

Fiziologia_rasteny
.pdfжгутиков и ресничек, в движении которых, по-видимому, играют важную роль.
Микрофиламенты, подобно микротрубочкам, найдены практически во всех эукариотических клетках. Они представляют собой длинные нити толщиной 5—7 нм, состоящие из сократительного белка — актина. Пучки микрофиламентов встречаются во многих клетках высших растений и играют определенную роль в движении цитоплазмы.
Вакуоли отделены от цитоплазмы тонопластом. Жидкость, заполняющая вакуоль, представляет собой концентрированный водный раствор, содержащий минеральные соли, сахара, органиче- ские кислоты, кислород, диоксид углерода, пигменты и некоторые отходы жизнедеятельности или вторичные продукты метаболизма.
Согласно принятой в настоящее время точке зрения, вакуоли могут возникать только из предсуществовавших уже вакуолей или из каких-то других полостей клетки, ограниченных мембранами, например при разрастании пузыревидных выпячиваний эндоплазматической сети или увеличении пузырьков Гольджи.
Вакуоли выполняют следующие функции:
1.Вода обычно поступает в концентрированный вакуолярный сок путем осмоса через избирательно проницаемый тонопласт.
Âрезультате в клетке развивается тургорное давление и цитоплазма прижимается к клеточной стенке. Осмотическое поглощение воды играет важную роль при растяжении клеток во время роста, а также в общем водном режиме растения.
2.Иногда в вакуоле присутствуют пигменты, называемые антоцианами. В эту группу входят антоцианы, имеющие красную, синюю или пурпурную окраску, и некоторые родственные соединения, окрашенные в желтый или кремовый цвета. Именно эти пигменты и определяют окраску цветов (например, у роз, фиалок, георгинов), а также окраску плодов, почек и листьев. У последних они обусловливают различные оттенки осенью. Окраска играет роль в привлечении насекомых, птиц и некоторых других животных, уча- ствующих в опылении растений и в распространении семян.
3.У растений в вакуолях иногда содержатся гидролитические ферменты, и тогда при жизни клетки вакуоли действуют как лизосомы. После гибели клетки тонопласт, как и все другие мембраны, теряет свою избирательную проницаемость и ферменты высвобождаются из вакуолей, вызывая автолиз.
4.В вакуолях растения могут накапливаться отходы жизнедеятельности и некоторые вторичные метаболиты. Из отходов иногда
22
обнаруживаются кристаллы оксалата кальция. Роль вторичных продуктов не всегда ясна. Это касается, в частности, алкалоидов, которые могут сохраняться в вакуолях. Возможно, что они, подобно танинам с их вяжущим вкусом, отталкивают травоядных животных, т. е. выполняют защитную функцию. Танины часто встре- чаются в вакуолях (как, впрочем, и в цитоплазме, и в клеточных стенках) листьев, коры, древесины, незрелых плодов и семенных оболочках. Может накапливаться в вакуолях и латекс (млечный сок растений), обычно в виде молочно-белой эмульсии, например млечный сок одуванчика.
Некоторые клетки (млечные клетки) специализируются на выделении млечного сока. Так, в млечном соке бразильской гевеи содержатся ферменты и соединения, необходимые для синтеза каучука, а в млечном соке мака снотворного — алкалоиды.
5. Часть компонентов вакуолярного сока играет роль запасных питательных веществ, при необходимости используемых цитоплазмой. Среди них в первую очередь следует назвать сахарозу, минеральные соли и инулин.
ßäðà имеются во всех эукариотических клетках, за исключе- нием зрелых члеников ситовидных трубок флоэмы и зрелых эритроцитов млекопитающих, и являются наиболее заметной структурой в цитоплазме клетки.
Ядро окружено двойной мембраной, которую часто называют ядерной оболочкой. В ядерной мембране имеются крупные, видимые в электронный микроскоп поры (диаметр 30—100 нм; приблизительно 1000 пор на одно ядро). Диаметр ядра составляет 5—10 мкм. Ядерная пора — это не простое отверстие, а сложная структура, через которую проходят макромолекулы и даже рибосомы. Небольшие аминокислотные молекулы поступают в ядро не путем диффузии через поры, а путем активного транспорта че- рез ядерную мембрану. Ядро необходимо для жизни клетки, поскольку именно оно регулирует всю активность. В клеточном ядре находятся ДНК-содержащие хромосомы и РНК-содержащие ядрышки, погруженные в свободный от нуклеиновых кислот матрикс, называемый нуклеоплазмой.
В форме ДНК в хромосомах содержится генетическая информация, с помощью которой контролируются все клеточные процессы обмена веществ, роста и развития.
Нуклеоплазма (кариоплазма) — внутренняя среда ядра, его матрикс. В матриксе есть зоны клеточного вещества — хроматина, который легко окрашивается щелочными красителями. В состав хроматина входит ДНК и специальные белки — гистоны.
23
Ядро выполняет следующие функции:
1)хранение информации;
2)передача информации от клетки к клетке (деление ядра, клеточное деление, размножение, наследственность) путем синтеза абсолютно идентичной ДНК, в которой закодирована эта информация;
3)передача информации в цитоплазму путем синтеза информационной РНК.
В нуклеоплазме содержатся кислые белки (в противоположность основным гистонам хромосом), среди которых имеются многие ферменты.
Каждое ядро содержит одно или несколько ядрышек, которые можно увидеть под световым микроскопом.
Ядрышки представляют собой округлые образования. В них синтезируются рибосомные РНК. Обычно в ядрах диплоидных организмов имеется два ядрышка, по одному для каждого гаплоидного набора хромосом. Состоят они в основном из белка, ядрышки содержат до 5 % РНК.
Эндоплазматическая сеть (эндоплазматический ретикулум) представляет собой систему связанных полостей, канальцев и трубочек, которые пронизывают всю мезоплазму (от плазмалеммы до тонопласта). Эти полости и каналы окружены мембраной.
К мембранам ЭР может прикрепляться большое количество рибосом — органоидов, которые состоят из белков и РНК приблизительно в одинаковых весовых соотношениях. Такой ЭР полу- чил название шероховатого (гранулярного) в отличие от гладкого (при отсутствии рибосом).
Рибосомы выявлены также в цитозоле, ядре, пластидах и митохондриях. Независимо от местонахождения они выполняют одну и ту же функцию — участвуют в синтезе белков.
Для характеристики молекулярной массы больших молекул, в частности рибосом, используют константы седиментации, которые измеряются с помощью ультрацентрифугирования. Константу седиментации выражают в единицах Сведберга (S), которые для эукариот составляют 80 S, прокариот — 70 S, для митохондрий и хлоропластов — 70 S.
Аппарат Гольджи — термин, который используется для обозначения всех диктиосом в клетке. Диктиосомы — это группы плоских, дисковидных пузырьков, или цистерн, которые по краям разветвляются в сложную систему трубочек. Диктиосомы в клетках высших растений обычно состоят из 4—8 цистерн, собранных вместе. Обычно в пачке цистерн различают формиру-
24

ющуюся и созревающие стороны. Мембраны формирующихся цистерн по структуре напоминают мембраны ЭР, а мембраны созревающих цистерн — плазматическую мембрану.
Диктиосомы участвуют в секреции, а у большинства высших растений — в образовании клеточных оболочек. Полисахариды клеточной оболочки, синтезируемые диктиосомами, накапливаются в пузырьках, которые затем отделяются от созревающих цистерн. Эти секреторные пузырьки мигрируют и сливаются с плазматической мембраной, при этом содержащиеся в них полисахариды встраиваются в клеточную оболочку.
Лизосомы — это овальной формы органоиды клетки (пузырьки диаметром 1 мкм), окруженные мембраной. В них содержится набор ферментов, которые разрушают белки, нуклеиновые кислоты, углеводы, липиды. Ферменты лизосом (гидролитические) расщепляют принесенные пузырьками вещества.
Мембрана лизосом прочная и затрудняет проникновение собственных ферментов в цитоплазму клетки, но когда лизосома повреждается какими-либо внешними факторами, то разрушается вся клетка или часть ее. Лизосомы обеспечивают дополнительным «сырьем» химические и энергетические процессы в клетке.
В типичной клетке имеется множество митохондрий — органелл, в которых происходит дыхание, а точнее синтез АТФ.
Митохондрии (îò ãðå÷. mitos — íèòü è chondros — зернышко) имеют округлую и продолговатую форму диаметром 0,4—0,5 мкм и длиной 1—5 мкм (рис. 1.5).
Ðèñ. 1.5. Строение митохондрии
25
Количество митохондрий варьируется от единиц до 1500—2000 на растительную клетку.
Митохондрии ограничены двумя мембранами: наружной и внутренней, толщина каждой из них 5—6 нм. Наружная мембрана выглядит растянутой, а внутренняя образует складки, называемые гребнями (кристами), различной формы. Пространство между мембранами, в состав которого входит также внутреннее пространство крист, называется межмембранным (перимитохондриальным) пространством. Оно служит средой для внутренней мембраны и матрикса митохондрий.
Митохондрии в целом содержат 65—70 % белка, 25—30 % липидов и небольшое количество нуклеиновых кислот. 70 % от общего содержания липидов составляют фосфолипиды (фосфатидилхолин и фосфатидилэтаноламин). Жирнокислотный состав характеризуется высоким содержанием насыщенных жирных кислот, обеспечивающих «жесткость» мембраны.
В митохондриях локализованы системы аэробного дыхания и окислительного фосфорилирования. В результате дыхания расщепляются органические молекулы и высвобождается энергия с передачей ее на молекулу АТФ.
Митохондрии содержат белки, РНК, тяжи ДНК, рибосомы, сходные с бактериальными, и различные растворенные вещества. ДНК существует в виде кольцевых молекул, располагающихся в одном или нескольких нуклеотидах.
Пластиды, наряду с вакуолями и клеточной оболочкой, — характерные компоненты растительных клеток. Каждая пластида окружена собственной оболочкой, состоящей из двух элементарных мембран. Внутри пластид различают мембранную систему и более или менее гомогенное вещество — строму.
Из пластид наибольшее значение имеют хлоропласты (îò ãðå÷. chloros — зеленый), в которых идет фотосинтез. Внутренняя структура хлоропласта довольно сложна. Строма пронизана развитой системой мембран, имеющих форму плоских пузырьков, называемых тилакоидами. Тилакоиды собраны в стопки — граны, напоминающие столбики монет (рис. 1.6).
Хлоропласты, в которых протекает фотосинтез, содержат хлорофиллы и каротиноиды. Размер — 4—5 мкм. В одной клетке мезофилла листа может содержаться 40—50 хлоропластов, в 1 мм2 листа — около 500 000. В цитоплазме хлоропласты обычно располагаются параллельно клеточной оболочке.
26
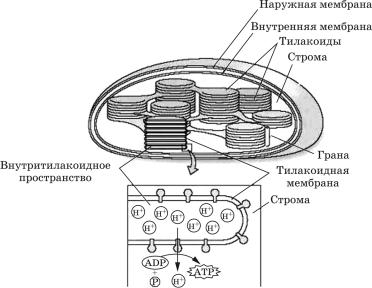
Ðèñ. 1.6. Схема строения хлоропласта
Хлорофиллы и каротиноиды встроены в тилакоидные мембраны. Хлоропласты зеленых растений и водорослей часто содержат зерна крахмала и мелкие липидные (жировые) капли. Крахмальные зерна — это временные хранилища продуктов фотосинтеза. Они могут исчезнуть из хлоропластов, находящихся в темноте, всего лишь за 24 часа и появиться вновь уже через 3—4 часа после переноса растений на свет.
В изолированных хлоропластах осуществляется синтез РНК, который обычно контролируется только хромосомной ДНК. Образование хлоропластов и синтез находящихся в них пигментов в значи- тельной степени контролируются хромосомной ДНК, малопонятным образом взаимодействующей с ДНК хлоропластов. Тем не менее в отсутствие собственной ДНК хлоропласты не формируются.
Хлоропласты могут считаться основными клеточными органеллами, поскольку первыми стоят в цепи преобразования солнечной энергии, в результате этих преобразований мы получаем пищу и топливо. В хлоропластах протекает не только фотосинтез. Они участвуют в синтезе аминокислот и жирных кислот, служат хранилищем временных запасов крахмала.
27
Хромопласты (îò ãðå÷. ñhroma — öâåò è plastes — образующий) — пигментированные пластиды. Многообразные по форме хромопласты не содержат хлорофилла, но синтезируют и накапливают каротиноиды, которые придают желтую, оранжевую и другую окраску. Корнеплоды моркови, плоды томатов окрашены пигментами, которые находятся в хромопластах.
Лейкопласты (îò ãðå÷. leukos — белый и plastes — образующий) являются местом накопления запасного вещества — крахмала. Особенно много лейкопластов в клетках клубней картофеля. На свету лейкопласты могут преобразовываться в хлоропласты (клубни картофеля зеленеют). Осенью хлоропласты преобразуются в хромопласты — зеленые листья и плоды желтеют и краснеют.
В отличие от пластид и митохондрий, которые ограничены двумя мембранами, микротельца представляют собой сфериче- ские органеллы, окруженные одной мембраной. Их диаметр колеблется от 0,5 до 1,5 мкм. Микротельца имеют гранулярное содержимое, иногда в них встречаются и кристаллические белковые включения. Микротельца обычно связаны с одним или двумя участками эндоплазматического ретикулума.
Некоторые микротельца, называемые пероксисомами, играют важную роль в метаболизме глиоксиловой кислоты, имеющей непосредственное отношение к фотодыханию. В зеленых листьях пероксисомы связаны с митохондриями и хлоропластами. Другие микротельца, называемые глиоксисомами, содержат ферменты, необходимые для превращения жиров в углеводы, что происходит во многих семенах во время прорастания.
Все известные одноклеточные и многоклеточные организмы разделяются на две большие группы — прокариоты и эукариоты. К первым относятся бактерии и сине-зеленые водоросли (цианобактерии), ко вторым — зеленые растения, в том числе и остальные водоросли. Клетки прокариот (от лат. pro — äî è ãðå÷. karyon — ядро) не имеют ядра, т. е. генетический материал (ДНК) прокариот находится в цитоплазме (отсутствует ядерная мембрана).
У эукариот (от греч. eu — настоящий, действительный и karyon — ядро), наоборот, есть ядро, таким образом, у них генетиче- ский материал ограничен двойной мембраной (ядерной оболоч- кой) и образует отдельную клеточную структуру, которую легко узнать. Прокариоты отличаются от эукариот по целому ряду признаков.
Клетки эмбриональных тканей животных и растений в общем плане строения очень сходны, если не учитывать особенностей ор-
28
ганизации клеточной стенки у растений. Морфологические различия, связанные с образом жизни и способом питания, проявляются уже в дифференцированных клетках специализированных тканей растений и животных.
Вещества, составляющие пищу растений (углекислый газ, минеральные элементы и вода), находятся вокруг растения в рассеянном виде. Зеленые растения на свету осуществляют автотрофный способ питания.
Эволюционно сложились некоторые особенности строения и роста растительных клеток. К ним относятся:
1)прочная полисахаридная клеточная стенка, окружающая клетку;
2)пластидная система (хлоропласты), возникшая в связи с указанным типом питания;
3)крупная центральная вакуоль, в зрелых клетках играющая важную роль в поддержании тургорного давления клеток;
4)плазмодесмы — узкие каналы, обеспечивающие практиче- ски во всем растении непрерывность протоплазмы и возможность диффузии малых молекул из клетки в клетку;
5)наличие особого типа роста — роста растяжением. Кроме того, у делящихся растительных клеток нет центриолей.
1.2. Мембранный принцип организации протопласта и органоидов
Основу живой материи составляют высокомолекулярные органические вещества — белки, нуклеиновые кислоты и некоторые другие. Их называют биополимерами. Однако «оживают» биополимеры лишь тогда, когда они в определенном порядке располагаются в пространстве, определенным образом взаимодействуют друг с другом. Основные биологические структуры, обеспе- чивающие такое взаимодействие, называют биомембранами.
Мембраны составляют не только наружную оболочку, многие клетки буквально начинены сложной системой внутренних мембран.
Клеточные мембраны играют важную роль по ряду причин. Они отделяют клеточное содержимое от внешней среды, регулируют обмен между клеткой и средой, делят клетки на отсеки, или компартменты, предназначенные для тех или иных специализированных метаболических путей. Некоторые химические реакции, в частности световые реакции фотосинтеза в хлоропластах
29
или окислительное фосфорилирование при дыхании в митохондриях, протекают на самих мембранах. На мембранах же располагаются рецепторные участки для распознания стимулов, поступающих из среды или из другой части самого организма. Знакомство со свойствами клеточных мембран необходимо для понимания функционирования клетки в целом.
Становление наших представлений о строении и функционировании биологических мембран неразрывно связано с прогрессом технических средств, используемых биологами в экспериментах. Началось все с первых экспериментов А. Левенгука, смастерившего в 1674 г. простейший микроскоп. Однако понадобилось более 150 лет, чтобы сформировать принципы устройства клеток животных и растений, и более 200 лет, чтобы биологическая мембрана заняла одно из центральных мест среди внутриклеточных структур.
Введение в практику электронного микроскопа позволило подойти к решению вопросов, связанных с внутриклеточными структурами.
Мембраны способны пропускать воду и другие малые молекулы; через них диффундируют глюкоза, аминокислоты, жирные кислоты и ионы, причем сами мембраны активно регулируют этот процесс — одни вещества пропускают, другие нет.
Ранние работы Э. Овертона и Р. Колландера в конце ХIХ и на- чале ХХ вв. по измерению проницаемости мембран показали, что органические растворители, например спирт, эфир, хлороформ, проникают через мембраны даже быстрее, чем вода. Это свидетельствует о том, что в мембранах есть какая-то неполярная часть; иными словами, как оказалось, мембраны содержат липиды. Позже данное предположение удалось подтвердить химиче- ским анализом.
Основной строительный материал мембран — белки и липиды. Весовое соотношение их: примерно 40 % липидов и 60 % белков. Однако у некоторых мембран это соотношение может быть и иным.
В биологических мембранах липиды представлены главным образом фосфолипидами и стероидами. Все мембранные липиды обладают выраженной амфифильностью, т. е. в их структуре представлена полярная часть, обладающая сродством к воде, и неполярная, не растворяющаяся в воде. Именно благодаря этому липиды в воде образуют мицеллы — замкнутые структуры, в которых полярные группы выставлены наружу, а неполярные углеродные цепи спрятаны внутрь и, контактируя друг с другом, пря- чутся от воды.
30

По-видимому, липидный состав мембраны успел сформироваться уже на ранних этапах биологической эволюции, о чем свидетельствует принципиальная близость набора липидов у разных организмов.
Второй компонент мембран представлен белками. Белки — высокомолекулярные соединения, относящиеся к классу линейных биополимеров. Они состоят из отдельных элементарных единиц, в роли которых выступает 20 различных аминокислот. Аминокислоты имеют общую структурную форму:
Все аминокислоты амфолиты, объединяющие в себе положительно заряженную аминогруппу NH3+ и отрицательно заряженную группу СОО–. Заряженность аминокислот контролируется концентрацией водородных ионов в среде (рН) — в кислых средах господствует положительный заряд (NH3+ , СООН), в нейтральных — нулевой (NH3+ , ÑÎΖ), в щелочных — отрицательный (NH2, ÑÎΖ). Но в физиологическом интервале рН (6—8) аминокислоты имеют и отрицательный, и положительный заряды.
С участием амино- и карбоксильных групп происходит полимеризация аминокислот, т. е. образование полипептидной цепи.
Молекулярная масса мембранных белков колеблется между 12 000 и 500 000 дальтон. По силе сцепления с мембраной белки разделяют на два класса — периферические и интегральные.
Периферические слабо связаны с мембраной и отделяются от нее при самых различных воздействиях: многократной промывкой водой или гипертоническими растворами солей, слабыми детергентами и ультразвуком. Эти белки не вступают в тесное взаимодействие с липидами и отделяются от мембраны в чистом виде. По аминокислотному составу периферические белки напоминают многие немембранные.
Более липофильные интегральные белки, составляющие свыше половины общего фонда, прочно связаны с липидным бислоем. Их удается выделить с помощью детергентов и малополярных растворителей, причем обязательно в комплексе с липидом. Лишившись липидной рубашки, интегральные белки теряют активность и немедленно агрегируют.
В мембранах присутствуют и углеводы, ковалентно связанные либо с белками (гликопротеиды), либо с липидами (гликолипи-
31