

Лекция №17. Ингибирование продуктом
Рассмотрим закономерности культивирования микроорганизмов в открытой проточной системе с учетом эффектов ингибирования образующихся продуктов для двух основных механизмов ингибирования: конкурентного и неконкурентного.
Система уравнений, описывающая кинетику процесса в режиме хемостата в том случае, если микроорганизм «чувствует» присутствие продукта, имеет вид
![]() (17.1)
(17.1)
![]() (17.2)
(17.2)
![]() (17.3)
(17.3)
Удельная скорость роста как функция концентрации субстрата и продукта реакции будет иметь вид
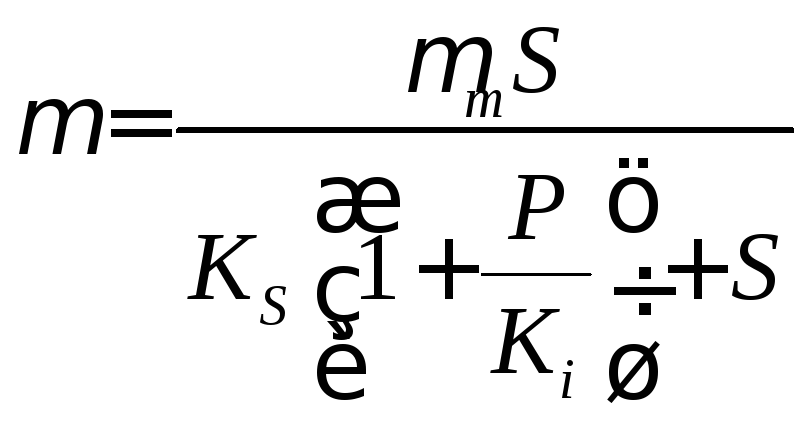 (конкурентное
ингибирование); (17.4)
(конкурентное
ингибирование); (17.4)
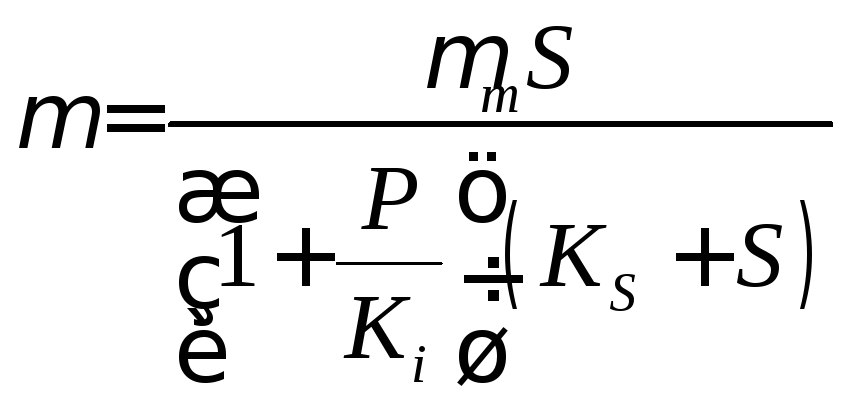 (неконкурентное
ингибирование). (17.5)
(неконкурентное
ингибирование). (17.5)
17.1. Конкурентное ингибирование продуктом
В
стационарном состоянии в условиях
![]() ,
,![]() ,
,![]() система
уравнений (17.1)—(17.3) с учетом (17.4)
может
быть представлена в виде
система
уравнений (17.1)—(17.3) с учетом (17.4)
может
быть представлена в виде
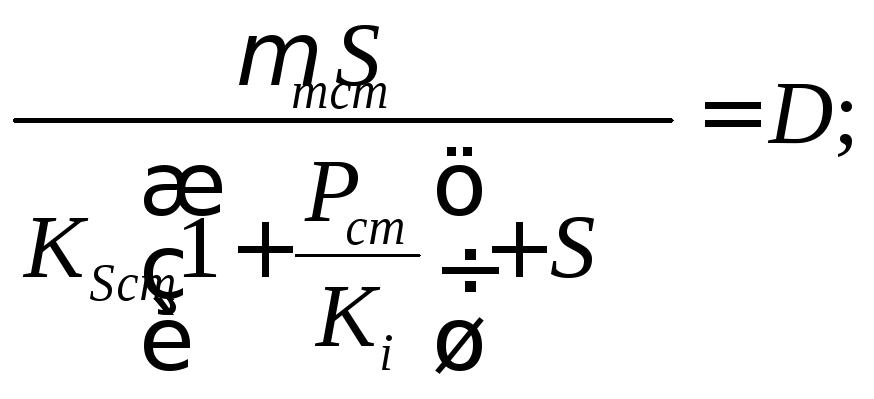 (17.6)
(17.6)
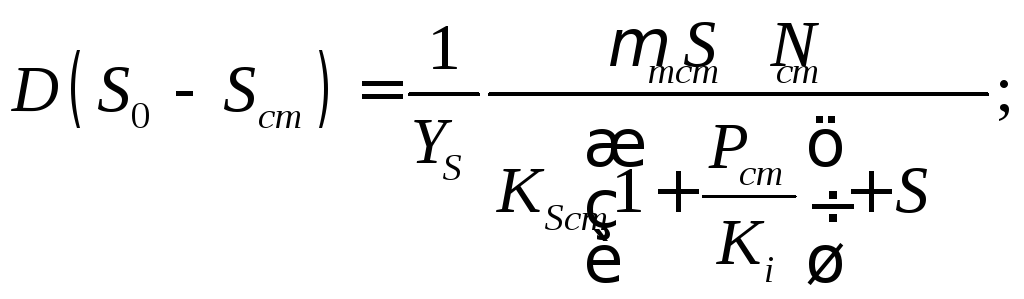 (17.7)
(17.7)
 (17.8)
(17.8)
Как и во всех рассмотренных ранее случаях, стационарная удельная скорость роста микроорганизма задается системе скоростью разбавления μ=D. Из стехиометрических соотношений следует
![]() (17.9)
(17.9)
Подстановка в (6) значения Рст, выраженного по уравнению (17.9), приводит к выражению
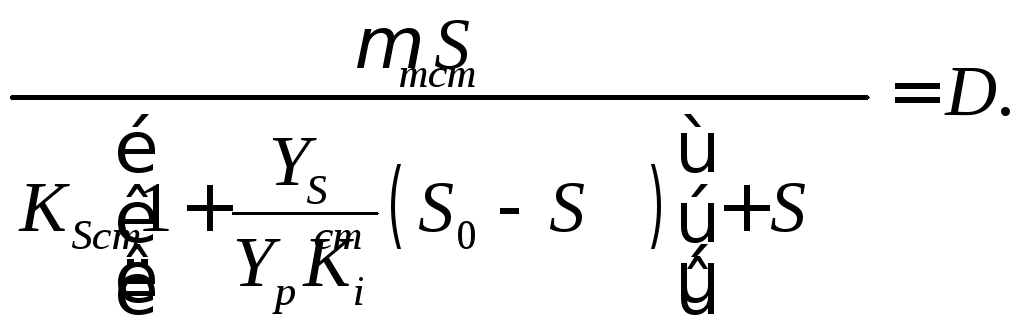 (17.10)
(17.10)
Это уравнение может быть решено относительно Sст:
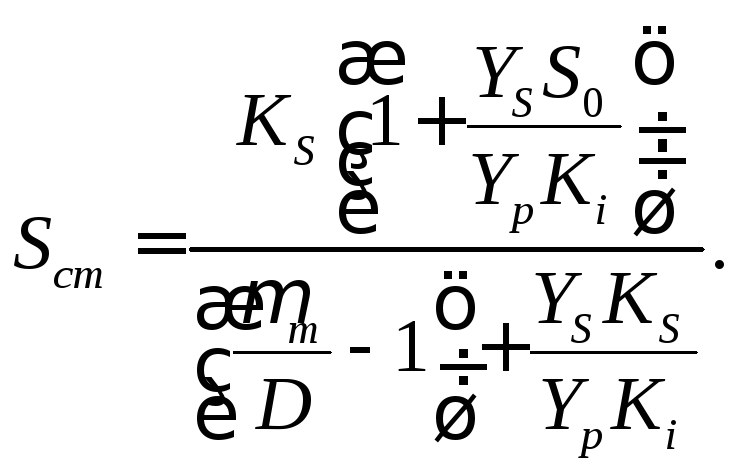 (17.11)
(17.11)
Уравнение (17.11) описывает зависимость стационарной концентрации субстрата от концентрации вводимого субстрата, скорости разбавления, кинетических параметров роста культуры μm и КS, константы равновесия ингибирования продуктом.
Следует подчеркнуть важную особенность ферментационного процесса с ингибированием продуктом — стационарный уровень концентрации субстрата является функцией начальной его концентрации. Это отличает процесс с ингибированием продуктом от процесса с неосложненным ростом. По физическому смыслу стационарная концентрация субстрата — величина, лежащая в диапазоне от нуля до S0; S0≥Sст≥0. Сопоставление этого неравенства с уравнением (17.12) позволяет выявить некоторые ограничивающие условия, в рамках которых возможно образование стационарного состояния для системы с конкурентным ингибированием продуктом.
Поскольку стационарная концентрация субстрата в ферментере не может превышать значения S0, можно записать
 (17.12)
(17.12)
Из этого неравенства следует
![]() (17.13)
(17.13)
Таким образом, существует критическая скорость разбавления, выше которой в системе невозможно стационарное состояние:
![]() (17.14)
(17.14)
Следовательно, критическая скорость разбавления для системы с конкурентным ингибированием продуктом та же, что и для неосложненного процесса.
Необходимо отметить одну особенность уравнения (17.11). При D=Dc стационарная концентрация субстрата равна S0; Sст(D=D∞) = S0 вне зависимости от остальных параметров, входящих в это уравнение.
Стационарные концентрации биомассы и продукта ферментационного превращения можно вычислить на основе Sст по уравнениям (17.7) и (17.8):
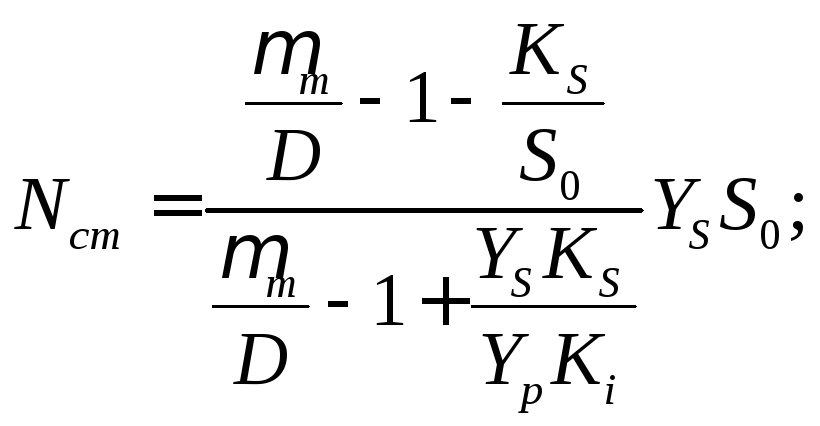 (17.15)
(17.15)
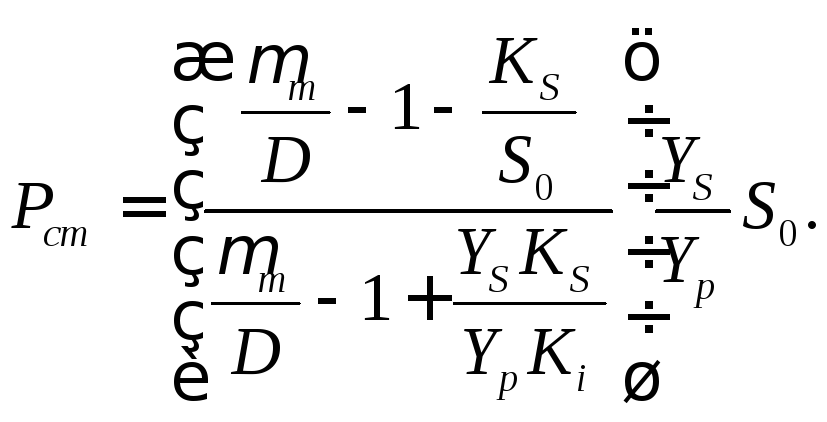 (17.16)
(17.16)
Полезно рассмотреть ряд частных случаев.
Константа ингибирования продуктом относительно велика (слабое ингибирование продуктом). Это соответствует условию
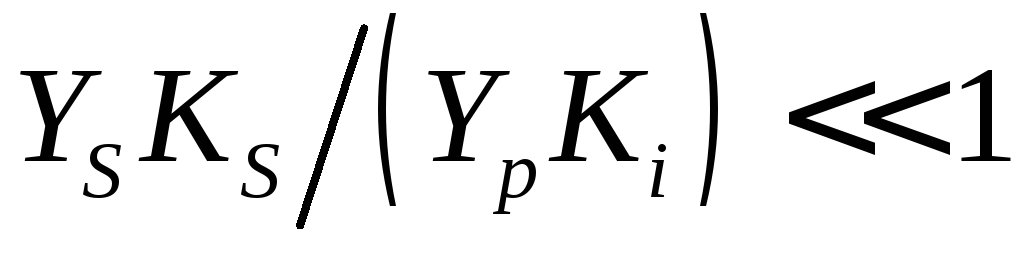 или
или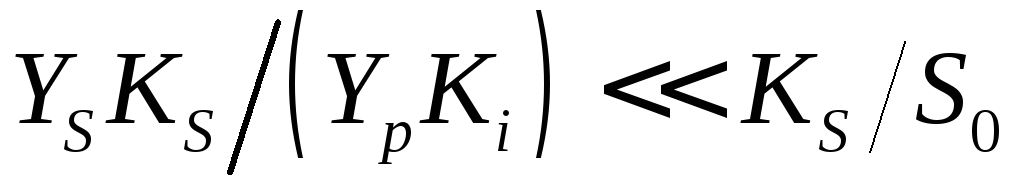 .
В
этом
режиме
имеем
.
В
этом
режиме
имеем
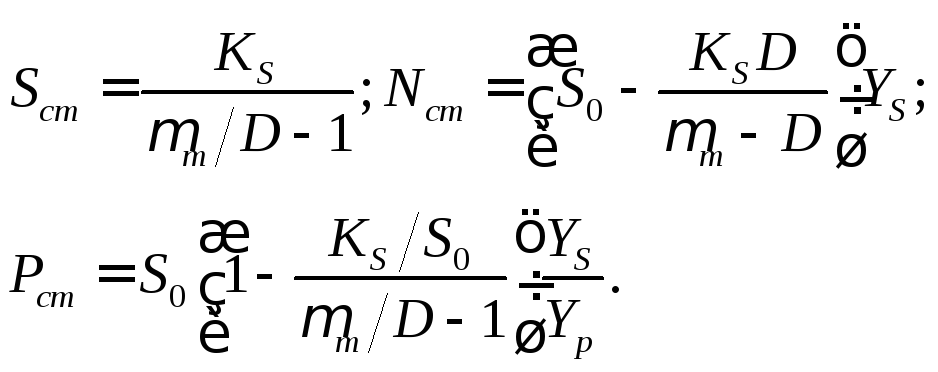 (17.17)
(17.17)
Уравнения (17.17) характеризуют протекание процесса, не осложненного ингибированием продуктом; в кинетике процесса ингибирование практически не проявляется.
2.
Константа
ингибирования продуктом очень мала
(сильное
эффективное
ингибирование продуктом), режим
![]() ,
,![]() .
При
этих условиях стационарные концентрации
компонентов будут даны уравнениями
.
При
этих условиях стационарные концентрации
компонентов будут даны уравнениями

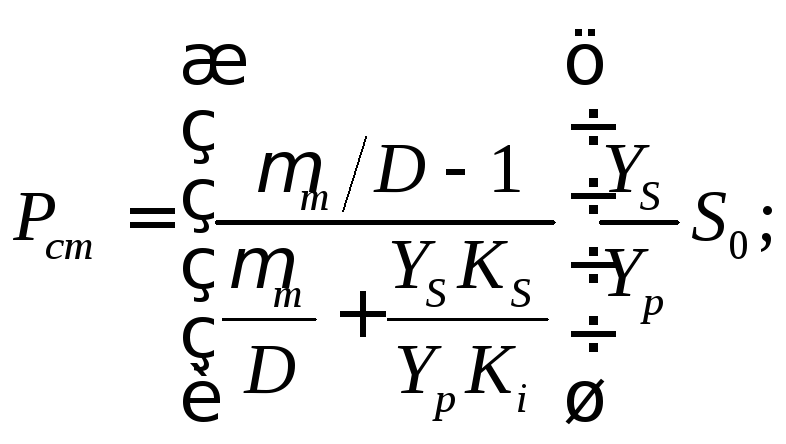 (17.18)
(17.18)
Если
режим работы хемостата таков, что
![]() т.
е. культивирование ведется при достаточно
высоких скоростях
разбавления, то из уравнений (17.18)
следует:
т.
е. культивирование ведется при достаточно
высоких скоростях
разбавления, то из уравнений (17.18)
следует:
К![]() ультура
«вымывается» из ферментера.
ультура
«вымывается» из ферментера.
В
этом режиме, для того чтобы наблюдать
процесс роста, необходимо
работать при весьма малых скоростях
разбавления, так
чтобы член
![]() вносил заметный вклад в уравнения роста.
вносил заметный вклад в уравнения роста.
3. Культивирование при критической скорости разбавления:
![]()
Из уравнений (17.18) следует:
![]()
Наблюдать рост культуры в этом режиме практически невозможно.
Для того чтобы идентифицировать механизм и определить параметры культуры, необходимо провести изучение стационарных уровней субстрата, продукта или биомассы в зависимости от начальных концентраций субстрата и вариации скоростей разбавления.
Стационарная концентрация субстрата. Уравнение (17.11) можно преобразовать к виду
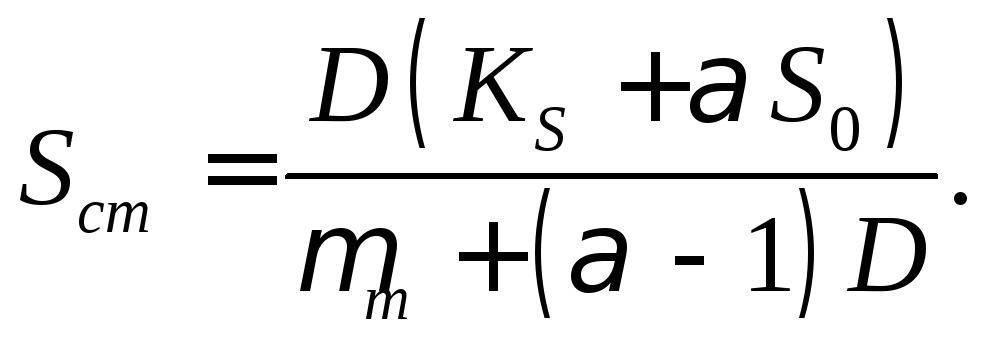 (17.19)
(17.19)
где
![]() - безразмерный параметр, характеризующий
относительную
эффективность ингибирования продуктом.
- безразмерный параметр, характеризующий
относительную
эффективность ингибирования продуктом.
Из уравнения (17.19) видно, что Sст линейно зависит от S0 и гиперболически от D. Его можно записать в форме
![]() (17.20)
(17.20)
Если процесс исследуется при данной постоянной скорости разбавления, но при различных значениях S0, то стационарная концентрация субстрата как функция S0 характеризуется линейной зависимостью, при этом параметры прямой даны следующими функциями:
![]() (17.21)
(17.21)
![]() (17.22)
(17.22)
Если α = 0 (ингибирование продуктом отсутствует), то стационарная концентрация субстрата не зависит от S0(tgφ = 0). Это соответствует кинетике неосложненного роста. Для определения параметров μm, КS и α необходимо знать значения а и tgφ хотя бы при двух различных скоростях разбавления. Если проведено полное исследование и известны а и tgφ при различных D, то можно воспользоваться уравнениями
![]() (17.23)
(17.23)
![]() .
(17.24)
.
(17.24)
Параметры этих прямых позволяют определить значения α, μm и КS. Исследование стационарного уровня субстрата в хемостате можно провести при различных скоростях разбавления в режиме двух или нескольких концентраций вводимого субстрата. Для этого уравнение (20) можно преобразовать к виду
![]() (17.25)
(17.25)
Экспериментальные данные при конкурентном ингибировании продуктом должны линеаризоваться в координатах этого уравнения при этом параметры равны
![]() (17.26)
(17.26)
![]() (17.27)
(17.27)
Прямая 1/Sст от 1/D пересекает ось абсцисс в точке
![]() (17.28)
(17.28)
При этом справедливы следующие условия (рис. 17.1)
α = 0 (1/D)n = 1/μm (кинетика неосложненного роста);
α = 1 (1/D)n = 0 (слабое ингибирование продуктом);
α >1 (1/D)n <0 (сильное ингибирование продуктом).

Рис.
17.1 Зависимость стационарной концентрации
субстрата от скорости разбавления в
обратных координатах для системы с
конкурентным ингибированием продуктом
при различных значениях
![]()
Точки пересечений прямой 1/Sст от 1/D с осью абсцисс не зависят от концентрации субстрата S0, что также следует из уравнения (17.28). Таким образом, 1/Sст от 1/D, полученные при различных значениях S0, пересекаются в одной точке. Из уравнений (17.26)—(17.27) следует, что тангенсы углов наклона прямых и отрезки, отсекаемые прямыми на оси ординат, связаны с концентрацией вводимого субстрата уравнениями
![]() (17.29)
(17.29)
![]() .
(17.30)
.
(17.30)
Эти анаморфозы позволяют определить искомые параметры КS, μm и α.
Стационарный уровень биомассы. Уравнение (17.15) можно записать в виде
![]() (17.31)
(17.31)
Таким образом, зависимость стационарного уровня биомассы от концентрации раствора вводимого субстрата при данной скорости разбавления должна представлять собой прямую линию со следующими параметрами:
![]() (17.32)
(17.32)
![]() (17.33)
(17.33)
Прямая Nст от S0 пересекает ось абсцисс в точке
![]() (17.34)
(17.34)
которая не зависит от α и экономического коэффициента YS, а определяется лишь скоростью разбавления и параметрами роста культуры. Если зависимость Nст от S0 исследована по крайней мере при двух различных скоростях разбавления, то по уравнению (17.34) можно найти KS и μm. В боле общем виде при серии определений Sn при различных скоростях разбавления целесообразно воспользоваться уравнением
![]() (17.35)
(17.35)
Для определения YS и α при найденных независимо параметрах KS и μm можно воспользоваться уравнениями (17.32) и (17.83). Например, уравнение (17.32) преобразуется в форму
![]() (17.36)
(17.36)
Из отрезка, отсекаемого на оси ординат, и тангенса угла наклона определяются параметры YS и значение α.