

ЭКОЛОГИЯ_ПОСОБИЕ / 12_ГЛАВА 8
.rtfГЛАВА 8
Динамика и стабильность экосистем
Каждая экосистема – это динамическая структура, состоящая из сотен и тысяч видов продуцентов, консументов и редуцентов, которых связывают пищевые цепи и непищевые взаимоотношения. Экосистема поддерживает свое существование за счет круговорота биогенов и постоянного притока солнечной энергии.
Экосистема изменяется во времени вследствие изменения климатических и других экологических факторов. По мнению Ф.Клементса (1916), изменения сообществ во времени – естественный процесс и он носит название сукцессии.
Смены сообществ классифицируют на основе разных признаков: по направленности (циклические, направленные), по причинам смен (эндогенные, экзогенные (рис. 18)), по объектам, составляющим ядро сообщества (смена гетеротрофных или автотрофных сообществ).
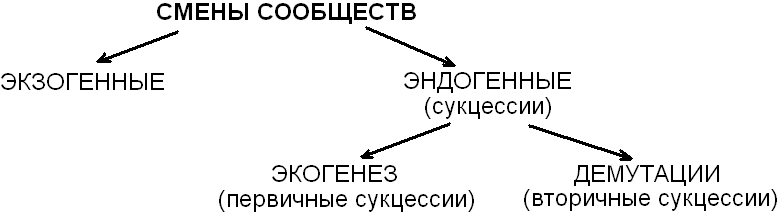
Рис. 18. Схема основных типов смен сообществ
В основе экзогенной смены сообществ лежит воздействие внешней среды.
Причины эндогенных смен – внутренние, определяются они особенностями и длительностью сосуществования видов в сообществе.
Сукцессии наиболее полно изучены на фитоценозах, так как в большинстве случаев именно фитоценоз определяет облик биоценоза. Это значит, что часто мы можем судить о динамике экосистемы по динамике фитоценоза. Сообщества в своей динамике проходят различные фазы (их называют стадиями или сериями), причём для каждой из них существует характерный набор видов, по которому можно сразу сказать, какую стадию сукцессии мы наблюдаем.
Сукцессия завершается формированием своего сообщества, которое адаптировано к климатическим условиям и природным особенностям экотопа. Такое сообщество Ф.Клементс назвал «климакс-формацией» или просто «климаксом».
Климаксные группировки не преобладают на большей части земной поверхности из-за постоянного влияния экзогенных факторов, которые «отбрасывают» сообщества на более ранние стадии. Значит, климакс стабилен лишь в стабильной среде. Климаксное сообщество представлено зональным типом растительности и почвы (в Средней полосе таким характерным сообществом будет сообщество на основе ассоциации дубрава + осока волосистая + лещина (зональные почвы – серые лесные)). Однако есть сериальные сообщества, которые способны существовать бесконечно долго, и длительность присутствия одной стадии не является признаком климакса. Этот факт связан с задержкой стадии развития сукцессии за счет действия внешних причин. В этом случае говорят о субклимаксе. Главным же отличием его от климакса является то, что устранение действия внешних факторов в дальнейшем непременно приведет к нормальному развитию сериального сообщества. Различают несколько типов субклимакса:
- рецидивный (при частом повторении одних и тех же нарушений – например, при частых пожарах)
- диаспорический (при отсутствии диаспор следующих стадий)
- ретардационный – замедление сукцессии под влиянием постоянно действующего фактора. Типичный пример – проточный водоём, в котором постоянно накапливающаяся органика периодически смывается, и в результате сериальная стадия задерживается на неопределенное время; так же происходит и вертикальный смыв органики под действием гравитации и воды – то есть, в горах сукцессия идет на порядок медленнее, чем на равнине.
Сукцессии проходят через фазу обнажения (появления незаселенного пространства), миграции (заселения его первыми поселенцами - пионерами); колонизации и приспособления к конкретным условиям обитания; соревнования (конкуренции с вытеснением ряда видов - пионеров) и реакции к стабилизации (обратного воздействия сообществ на биотоп и условия среды), формированию климаксного сообщества.
Структура сообщества создается постепенно в течение длительного времени. Так, при заселении обнаженной горной скалы деревья и кустарники не могут расти на ней, т. к. здесь нет необходимой для них почвы. Однако, водоросли и лишайники разными способами заселяют эти территории, образуя пионерные сообщества. Произрастая в мельчайших трещинах скал, они улавливают частички породы, которые приносит ветер или вода.
Постепенное накопление этих частичек, а также отмерших и разлагающихся организмов и эрозия горной породы (в результате выветривания и воздействия живого вещества) приводят к формированию слоя почвы, достаточного для поселения таких видов как мхи и папоротники. За этими видами последуют еще более требовательные к питательным веществам - травы. Они крошат своими корнями скалу и в конце концов слой почвы оказывается достаточным для развития деревьев и кустарников.
Такая смена одних видов другими за некоторый период времени называется «экологической сукцессией». Завершающее сообщество – устойчиво, самовозобновляется и находится в равновесии со средой; оно называется «климаксным сообществом». В животном мире тоже происходит смена одних видов другими, и она в значительной мере обусловлена сменой растительности. Этот процесс также зависит еще и от того, какие животные могут мигрировать из соседних сообществ.
Описанный выше тип сукцессии, начинающейся с заселения горной скалы или другой поверхности, лишенной почвы (песчаные дюны, бывшее ложе ледника), называется «первичной сукцессией».
Сукцессия, начинающаяся с твердого субстрата, называется ксеросерией; однако субстратом для возникновения сообщества может стать и вода. В этом случае мы говорим о гидросерии. И в гидросериях, и в ксеросериях сукцессии стремятся к мезофилизации условий: на исходно влажных местообитаниях накапливаются органические вещества, которые не успевают разлагаться из-за высокой влажности и недостатка О2. Это органическое вещество накапливается в виде торфа; постепенно его накопление поднимает поверхность над уровнем грунтовых вод. Со временем текущее сообщество попадает в менее влажные условия, чем те, в которых оно образовалось, и «чувствует себя хуже». Представители следующего сообщества более мезофильны, а значит, способны успешно конкурировать с представителями предыдущего.
В ксеросериях также происходит мезофилизация условий: при дефиците воды часть органики, образовавшейся в сообществе, не разлагается и накапливается в виде «сухой ветоши», которая гигроскопична и удерживает влагу. Значит, грунт становится более мезофильным.
Примером первичной сукцессии может служить и зарастание непроточного водоема (рис. 19) – например, пожарного пруда. Скорость и путь такого зарастания зависит от глубины водоема. При небольшой глубине зарастание происходит от берегов водоема, при большой – активное участие в освоении водной территории принимают сплавины.
Самой ранней стадией сукцессии наземных сообществ непроточных водоемов являются группировки, где доминирует сабельник. На следующей стадии доминирует Sphagnum balticum, после – Sphagnum fuscum. На этом сфагнуме поселяется клюква, багульник и некоторые другие виды. Со временем, по мере увеличения откладывания органики поверхность субстрата всё больше поднимается над зеркалом грунтовых вод. Происходит мезофилизация условий и замена болотных группировок на лесные сообщества, в которых доминируют сосна, зеленые мхи и брусника (такая ассоциация называется «сосняк-брусничник-зеленомошник»). Позже сосняк переходит в ельник-черничник, а тот – в лещиновую дубраву.

Рис. 19. Зарастание непроточного водоема
По-другому зарастают проточные водоемы, например, лесные ручьи. Сперва на самых влажных участках их берегов поселяются крупные осоки, камыш, тростник, на следующей стадии доминирует таволга вязолистная, после – гравилат речной, после – ольшаники, которые сменяются кленовыми или осиновыми лесами. Затем приходят ельники-майниковые, после – дубравы.
Из этого следует, что у гидросерии и ксеросерии цель одна – всё стремится к климаксному сообществу, характерному для данной местности.
Вторичная сукцессия начинается там, где поверхность полностью или в значительной степени лишена растительности, но прежде находилась под влиянием живых организмов и обладала органическим компонентом, т.е. это восстановление экосистемы, когда-то уже существовавшей на данной территории. Например, восстановление лесных вырубок или выгоревших участков. Из сохранившихся в почве семян, спор и других органов размножения вырастают мхи, лишайники и травы, которые быстро захватывают пространство. Затем они вытесняются злаками и разнотравьем. Постепенно в травяной покров внедряются молодые сосны, хорошо выдерживающие прямой солнечный свет и жару и затеняющие низкорослые, требующие хорошего освещения травы, и поле сменяется сосняком. Но под его пологом всходы сосны уже не могут развиваться, им тоже не хватает света. Вместо них формируется подрост широколиственных пород (дуб, клен), выдерживающий прохладные затененные условия. Их всходы нормально развиваются под родительским пологом, обеспечивая смену поколений. Сукцессия завершена.
При общности целей любых сукцессий пути достижения конечного устойчивого сообщества разные. Да, если взять две полные смены сообществ – от голого субстрата до климакса, общий вид серий будет схожим: сообщества низших растений сменятся сообществом трав, после появятся кустарники, после – деревья. Но сходство ограничится лишь единообразием жизненных форм в каждой отдельной стадии, с высокой долей вероятности мы сможем назвать и общие рода в списках сходства этих сообществ. Но вот присутствие тех или иных видов на стадии зависит от сложного переплетения микроусловий, больше случайного, чем закономерного. Можно привести такой пример: образование горных лесов – процесс сложный, а главное, долгий (с наклонного субстрата накопленную органику смывает первый же сильный дождь). Вырубая такие леса, человек возвращает сообщество на более раннюю стадию сукцессии – стадию луга. Произойди это на равнине, через два-три года территорию вырубки займут молодые деревца – осины, березы и т.п., а дальше сукцессия пойдет своим чередом. В горах же с открытого теперь с помощью рубки участка в кратчайшие сроки размоет всю накопленную за века органику до голого субстрата. Значит, на этом участке заново начнется экогенез, и леса там не будет еще много десятилетий. В силу этих причин для горного леса не рекомендуются сплошные рубки, то есть лес следует вырубать не полосами (как на равнинах), а делянками, и желательно, в шахматном порядке, что обеспечивает некоторую защиту от ветра и потоков воды.
Как при первичной, так и при вторичной сукцессиях флора и фауна окружающих территорий являются главным фактором, определяющим типы растений и животных, включающихся в сукцессию в результате случайного расселения и миграций.
На ранних стадиях сукцессии наиболее важны взаимодействия между растительными организмами и «абиотической» средой, которая, как правило, уже заселена микроорганизмами и некоторыми видами простейших. В результате деятельности организмов эта среда с течением времени изменяется. Хорошим примером может служить образование песчаных дюн вследствие накопления песка вокруг побегов песколюбов (Ammophila или Psamma), или формирование нивальных ландшафтов в высокогорьях Кавказа и других ледниковых пространств.
Время стабилизации дюн составляет несколько лет. Этот процесс на взморьях, в долинах рек и вокруг крупных озер приводит к появлению более крупных и не требовательных к питательным веществам растений (кустарники, тополя, сосны), поскольку там за длительный период времени образуется почва, в которой накапливается органический углерод, поступающий из подстилки. Увеличение численности азотфиксирующих организмов приводит к росту количества азота в почве. Здесь почвенные факторы играют определяющую роль в сукцессии и сами изменяются в процессе сукцессионных смен. Однако может потребоваться тысяча и более лет, чтобы химический состав почвы стабилизировался и сукцессию можно было считать законченной.
Этот пример иллюстрирует важный общий экологический принцип: ранние стадии сукцессии проходят значительно быстрее, чем поздние. Дальнейшее изменение климаксного сообщества происходит только при нарушении сложившихся условий окружающей среды, биосферы.
Переход из одной стадии сукцессии в другую стадию резких границ, конечно, не имеет – как и везде в природе, условия жизни не сменяются резко, а, напротив, приспособившиеся к ним сообщества «перетекают» друг в друга. Например, в сфагновом болоте есть кочки с той же брусникой и зелеными мхами. Или на границе ельника-кисличника с сенокосным лугом можно среди кислицы Oxalis acetosella и разных осок (род Carex) – типичных трав затененных местообитаний – встретить светолюбивые луговые травы, те же колокольчики (виды рода Campanula) или ромашки (виды Matricaria).
Лучше всего это видно на опушках лесов, по краям болот – вообще в зонах, где два абсолютно разных сообщества вливаются друг в друга, «борются» друг с другом за право владеть территорией, «засылая десант» – виды из своей группировки. Особенно это заметно в местах, где условия среды сочетаются удачно для обоих соседствующих сообществ. Как видим, зарастание этого озера идет ступенчато, в соответствии с теми изменениями, которые текущее сообщество вносит в окружающую среду. Так, «клюквенник-багульничник» сменяет сообщество Sphagnum magellanicum – просто потому, что этот последний слишком иссушил для себя среду!
Биотическое сообщество и биогеоценоз в целом могут локально изменять климат и формировать микроклимат. Аналогичным образом создаются микроместообитания и консорции, такие, как отмершие деревья, в которых может обитать около 200 (до 800) видов животных, навоз (более 300 видов животных, в основном жуки и мухи) и падаль (также заселяется жуками и мухами).
Своеобразие биосферных условий каждого региона Земли привело к развитию различных экосистем на земной поверхности. Режим температуры и осадков на некоторой территории в течение длительного времени и есть то, что мы называем “климатом”. Именно эти два экологических фактора, регулируемые потоком солнечного излучения, вместе с воздействием всей совокупности организмов определяют размещение по земной поверхности основных наземных биомов. Годовая сумма осадков ниже 1000 мм соответствует стрессовой зоне для многих видов деревьев, а предел устойчивости большинства из них составляет около 750 мм в год. В то же время у большинства злаков такой предел значительно ниже, примерно 250 мм/год, а кактусы и другие пустынные растения способны произрастать даже при 50-100 мм осадков в год. Соответственно, в местах с количеством осадков выше 750 мм/год обычно развиваются леса, от 250 до 750 мм/год - злаковые степи, а там, где осадков выпадает еще меньше, растительность представлена растениями пустынь. При промежуточных значениях годовой суммы осадков развиваются экосистемы переходного типа: лесостепи, полупустыни.
Температура также очень важна для определения характера экосистемы. Так, если развиваются лесные сообщества, то температура обуславливает, какой именно тип леса будет формироваться в регионе. Например, ельники и пихтарники характерны для холодных регионов, для севера и высокогорий. Листопадные деревья также в состоянии переносить морозную зиму, но требуют более долгого вегетационного периода, поэтому преобладают в умеренных широтах. Мощные вечнозеленые широколиственные породы с быстрым ростом не способны выдерживать даже кратковременных морозов и поэтому доминируют в субтропиках и тропиках. Более правильно отражают закономерности распределения экосистем (биогеоценозов) в биосфере Земли величины гидротермического индекса, представляющего собой отношение среднегодовых температур к осадкам, а не сами величины температур и осадков.
На более поздних стадиях сукцессии формируется детальная структура сообщества. Возрастает разнообразие живых организмов, их взаимодействия неизбежно становятся более сложными. Если сопоставить по количеству видов молодые сообщества Севера, находящиеся на ранней стадии развития (тундры) и зрелые климаксные сообщества тропиков, то очевидный перевес будет на стороне тропических климаксных лесов, где большинство видов фактически являются эдификаторами, и где каждый из них играет свою неповторимую роль – настолько уравновешена эта экосистема.
Из всего вышесказанного следует, что сукцессии есть закономерная и направленная смена одного сообщества другим, обусловленная внутренними причинами. Длительное существование сообщества вызывает накопление определенных веществ, то есть происходит изменение параметров среды, а значит – условия жизни представителей этого сообщества становятся непригодными. В результате новые виды вытесняют представителей этого сообщества, и происходит смена.
На всех стадиях сукцессии экосистема достаточно сбалансирована. Это динамическое равновесие (или «устойчивое неравновесие», по Вернадскому и Бауэру) обеспечивается за счет особых механизмов, обеспечивающих равновесие естественных популяций.
Популяционное равновесие может достигаться за счет взаимодействия «хищник – жертва», «паразит – хозяин», «фитофаг – растение» и др. Например в системе «фитофаг – растение» важную роль играют фитофаги, т. е. растительноядные животные, включая тысячи видов насекомых, нематод и др. беспозвоночных, а также крупных и мелких позвоночных. Обычно каждый из них специализируется на каком-то одном или нескольких видах растений. Когда плотность популяции какого-либо растения велика, специализированные на нем виды фитофагов получают преимущество и начинают активно размножаться. В этом случае их обилие растет быстрее, чем его успевают сокращать естественные враги и происходит вспышка численности фитофага. В результате этого популяция растения, служащего источником пищи для фитофагов, значительно уменьшается, и фитофаги начинают погибать из-за недостатка корма и усилившегося воздействия его врагов.
Вспышка численности фитофагов может повториться, если обилие растительной пищи вновь возрастет. Таким образом, для экосистем с низким разнообразием характерны резкие колебания размеров популяции слагающих их организмов. Однако низкая популяционная плотность какого-то растения дает возможность другим видам занять свободное место.
Растения будут мирно сосуществовать на одной территории до тех пор, пока одно из них не начнет вытеснять и увеличивать плотность популяции, что вызовет вспышку численности соответствующего фитофага.
В ходе естественного отбора виды все более приспосабливаются к сосуществованию с хищниками и паразитами, к климатическим условиям и другим экологическим факторам. При изменении любого фактора вид, плохо приспособленный к новым условиям, может мигрировать, вымереть или у него наступит адаптация к неблагоприятным факторам среды. Часть популяции может мигрировать, найти местообитания с подходящими условиями и продолжить там свое сосуществование.
Если в генофонде присутствуют гены, которые позволят некоторым особям выжить в новых условиях и оставить потомство, то через несколько поколений под действием естественного отбора возникнет популяция, хорошо приспособленная к изменившимся условиям существования. Когда же ни одна особь популяции не может мигрировать, спасаясь от воздействия неблагоприятных факторов, то популяция вымрет и ее генофонд исчезнет.
В том случае, когда виды вымирают, а выжившие особи других видов размножаются, адаптируются и изменяются под действием естественного отбора, говорят об эволюционной сукцессии.
Первое условие адаптации - выживание и размножение хотя бы нескольких особей в новых условиях. Это зависит от нескольких факторов:
- генетического разнообразия фонда;
- степени изменения среды;
- географического местоположения.
Если генофонд разнообразен (включает большой набор генов), то даже при сильных изменениях среды некоторые особи сумеют выжить и наоборот, малейшее колебание окружающих условий может привести к вымиранию вида, поскольку генов, позволяющих особям противостоять отрицательному воздействию среды, не найдется. Например, во время засухи из-за обедненного генофонда вначале гибнут культурные растения, а потом сорняки, т.к. их диапазон наследственности намного выше, чем сельскохозяйственных растений, и среди сорняков всегда найдутся виды, способные выжить в экстремальных условиях.
Когда изменение среды малозаметно и происходит постепенно, то большинство видов приспосабливается и выживает. Чем резче изменения, тем большее разнообразие генофонда требуется для выживания.
Еще один фактор, влияющий на жизнедеятельность вида, – это его географическое расположение. Чем шире распространен вид, тем выше его генетическое разнообразие и, наоборот. При обширном ареале некоторые его участки могут оказаться удаленными или изолированными от районов, где резко изменяются условия существования вида. В них вид сохранится, хотя исчезнет из других мест.
Если, благодаря наличию определенных генов, часть особей выживет в новых условиях, то восстановление популяции и дальнейшая ее адаптация будет зависеть от скорости воспроизводства вида, так как изменение признаков происходит только путем отбора в каждом поколении. Например, пара насекомых обычно дает несколько сотен потомков за непродолжительный период времени. Следовательно, скорость воспроизведения здесь в 1000 раз выше, чем у видов птиц, выкармливающих только 26 птенцов в год. И уровень приспособленности насекомых к новым условиям разовьется во столько же раз быстрее. Стоит ли удивляться, что насекомые быстро адаптируются и приобретают устойчивость к применяемым против них пестицидам, тогда как другие виды от этих обработок погибают. Таким образом, сокращение генетического разнообразия с одной стороны и ускоряющееся резкое изменение окружающей среды с другой, приводит к непредсказуемым экологическим последствиям.
Живые организмы, образующие популяции в различных экосистемах, в определенной степени способны противостоять внешним изменениям и восстановится до первоначального состояния, если эти внешние воздействия не слишком сильные, т. е. организмы имеют некоторую степень устойчивости. Существует 3 вида устойчивости живых систем:
Инертность – это способность живых систем сопротивляться различным нарушениям или изменениям.
Постоянство – это способность живых систем сохранять свои размеры.
Упругость – способность живых систем самовосстанавливаться после действия внешних нарушений, если они не были катастрофическими.
За счет чего же поддерживается стабильность экосистем? Как экосистема сохраняет свой состав, несмотря на столкновение интересов и конкуренцию многих слагающих ее видов и на изменения окружающей среды? Одно из условий, позволяющих экосистемам длительное время сохранять постоянный видовой состав состоит в том, что отношения между всеми их компонентами находятся в динамическом равновесии. Поддерживается равновесие между смертностью и рождаемостью видов с одной стороны, а с другой – каждый вид и популяция видов выполняет определенную функцию в экосистеме.
Роль разных видов в сообществе не одинакова. Обычно выделяют виды-доминанты и второстепенные виды – по численности, по плотности и биомассе. Например, по численности в ельнике-кисличнике доминируют насекомые и бактерии, а по биомассе – ель. На практике доминирующие виды определяют в разных систематических группах: в древесном ярусе – свои доминанты, среди птиц – свои; так же и среди мелких млекопитающих, насекомых и т.д.
Другая важная категория – виды-эдификаторы. Существует два основных толкования этого термина:
– виды-средообразователи, формирующие условия для других видов
– виды, формирующие внешний облик сообщества
Например, в ковыльной степи внешний облик сообщества определяет ковыль, а виды-средообразователи – копытные животные.
Эдификаторов сообщества можно определить по последствиям их уничтожения: при отсутствии такого вида сообщество сразу сменяется другим, рушится (остальные виды при этом мигрируют или погибают).
Кроме эдификаторов в сообществе можно выделить эксплеренты. Это виды, характерные для сообщества, с широкой экологической амплитудой. Они, как правило, замещают виды-эдификаторы при их частичном уничтожении (в случае слабых нарушений сообщества). Виды, чуждые сообществу, называются ценофобами. Они приспособлены к обитанию в разных условиях, внедряются в сообщество при нарушениях и живут в сообществе, пока последствия нарушений не устранятся.
Сохранение или рост численности популяции зависит от многих факторов: скорости размножения, способности животных мигрировать, а семян рассеиваться в подходящие местообитания на новых территориях; устойчивости к неблагоприятным условиям среды и болезням и т. д. Совокупность всех этих факторов, способствующих увеличению численности вида называют его биотическим потенциалом.
При благоприятных условиях среды численность индивидуумов увеличивается очень быстро. Например, в лабораторных условиях пара мышей дала потомство 2557 особей за один год. Но в естественных условиях такое наблюдается крайне редко, т. к. вероятность того, что все условия для жизни вида окажутся благоприятными, очень низка. Обычно один или несколько физических (неоптимальные температура, кислотность, влажность) и/или биотических (присутствие хищников, паразитов, болезнетворных организмов, нехватка пищи) факторов становятся лимитирующими. Сочетание всех таких ограничителей называют сопротивлением среды. Таким образом, рост, снижение или постоянство численности популяции зависит от соотношения между биотическим потенциалом (прибавлением особей) и сопротивлением среды (гибелью особей). При нарушении этого равновесия численность популяции изменяется.
Равновесие в популяции непрерывно регулируется, т. к. факторы среды редко подолгу остаются неизменными. Например, если в популяции резко снижается численность особей из-за засухи, то в последующие годы с нормальным увлажнением она полностью восстанавливается. Подобные циклические колебания обычно продолжаются неопределенно долго. Пока сократившаяся популяция еще способна восстановить прежнюю численность, равновесие существует. Если плотность ее растет, то сопротивление среды усиливается, в связи с чем увеличивается смертность и рост численности популяции прекращается или даже сменяется ее спадом. И напротив, с уменьшением плотности особей сопротивление среды обычно ослабевает, позволяя прежней численности восстановиться. Кроме того, биотический потенциал многих видов зависит от определенной минимальной, т. е. критической численности популяции. Если она падает ниже уровня, гарантирующего воспроизведение группы, то биотический потенциал резко снижается и вымирание становится практически неизбежным.