки
С кровью воротной вены большая часть глюкозы (около половины) из кишечника поступает в печень, остальная глюкоза через общий кровоток транспортируется в другие ткани. Концентрация глюкозы в крови в норме поддерживается на постоянном уровне и составляет 3,33-5,55 мкмоль/л, что соответствует 80-100 мг в 100 мл крови. Транспорт глюкозы в клетки носит характер облегченной диффузии, но регулируется во многих клетках гормоном поджелудочной железы - инсулином, действие которого приводит к перемещению белков-переносчиков из цитозоля в плазматическую мембрану.