

Мекк мек »► erk
МАР-киназный каскад JNK/SAPK-киназный каскад HOG/рЗв-киназный каскад
Raf, Mos, MEKK1
МЕК1.2
ERK1.2
MEKK1
\
JNKK
I
JNK/SAPK
MEKK1
\
JNKK.MKK3
I
HOG/D38
MAP — киназные модули млекопитающих; ERK — extracellular signal-regulated kinase; MKK — MAP kinase/ERK kinase; МЕК — то же, что и МАРКК; МЕКК — МЕК kinase; JNK/SAPK — Jun-N-terminal kinase/stress-activated protein kinase; JNKK — JNK kinase. Пояснения в тексте.
щие один и тот же путь, способны вызывать абсолютно разные клеточные реакции. Так, активация Ras в клетках линии PC 12 эпидермальным фактором роста вызывает клеточную пролиферацию, а фактором роста нервов — дифференцировку.
Разнообразие сигналов, использующих МАР-киназный путь, свидетельствует о том, что он «служит» разным целям. Конкретные же последствия его активации (иногда диаметрально противоположные) определяются той конкретной ситуацией, в которой оказалась данная клетка. Вместе с тем важно подчеркнуть, что этот сигнальный путь, как и другие, не «линеен», а входит как элемент в сложную сеть конвергирующих и ветвящихся информационных потоков.
Наиболее полно изученный и состоящий из 3 ферментов МАР-киназный каскад является каноническим для ряда других 3-киназных каскадов, обнаруженных в клетках млекопитающих и обладающих иными внутриклеточными мишенями (схема 4.1). Согласно концепции трехкомпонентных МАР-ки-назных модулей, они, будучи во многих отношениях родственными, «обслуживают» разные рецепторы и образуют параллельные информационные потоки. Наряду с каноническим МАР-киназным каскадом (МАРККК -> МАРКК -> МАРК) существуют трехкомпонентные JNK/SAPK (Jun-N-terminal kinase/stress-activated protein kinase) и HOG/p38 каскады (одни и те же киназы известны иногда под разными названиями, что объясняется их независимым обнаружением в разных лабораториях).
Активность ферментов, участвующих в киназных каскадах, уравновешивается активностью противодействующих им и находящихся под столь же строгим контролем фосфатаз.
13—1385
193
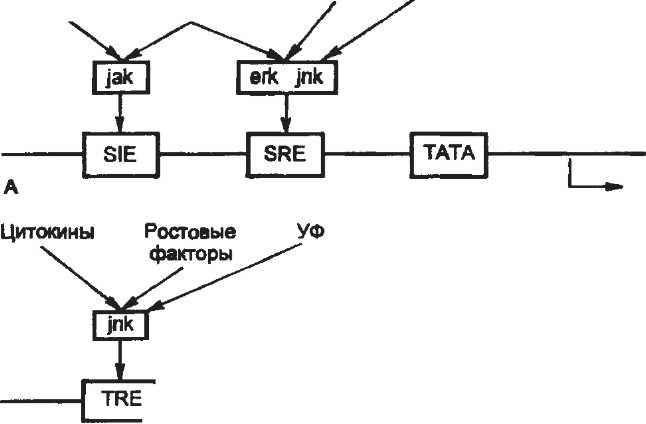
Активация транскрипции. Группа генов, определяющих вхождение клетки в фазу S, активируется транскрипционным фактором АР-1, представляющим собой комплекс белков — членов семейств Jun и Fos (гены, их кодирующие, — c-jun и c-fos, относятся к числу протоонкогенов; с — от cell, обозначает их клеточное происхождение в отличие от вирусных онкогенов v-jun и v-fos). Эти транскрипционные факторы могут взаимодействовать между собой с образованием множества гомо- и гетеродимеров, связывающихся с определенным участком ДНК и стимулирующих синтез РНК на прилежащих к этим участкам генов.
Регуляторные участки (прямоугольники), прилежащие к генам c-fos и c-jun, взаимодействуют с транскрипционными факторами (на схеме не представлены), активированными соответствующими протеинкиназами (последние принадлежат к различным киназным каскадам). SIE (sis-induc-ible enhancer), SRE (serum response element), TRE (TPA response element) — регуляторные участки генов; TATA (так называемый TATA-box) — короткая нуклеотидная последовательность, играющая важную роль в формировании транскрипционного комплекса; Jak, Jnk, Erk — последние члены разных киназных каскадов; А — активация c-fos (стрелка показывает начало считывания РНК); Б — активация c-jun.
|
1 1 |
TATA |
|
|
J 1 |
1 |
МАР-киназы повышают активность АР-1 двояким образом: а) опосредованным, активируя соответствующие гены и увеличивая тем самым содержание комплекса в клетке, и б) прямым, фосфорилируя входящие в их состав сериновые и треониновые остатки. В первом случае (схема 4.2) МАР-киназы фосфорилируют в строго определенных сайтах соответст-
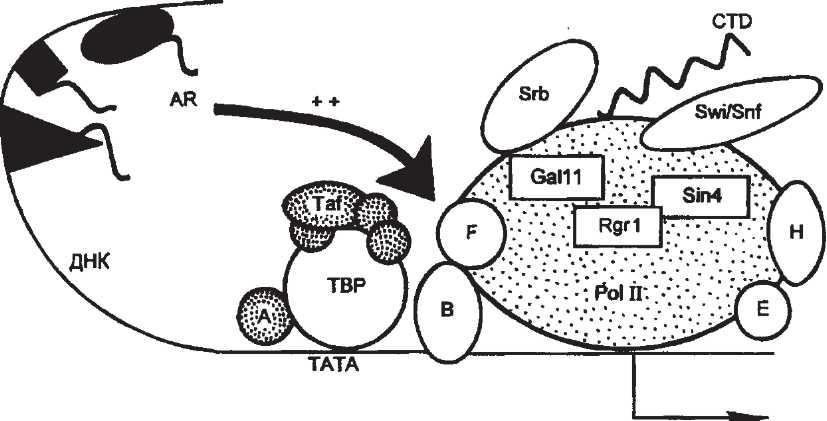
Рис.
4.9. Транскрипционная триада — типичный
эукариотический транскрипционный
комплекс в промоторном участке гена.
Белки-активаторы (Аг) «садятся» на
участок энхансера (регуляторная
последовательность ДНК, усиливающая
транскрипцию) и стимулируют (++) синтез
РНК посредством белок-белкового
взаимодействия своих активирующих
доменов (волнистая линия) с компонентами
базового аппарата транскрипции. Последний
состоит из двух основных компонентов
— ТВР
(TATA-binding protein)
и РНК-полимеразы II
(Pol II).
С ТВР взаимодействует около 10
дополнительных
белков —
Taf (TBP-associated factors)
и транскрипционный фактор
TFIIA
(А). В состав комплекса с РНК-полимеразой
входят транскрипционные факторы
TFIIB
(В),
TFIIE
(Е),
TFIIF (F), TFIIH
(Н) и ряд дополнительных белков,
способствующих правильной посадке
комплекса и точной транскрипции, —
Srb, Gall
1,
Sin4, RgrI,
а также мультифермент-ный комплекс
Swi/Snf
(ДНК-стимулируемая АТФаза), вызывающий
разрушение структуры нуклеосом и
движение РНК-полимеразы по нити хроматина.
вующие транскрипционные факторы (в частности, TCF — ternary complex factor). Последний взаимодействует с так называемым базовым аппаратом транскрипции — мультифермент-ным комплексом, формирующимся на промоторном участке гена (в частности, на последовательности так называемого ТАТА-бокса), в состав которого входит РНК-полимераза, и активируют его (рис. 4.9).
Активация гена, таким образом, обеспечивается триадой: а) транскрипционные белки-активаторы, связывающиеся с энхансерами; б) транскрипционные факторы, взаимодействующие с ТАТА-боксом; в) РНК-полимераза II в комплексе со вспомогательными белками. Гены c-jun и c-fos относятся к числу генов немедленного реагирования (immediate-early genes), чья активность зависит от синтеза новых бел-
13*
195
ков и очень быстро возрастает вслед за клеточной стимуляцией.
Другой путь активации комплекса АР-1 — непосредственное фосфорилирование Fos (треонина в положении 232) и Jun (серины в положении 73 и 63). Эти события способствуют, с одной стороны, формированию активного комплекса Fos/Jun, а с другой — усилению его активирующего воздействия на базовый аппарат транскрипции. В фосфорилировании Jun участвует JNK (Jun N-terminal kinase), а в фосфорилировании Fos — FRK (Fos regulating kinase), входящие в семейство МАР-киназ.
В результате активации генов продуцируются белки, необходимые для синтеза ДНК и последующего митоза. Некоторые из новообразованных белков (Fos, Jun, Мус), известные как белки раннего ответа (immediate-early proteins), выполняют регуляторные функции; связываясь со специфическими участками ДНК, они активируют прилежащие гены. Другую группу составляют ферменты, такие как тимидинкиназа, рибонуклео-тидредуктаза, дигидрофолатредуктаза, тимидилатсинтаза, ор-нитиндекарбоксилаза, ДНК-полимеразы, топоизомеразы и множество других, которые имеют непосредственное отношение к синтезу ДНК. Усиливается, кроме того, общий белковый синтез, поскольку при каждом цикле удвоения воспроизводятся все клеточные структуры.
Нормальное прохождение клеткой отдельных фаз клеточного цикла обеспечивается строгой последовательностью и упорядоченностью во времени многочисленных ферментативных процессов. Роль «водителя ритма» при этом играют белки циклины, закономерным образом сменяющие друг друга на протяжении цикла.