

 ных,
как правило, одной копией. Большая часть
рибосомальных белков
выполняет структурную функцию.
ных,
как правило, одной копией. Большая часть
рибосомальных белков
выполняет структурную функцию.
Синтез белка осуществляется агрегатами, состоящими из рибосом, молекул информационной и транспортных РНК и называемыми полирибосомами, или полисомами. Последние могут находиться в цитоплазме или же быть связанными с мембранными структурами.
Генетический аппарат и репликация хромосомы
Строение генетического аппарата прокариох долгое время было предметом жарких дискуссий, суть которых сводилась к тому, есть у них такое же ядро, как у эукариот, или нет. Установлено, что генетический материал прокариотных организмов, как и эукариотных, представлен ДНК, но имеются существенные различия в его структурной организации. У прокариот ДНК представляет собой более или менее компактное образование, занимающее определенную область в цитоплазме и не отделенное от нее мембраной, как это имеет место у эукариот. Чтобы подчеркнуть структурные различия в генетическом аппарате прокариотных и эукариотных клеток, предложено у первых его называть нуклеоидом в отличие от ядра у вторых.
При электронно-микроскопическом наблюдении видно, что нуклеоид прокариот, несмотря на отсутствие ядерной мембраны, довольно четко отграничен от цитоплазмы, занимает в ней, как правило, центральную область и заполнен нитями ДНК диаметром около 2 нм. Не исключено, что на выявляемую в электронном микроскопе организацию прокариотной хромосомы большое влияние оказывают условия фиксации препарата. По имеющимся наблюдениям, в живой клетке нуклеоид занимает больше места в цитоплазме.
Вся генетическая информация прокариот содержится в одной молекуле ДНК, имеющей форму ковалеытно замкнутого кольца и получившей название бактериальной хромосомы10. Длина молекулы в развернутом виде может составлять более 1 мм, т. е. почти в 1000 раз превышать длину бактериальной клетки. Длительное время считали, что в распределении нитей ДНК бактериальной хромосомы не прослеживается никакой закономерности. Однако если исходить из того, что молекула ДНК образует беспорядочный клубок, трудно объяснить процесс репликации и последующее распределение образовавшихся хромосом по дочерним клеткам. Специальные исследования показали, что хромосомы прокариот представляют собой высокоупорядоченную структуру, имеющую константу седиментации 1300—2000S для свободной и 3200—70005 для
10 В прокариотной клетке ДНК может находиться м вне бактериальной хромосомы в плазмидах, но последние не являются обязательными клеточными компонентами.

Рис. 16. Строение ДНК:
А — фрагмент нити ДНК, образованной чередующимися остатками дезоксирибозы и фосфорной кислоты. К первому углеродному атому дезоксирибозы присоединено азотистое основание: / — цитозин; 2 — гуанин; Б — двойная спираль ДНК: Д ■— дезоксирибоза; Ф — фосфат; А — аденин; Т — тимин; Г ■— гуанин; Ц — цитозин
связанной с мембраной формы. В том и другом случае часть ДНК в этой структуре представлена системой из 20—100 независимо суперспирализованных петель. В обеспечении супер-спирализованной организации хромосом участвуют молекулы РНК.
Хромосомы большинства прокариот имеют молекулярную массу в пределах 1—3-Ю9 Да. В группе микоплазм генетический материал представлен молекулами, имеющими наименьшее для клеточных организмов количество ДНК (0,4—0,8• 109), а наибольшее содержание ДНК обнаружено у нитчатых ци-анобактерий (8,5■ 109). Хотя каждая прокариотная клетка содержит 1 хромосому, часто в экспоненциально растущей культуре количество ДНК на клетку может достигать массы 3, 4, 8 и более хромосом. Нередко в клетках при действии на Них определенных факторов (температуры, рН среды, ионизирующего излучения, солей тяжелых металлов, некоторых анти-
54
55
биотиков и др.) происходит образование множества копий хромосомы. При устранении воздействия этих факторов, а также после перехода в стационарную фазу в клетках, как правило, обнаруживается по одной копии хромосомы11.
ДНК прокариот построена так же, как и зукариот (рис. 16). Молекула ДНК несет множество отрицательных зарядов, поскольку каждый фосфатный остаток содержит ионизированную гидроксильную группу. У эукариот отрицательные заряды нейтрализуются образованием комплекса ДНК с основными белками — гистонами. В клетках подавляющего большинства прокариот не обнаружено гистонов, поэтому нейтрализация зарядов осуществляется взаимодействием ДНК с полиаминами (спермином и спермидином), а также с ионами Mg2+. В последнее время у некоторых архебактерий и цианобактерий обнаружены гистоны и гистоноподобные белки, связанные с ДНК. Содержание пар оснований A-f-T и Г+Ц в молекуле ДНК является постоянным для данного вида организма и служит важным диагностическим признаком. У прокариот молярная доля ГЦ в ДНК колеблется в очень широких пределах: от 23 до 75%.
Деление молекулы ДНК (репликация) происходит по полуконсервативному механизму и в норме всегда предшествует делению клетки. С помощью электронного микроскопа установлено, что репликация ДНК начинается в точке прикрепления кольцевой хромосомы к ЦПМ, где локализован фермента-
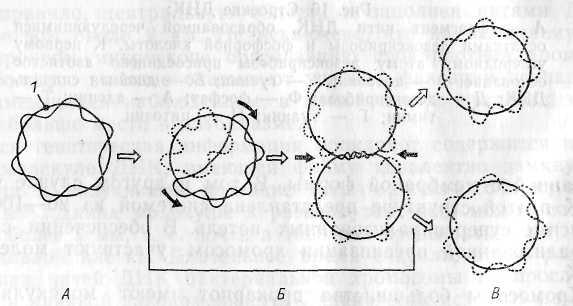
Рис. 17. Репликация кольцевой бактериальной хромосомы в
двух направлениях.
А — родительская молекула ДНК; Б — промежуточные ре-пликативные формы; В — дочерние молекулы ДНК после завершения процесса репликации и расхождения: ) — точка начала репликации; черными стрелками показано направление репликации
11
Из изложенного выше следует, что термины
«нуклеоид» и «хромосома»
не всегда совпадают. В зависимости от
условий нуклеоид прокариотной клетки
может состоять из одной или некоторого
числа копий хромосомы.
56
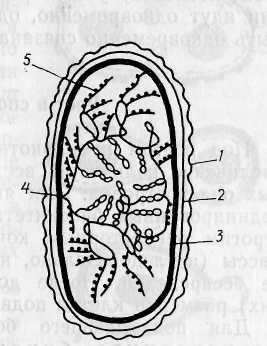
тивный аппарат, ответственный за репликацию. Часто можно обнаружить, что контакт ДНК с ЦПМ осуществляется посредством мезосом. Репликация, начавшаяся в точке прикрепления, идет затем в двух противоположных направлениях, образуя ха-

Рис. 19. Модель организации нуклеоида Е. coli:
— наружная мембра на клеточной стенки;
— пептидогликановый слой; 3 — ЦПМ; 4 — точка прикрепления бак териальной хромосомы к ЦПМ; 5 — рибосомы, «сидящие» на иРНК; остальные объяснения см. в тексте (по Громо ву, 1985)
. Рис. 18. Механизм распределения
бактериальных хромосом: А — бактериальная клетка содержит частично реплицированную хромосому, прикрепленную к мембране в точке (или точках) репликации; Б —■ репликация хромосомы завершена. В бактериальной клетке две дочерние хромосомы, каждая из которых прикреплена к ЦПМ. Показан синтез клеточной стенки и ЦПМ; В ■— продолжающийся синтез мембраны и клеточной стенки приводит к разделению дочерних хромосом. Показано начало деления клетки путем образования поперечной перегородки: 1 — ДНК; 2 — прикрепление хромосомы к ЦПМ; 3 — ЦПМ; 4 — клеточная стенка; 5 — синтезированный участок ЦПМ; 6 — новый материал клеточной стенки
рактерные для кольцевой хромосомы промежуточные структуры (рис. 17). Возникающие дочерние хромосомы остаются прикрепленными к мембране. .Репликация молекул ДНК происходит параллельно с синтезом мембраны в области контакта ДНК с ЦПМ. Это приводит к разделению (сегрегации) дочерних молекул ДНК и оформлению обособленных хромосом (рис. 18).
Модель строения бактериальной хромосомы должна объяснять также прохождение в клетке процессов транскрипции и трансляции. Согласно существующим представлениям супер-спирализованные петли соответствуют неактивным в данное время участкам ДНК и находятся в центре нуклеоида. По его периферии располагаются деспирализованные участки, на которых происходит синтез информационной РНК (иРНК), при этом, поскольку у бактерий процессы транскрипции и трансляции идут одновременно, одна и та же молекула иРНК может быть одновременно связана с ДНК и рибосомами (рис. 19).
Рост и способы размножения
Под ростом прокариотной клетки понимают согласованное увеличение количества всех химических компонентов, из которых она построена. Рост является результатом множества скоординированных биосинтетических процессов, находящихся под строгим регуляторным контролем, и приводит к увеличению массы (а следовательно, и размеров) клетки. Но рост клетки не беспределен. После достижения определенных (критических) размеров клетка подвергается делению.
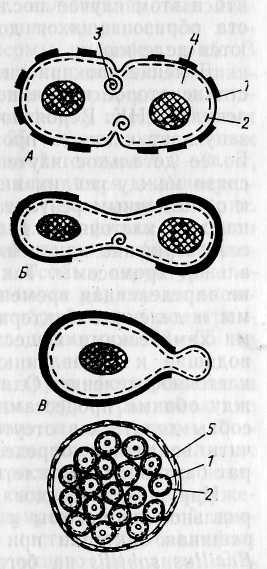
Для подавляющего большинства прокариот характерно равновеликое бинарное поперечное деление, приводящее к образованию двух одинаковых дочерних клеток. При таком способе деления имеет место симметрия в отношении продольной и поперечной оси. У большинства грамположи-тельных эубактерий и нитчатых цианобактерий деление происходит путем синтеза поперечной перегородки, идущего от периферии к центру (рис. 20, Л). Так у Bacillus subtilis в середине клетки сначала имеет место кольцевое впячивание ЦПМ, сопровождающееся формированием мезосом разного внешнего вида. Они образуются в месте закладки поперечной перегородки, и предполагается их активное участие в процессах синтеза пептидогликана и других компонентов клеточной стенки. Поперечная перегородка формируется из ЦПМ и пептидоглика-нового слоя, ее наружные слои синтезируются позднее. Клетки большинства грамотрицательных эубактерий делятся путем перетяжки. У Е. coli на месте деления обнаруживается постепенно увеличивающееся и направленное внутрь искривление ЦПМ и клеточной стенки (рис. 20,5). Синтез новой клеточной стенки может происходить в нескольких местах или только в зоне формирования поперечной перегородки (рис. 20, А, Б).
Вариантом бинарного деления является почкование, которое можно рассматривать как неравновеликое бинарное деление. При почковании на одном из полюсов материнской клетки образуется маленький вырост (почка), увеличивающийся в процессе роста. Постепенно почка достигает размеров материнской клетки, после чего отделяется от последней. Клеточная
58
стенка почки полностью синтезируется заново (рис. 20, В). В процессе почкования симметрия наблюдается в отношении только продольной оси. При равновеликом бинарном делении материнская клетка, делясь, дает начало двум дочерним клеткам и сама, таким образом, исчезает. При почковании мате-
ринская клетка дает начало дочерней клетке, и между ними можно в большинстве случаев обнаружить морфологические и физиологические различия: есть старая материнская клетка и новая дочерняя. В этом случае можно наблюдать процесс старения. Так, для некоторых штаммов Rhodotnicrobium показано, что материнская клетка способна отпочковывать не более 4 дочерних клеток. Дочерние клетки лучше приспосабливаются к меняющимся условиям. Почкование обнаружено в разных группах прокариот: среди фото- и хемотрофов, осуществляющих
Рис. 20. Способы деления и синтез клеточной
стенки у прокариот:
А •— деление путем образования поперечной перегородки; Б — деление путем перетяжки; В — почкование; Г — множественное деление: 1 — клеточная стенка (толстой линией ■обозначена клеточная стенка материнской клетки, тонкой — заново синтезированная); 2 — ЦПМ; 3 — мембранная структура; 4 ■— цитоплазма, в центре которой расположен нуклеоид; 5 ■— дополнительный фибриллярный слой клеточной стенки
авто- и гетеротрофный конструктивный метаболизм. Вероятно, оно в процессе эволюции возникало несколько раз.
Бинарное деление может происходить в одной или нескольких плоскостях. В первом случае, если после деления клетки не расходятся, это приводит к образованию цепочек палочковидных или сферических клеток, во втором — к клеточным скоплениям разной формы (см. рис. 3,4—6). Расхождение образовавшихся дочерних клеток происходит в результате лизиса среднего слоя клеточной стенки.
Для одной группы одноклеточных цианобактерий описано размножение путем множественного деления. Оно начинается с предварительной репликации хромосомы и увеличения размеров вегетативной клетки, которая затем претерпевает ряд х быстрых последовательных бинарных делений, происходящих внутри дополнительного фибриллярного слоя материнской клеточной стенки. Это приводит к образованию мелких клеток, по-
59
лучивших название баеоцитов 12, число которых у разных вадов колеблется от 4 до 1000. Освобождение баеоцитов происходит путем разрыва материнской клеточной стенки (рис. 20, Г). Таким образом, в основе множественного деления лежит принцип равновеликого бинарного деления. Отличие заключается в том, что в этом случае после бинарного деления не происходит роста образовавшихся дочерних клеток, а они снова подвергаются делению.
Деление прокариотной клетки начинается, как правило, спустя некоторое время после завершения цикла репликации молекулы ДНК- Вероятно, репликация бактериальной хромосомы запускает какие-то процессы, ведущие к клеточному делению. Более детальное изучение у разных видов прокариот взаимосвязи между репликацией ДНК и делением клетки не привело, к однозначным результатам. Получены данные о том, что сиг-* налом к клеточному делению служит начало репликации ДНК, ее завершение или репликация определенного локуса бактериальной хромосомы. Таким образом, в норме существует вполне определенная временная связь между репликацией хромосомы и делением бактериальной: «летки. Воздействия различны-: ми химическими веществами и физическими факторами, приводящие к подавлению репликации ДНК, останавливают и клеточное деление. Однако при некоторых условиях связь между обоими процессами может быть нарушена, и клетки способны делиться в отсутствие синтеза ДНК- Это удалось получить введением определенных мутаций в генетический аппарат бактериальной клетки.
Нарушить последовательность процессов репликации бактериальной хромосомы и клеточного деления также можно, выращивая бактерии при разной температуре. Культивирование Bacillus sublilis на богатой питательной среде при 37° приводит к интенсивному делению бактериальной хромосомы и росту клеток, в результате чего в культуре образуются нитевидные клетки, содержащие множество хромосомных копий с отсутствующими совсем или недосформированными (незамкнутыми) поперечными перегородками. При замедлении скорости роста наблюдается деление нитевидных клеток, приводящее к образованию бактериальных клеток нормальной длины.
Внутрицитоплазматические включения
В цитоплазме прокариот обнаруживаются различные включения. Одни из них следует рассматривать как активно функционирующие структуры, другие — как продукты клеточного метаболизма, не выделяющиеся наружу, но откладывающиеся внутри клетки. Некоторые цитоплазматические включения име-
12
Баеоцит — по-гречески маленькая клетка.
60
ют явно приспособительное значение. И наконец, многие из них являются запасными веществами, отложение которых клеткой происходит в условиях избытка питательных веществ в окружающей среде, а потребление наблюдается, когда организм попадает в условия голодания.
К числу внутрицитоплазматических включений, выполняющих определенную функцию в фотосинтезе, относятся хлоросомы зеленых бактерий и фикобилисомы цианобактерий. В этих структурах локализованы пигменты, поглощающие кванты света и передающие их в реакционные центры, т. е. выполняющие роль антенны. Хлоросомы имеют форму продолговатых пузырьков длиной 90—150 и шириной 25—70 нм, окруженных однослойной электронно-плотной мембраной толщиной 2—3 нмг построенной только из белка. Они располагаются в непосредственной близости от ЦПМ, плотно к ней примыкая (см. рис. 4). В хлоросомах локализованы бактериохлорофиллы с, d или е. Водорастворимые пигменты белковой природы (фикобили-протеины) цианобактерий содержатся в особых структурах — фикобилисомах, расположенных правильными рядами на внешних поверхностях фотосинтетических мембран и под электронным микроскопом имеющих вид гранул диаметром 28—55 нм (см. рис. 4).
В клетках некоторых прокариот из групп фототрофных и хемолитотрофных эубактерий содержатся структуры, имеющие форму многогранника с 4—6 сторонами и диаметром 90— 500 нм, получившие название карбоксисом, или полиэдральных тел (см. рис. 4). Под электронным микроскопом удалось показать, что они заполнены гранулярным содержимым и окружены однослойной мембраной белковой природы толщиной примерно 3 нм. Карбоксисомы состоят из частиц рибулозодифосфаткарбоксилазы, • фермента, катализирующего фиксацию СО2 на рибулозодифосфате в восстановительном пентозофосфатном цикле. До настоящего времени окончательно не выяснено, в^ какой форме находится фермент в карбоксисомах: в инертном пли функционирующем состоянии. Имеются данные в пользу того, что в активно растущей культуре больше фермента находится в растворимой форме. При переходе в стационарную фазу увеличивается доля рибулозодифосфаткарбоксилазы в составе карбоксисом. Эти данные указывают на возможную роль карбоксисом как структур, обеспечивающих защиту фермента от воздействия внутриклеточными протеазами и, таким образом, его консервирование.
Примером внутрицитоплазматических включений, имеющих приспособительное значение, служат магнитосомы и газовые вакуоли, или аэросомы, обнаруженные у водных прокариот. Газовые вакуоли найдены у представителей, относящихся к 15< таксономическим группам. Это сложно организованные структуры, напоминающие пчелиные соты (см. рис. 4). Состоят из: множества регулярно расположенных газовых пузырьков, име-
61
азоту указывают на то, что они в клетке служат резервом азота, мобилизуемым при его недостатке в среде.
Для прокариот, метаболизм которых связан с соединениями серы, характерно отложение в клетках молекулярной серы. Сера накапливается, когда в среде содержится сероводород, и окисляется до сульфата, когда весь сероводород среды оказывается исчерпанным. Для аэробных тионовых бактерий, окисляющих H2S, сера служит источником энергии, а для анаэробных фотосинтезирующих серобактерий она является донором электронов.
Обращает внимание, что все запасные вещества представлены в виде высокомолекулярных полимерных молекул, в ряде случаев отграниченных от цитоплазмы белковой мембраной, т. е. находятся в осмотически неактивном состоянии. Это важно, так как в противном случае сосредоточение в цитоплазме «большого числа молекул осмотически активных веществ оказало бы на клетку отрицательное действие.
Глава 5
МОРФОЛОГИЧЕСКАЯ ДИФФЕРЕНЦИРОВКА И УРОВНИ КЛЕТОЧНОЙ ОРГАНИЗАЦИИ ПРОКАРИОТ
Для эволюции прокариотных организмов характерен ярко выраженный физиолого-биохимический уклон, т. е. основное развитие прокариот шло по линии формирования и опробования различных функций, результатом чего и явилось сегодняшнее многообразие типов жизни в микромире. Поразительное физиологическое разнообразие прокариот сформировано на базе весьма ограниченного числа морфологических форм. Действительно, морфологическая эволюция прокариот прошла незначительный путь, так что мы можем говорить лишь о зачатках морфологической дифференцировки на базе прокариотной .клеточной организации. Относительно «продвинутыми» в этом направлении оказались только эубактерий. Для архебактерий характерно отсутствие сложных морфологических форм и какой-либо клеточной дифференцировки.
Вегетативные клетки многих эубактерий в определенных условиях дают начало структурам, морфологически отличающимся от исходных. Ими могут быть вегетативные клетки, но измененной формы, клеточные структуры с четко выраженной функциональной специализацией, различные многоклеточные образования. В подавляющем большинстве случаев все известные проявления морфологической дифференцировки эубактерий направлены на повышение их выживаемости. Это выражается как в формировании специальных клеток, обладающих повышенной устойчивостью к перенесению неблагоприятных условий (эндоспоры, цисты), так и в формировании структур, обеспечивающих
64
эффективное размножение вида (гормогонии и баеоциты циа-|нобактерий).
В основе морфологической дифференцировки лежат определенные биохимические процессы, которые в свою очередь являются выражением соответствующей генетической информации 13. Последняя запрограммирована в генетическом аппарате клетки и реализуется в процессе ее развития или же в зависимости от действия различных внешних факторов.
МОРФОЛОГИЧЕСКИ ДИФФЕРЕНЦИРОВАННЫЕ КЛЕТКИ
Образование морфологически дифференцированных клеток у разных представителей эубактерий суммировано в табл. 6. Большинство таких структур относится к категории покоящихся форм, назначение которых — обеспечить переживание вида в течение длительного времени в неблагоприятных условиях. Это
Таблица 6 Морфологически дифференцированные клетки эубактерий
|
Специализированные клетки |
Представители эубактерий, | у которых они обнаружены |
|
Эндоспоры |
Bacillus, Clostridium, Desulfotomaculum, Sporosarcina, некоторые актиномицеты |
|
Экзоспоры |
некоторые виды Methylosinus, Rhodo-microbium, многие актиномицеты |
|
Цисты |
миксобактерии, скользящие бактерии, Azotobacter, Bdellovibrio |
|
Бактероиды |
клубеньковые бактерии |
|
Гетероцисты, акинеты, баеоциты, гормогонии |
цианобактерии |
эндоспоры ряда грамположительных бактерий, цисты азотобактера и миксобактерии, акинеты цианобактерии, экзоспоры отдельных представителей метилотрофных и фототрофных бактерий, экзо- и эндоспоры актиномицетов. После попадания в подходящие условия покоящиеся формы прорастают, давая начало вегетативным клеткам. Другие морфологически дифференцированные клетки служат для размножения. К ним относятся, например, гормогонии и баеоциты цианобактерии. Наконец, третьи (гетероцисты цианобактерии, бактероиды клубеньковых бактерий) связаны с осуществлением уникального процесса, свойственного только прокариотным организмам, — фиксацией молекулярного азота атмосферы.
13
Ф. Жакоб (F.
Jacob)
и Ж. Моно (J.
Monod)
определили дифферен-Цировку
следующим образом: «одну клетку следует
считать дифференцированной
по сравнению с другой, если при одинаковых
геномах набор белков,
синтезированных в этих клетках, различен».
65
3 Микробиология
