

Тимошкин и др 2014 эвтрофикация
.pdfОбщая гидробиология
ÓÄÊ 574.21
О. А. Тимошкин1, Н. А. Бондаренко1, Е. А. Волкова1, И. В. Томберг1, В. С. Вишняков1,2, В. В. Мальник1
МАССОВОЕ РАЗВИТИЕ ЗЕЛЕНЫХ НИТЧАТЫХ ВОДОРОСЛЕЙ РОДОВ SPIROGYRA И STIGEOCLONIUM (CHLOROPHYTA) В ПРИБРЕЖНОЙ ЗОНЕ ЮЖНОГО БАЙКАЛА*
В результате шестилетних (2008—2013 гг.) исследований макроводорослей прибрежной зоны западного берега южной котловины Байкала (исток р. Ангары
— зал. Лиственничный, междисциплинарный полигон у м. Березового, бух. Большие Коты от р. Черной до пади Варначки) обнаружено, что на глубине от 0,3 до 3,0 м во второй половине лета — осенью доминируют виды р. Spirogyra Link, ранее в озере не встречаемые. Изучение биологии водорослей в природной среде и в условиях культуры позволило идентифицировать один из доминирующих видов как S. fluviatilis Hilse. В зоне уреза бух. Большие Коты в ноябре 2012 г. впервые в масштабе всей бухты отмечено массовое развитие другой нитчатой водоросли — Stigeoclonium tenue (C. Agardh) K tz. Оценены масштабы продуцирования представителей р. Spirogyra в районе исследований. Показано, что в период максимального развития (ноябрь) водоросли могут создавать фитомассу до 317
± 143 ã/ì2 при 100%-ном проективном покрытии каменистого субстрата дна.
Ключевые слова: прибрежная зона, зеленые нитчатые водоросли, массовое развитие, Spirogyra, Stigeoclonium tenue, оз. Байкал.
Макроводоросли являются важным компонентом первичного трофиче- ского звена прибрежья озер и водохранилищ, они оказывают влияние на кислородный и биогенный режим, служат пищей и убежищем для беспозвоночных. Известно, что изменение трофического статуса водоемов (в том числе и в результате антропогенного воздействия) может приводить к массовому развитию несвойственных им макроводорослей, смене аборигенных видов-доминантов или к нарушению естественной для экосистемы поясности в их распределении [15, 23, 31, 34]. Великие Американские озера являются классическим примером массового развития зеленых нитчатых водорослей, связанного с увеличением биогенной нагрузки [16, 26—29, 38]. Береговые скопления детрита, состоящие в основном из отмерших и перегнивающих талломов водорослей, могут создавать серьезные проблемы. Известно,
* Работа выполнена в рамках госбюджетного проекта Сибирского отделения РАН ¹ VI.51.1.10 «Современное состояние, биоразнообразие и экология прибрежной зоны озера Байкал».
© О. А. Тимошкин, Н. А. Бондаренко, Е. А. Волкова, И. В. Томберг, В. С. Вишняков, В. В. Мальник, 2014
ISSN 0375-8990 Гидробиол. журн. 2014. ¹ 5. Т. 50 |
15 |

Общая гидробиология
что эти процессы негативно сказываются на рекреационных свойствах водоемов, вода становится непригодной для использования в пищевых целях. Гниющие водоросли являются одной из основных причин так называемого вторичного загрязнения водоемов органическими веществами и тяжелыми металлами, при этом могут происходить массовые заморы донных животных и рыб [24, 35, 36]. Кроме того, гниющие водоросли являются субстратом для размножения патогенных организмов кишечной микрофлоры [21, 25].
Большое значение макроводорослей для функционирования прибрежных сообществ оз. Байкал показано ранее [5, 6, 9]. Хорошо изучены их систематический состав, количественные параметры и особенности биологии [3, 5, 8, 11]. В годовой динамике развития донных макроводорослей озера выделяется летний максимум, когда кроме растущих круглый год видов, не имеющих ярко выраженных сезонных колебаний, в массе обнаруживаются водоросли с коротким периодом вегетации, дающие один пик биомассы. В это время четко выражены все пять известных для Байкала сменяющихся по вертикали растительных поясов [5, 8, 11]. Совсем недавно появились работы, свидетельствующие о нарушении зональности распределения макроводорослей озера. В частности, сообщается о замене в летний период 2011 г. в районе зал. Лиственничного эндемичных видов Draparnaldiodes, доминирующих в третьем растительном поясе, нитчатой водорослью Spirogyra sp. и о находке в 2012 г. большого количества спирогиры, прикрепленной к каменистому субстрату этого залива на глубине 0,3—0,5 м (в составе I растительного пояса) [3, 7]. Ранее в открытом Байкале нити спирогиры практически не встречались [5]. Также упоминается массовое развитие зеленой водоросли Stigeoclonium tenue (C. Agardh) Kütz. поздней осенью 2011 г. в урезовой зоне пади Жилище (бух. Большие Коты) [3]. Эта водоросль в значительном количестве была обнаружена на камнях уреза и небольшей глубине в зал. Лиственничном. Массовое развитие стигеоклониума в урезовой зоне Байкала при отсутствии обычного улотрикса до начала наших исследований не отмечалось. S. tenue встречается в Байкале вдоль северо-западного побережья южной котловины, в августе — сентябре — на каменистом субстрате на глубине 1,0—2,5 м, в этот же период он обильно вегетирует в р. Большой Котинке, впадающей в бух. Большие Коты [5].
Цель настоящей статьи — дать краткую характеристику новому для прибрежья Байкала явлению — смене видов доминантов (на примере бух. Большие Коты и зал. Лиственничного), привести сведения по экологии представителей родов Spirogyra Link. è Stigeoclonium Kütz., а также по идентификации доминирующих видов рода Spirogyra.
Материал и методика исследований**. В работе использованы материалы, полученные в 2008—2013 гг. как по натурным наблюдениям, так и в условиях культур. Для сравнения приводятся результаты анализа проб, собранных в 2003 г. в районе междисциплинарного полигона Березового на станциях трансект, расположенных над глубинами от 1,2 до 4,2 м. Пробы
** Помощь в отборе проб и организации экспедиций оказали сотрудники ЛИН СО РАН А. Г. Лухнев и Е. П. Зайцева. Авторы благодарны к. б. н. Л. С. Кравцовой за ценные советы и рекомендации.
16

Общая гидробиология
хранятся в фонде лаборатории биологии водных беспозвоночных Лимнологического института СО РАН (ЛИН СО РАН).
Многолетние полевые исследования проводили у западного берега южной котловины озера, в бух. Большие Коты, от р. Черной до пади Варначки (4—5 км), на базе стационара ЛИН СО РАН, а также в истоке р. Ангары. Распределение макрофитов изучали на трансектах, расположенных перпендикулярно линии берега. Стандартные трансекты, как правило, были длиной не менее 50—70 м и достигали свала глубин. Особенности вегетации макроводорослей обоих родов в пределах первых двух поясов растительности устанавливали на укороченных трансектах (10—20 м) до глубины 1,7—2,0 м. Отбор проб проводили при помощи аквалангистов либо с помощью камнещупов нескольких конструкций по заранее заданной схеме. Она включала: 1) видеопрофилирование трансекты, 2) отбор проб в пределах каждого пояса макрофитов, 3) макрофотографирование ландшафтов и участков дна в месте отбора проб с помощью учетной рамки площадью 0,25 м2 или масштабной линейки (для определения площади проективного покрытия) и 4) отбор количественных проб методом «stone-unit» [17] .
Для определения площади проективного покрытия не менее трех камней с обрастанием, взятых с каждой глубины и места, фотографировали, используя масштабную линейку. Площадь определяли по макрофотографиям с использованием оригинальной программы Sponge area***. Применение этого метода на других группах гидробионтов, его описание и последовательность компьютерной обработки фотографий приведены ранее [33].
Биомассу измеряли на весах Iuchi Sefi (IB-200H) (Япония) и Adventurer Ohaus AR 2140 (Китай). Обрастание с камней счищали скальпелем, тщательно промывали в воде и отделяли от примесей. Перед взвешиванием излишек воды удаляли фильтровальной бумагой. Для определения сухой массы водоросли высушивали в сушильном шкафу при температуре 105oС в течение трех часов.
Для расчета частоты встречаемости водорослей готовили временные прижизненные препараты с двух-трех точек поверхности каждого камня («стоун-юнита»). Конкретному месту и дате отбора соответствовало 25—30 микрофотографий, выполненных при увеличениях, достаточных для определения видовой или родовой принадлежности макроводорослей (от 200 до400). Микрофотографию условно приравнивали к отдельной пробе, в ней отмечали присутствие или отсутствие интересующих водорослей. Для некоторых проб (микрофотографий) просчитывали количество попавших в кадр нитей спирогиры или стигеоклониума, находили их относительное содержание (% общего количества макроводорослей в кадре). Микрофотографии получены с использованием световых микроскопов Olympus CX 21 и Meiji Techno при увеличении от 40 до 400, фотоаппаратов Olympus C-3040 с фотонасадкой NY 2000S 01705 и SONY Cyber-shot. Всего проанализировано 135 проб и более 2000 микрофотографий.
*** Разработчик программы К. П. Букшук.
17

Общая гидробиология
Культивировали водоросли в чашках Петри при естественном освещении и комнатной температуре на подоконнике лабораторной комнаты с северной стороны здания. Сначала их помещали в чашки с озерной водой, затем постепенно добавляли питательную среду Z-8 [30], имеющую активную реакцию (рН) соответствующую таковой исходной воды. Такие смешанные культуры в дальнейшем служили для изучения жизненного цикла водоросли и ее идентификации.
Для гидрохимического анализа воды отбирали два вида проб: поверхностные и придонные, последние — при помощи аквалангистов либо вручную, двумя шприцами, объемом 300 мл каждый, закрепленными на шесте длиной 1,5—2 м. Анализ выполнен общепринятыми в химии пресных вод методами [10, 12]. Предварительно пробы фильтровали от взвеси через мембранные (поликарбонатные) фильтры с диаметром пор 0,45 мкм.
Результаты исследований и их обсуждение
Â2009 г. при исследовании донных альгоценозов в бух. Большие Коты и
ååпритоках в незначительном количестве были найдены водоросли р. Spirogyra. В дальнейшем они были идентифицированны как S. varians (Hass.)
Kütz. (ðèñ. 1, ã, ä, å), которая развивается в реках Большой Котинке и Черной, и S. porticalis (O. Müll.) Cleve, обитающая в районе устья р. Малой Котинки. В р. Жилище также регистрировались стерильные нити спирогиры, морфологически близкие к S. varians из рек Черной и Большой Котинки.
В летне-осенний период 2011—2012 гг. по всему мелководью бух. Большие Коты, где расположен одноименный поселок, уже наблюдали массовую вегетацию чужеродных для озера водорослей р. Spirogyra. Они начинали интенсивно развиваться летом при температуре воды выше 10°С, достигая максимальной биомассы в ноябре при температуре воды около 4°С. В зимний период (t ~ 0°С) в прибрежье также отмечались незначительные скопления спирогиры. Аналогичная картина наблюдалась и в мелководной зоне зал. Лиственничного в период открытой воды 2012 г. Биомасса этих водорослей в осенний период сопоставима с максимальными значениями (210 г/м2) обычного обитателя урезовой зоны озера U. zonata в его летний пик развития (табл. 1).
При изучении жизненного цикла водорослей в условиях культуры было обнаружено, что по всему побережью доминировал ранее не зарегистрированный в озере вид Spirogyra fluviatilis Hilse [37] (ñì. ðèñ. 1, æ, ç, è). Он также не был встречен ни в одном из притоков бух. Большие Коты.
Как известно, S. fluviatilis — широко распространенная водоросль, обитающая в теплый период года в озерах, реках и ручьях [18, 32]. Часто массовое развитие представителей р. Spirogyra ухудшает качество воды [1, 14 и др.]. Например, увеличение обилия нитчатых водорослей родов Spirogyra è Zygnema Ag. в литорали оз. Конесус (Conesus Lake, США) привело к изменению экологического статуса этой зоны и может оказать каскадный эффект на экосистему озера в целом [14]. Авторы связывают это явление с контро-
18
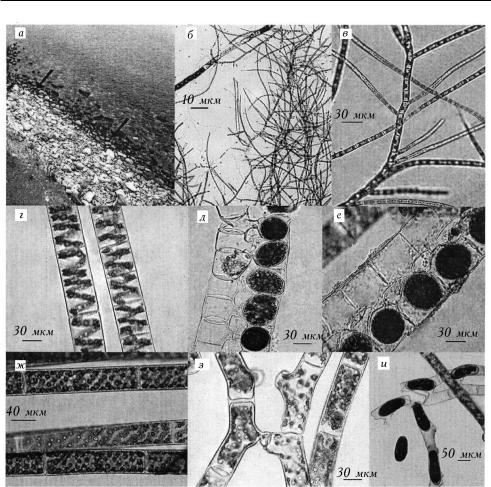
Общая гидробиология
1. Фото прибрежья озера и микрофотографии водорослей: à — ïîÿñ Stigeoclonium tenue (стрелки) в прибрежье оз. Байкал, бух. Большие Коты; á, â — S. tenue, 100, 400; ã, ä — Spirogyra varians из р. Большой Котинки: вегетативные нити (ã), коньюгация с зигоспорами (ä), 400; å — S. varians из р. Черной: коньюгация с зигоспорами, 400; æ, ç, è — S. fluviatilis из бух. Большие Коты: вегетативные нити (æ), коньюгирующие нити (ç), 400, коньюгация с зигоспорами (è), 100.
лируемой гидрометеорологическими событиями водностью притоков, привносящих значительное количество растворимых форм фосфора и азота.
Показано, что S. fluviatilis интенсивно развивается летом в водах, богатых азотом и фосфором. При этом важным фактором для успешного потребления питательных элементов водорослями и интенсивности их фотосинтеза является умеренная скорость течения (от 12 до 15 см/сек) [18, 19].
Стерильные нити спирогиры в оз. Байкал отмечены многими авторами [5, 6, 8 и др.], однако ее массовое развитие ранее никогда не регистрировалось. Недостаток регулярных исследований донных макроводорослей за последние 15 лет не позволяет точно установить начало обсуждаемого явления. В связи с этим был проведен анализ проб, отобранных в 2003 г. Обнару-
19

Общая гидробиология
1. Площадь проективного покрытия и масса S. fluviatilis (осень 2012 г.)
Äàòà |
Места отбора |
Проективное |
Сырая масса, |
Сухая масса, |
|
покрытие, % |
ã/ì2 |
ã/ì2 |
|||
|
|
||||
18 сентября |
Бух. Большие Коты, |
53 ± 13,5 |
190 ± 70 |
33,90 ± |
|
|
напротив биостанции, |
|
|
12,65 |
|
|
глубина 1,0—1,5 м |
|
|
|
|
9 ноября |
Бух. Большие Коты, |
72 ± 27,5 |
176 ± 91 |
31,45 ± |
|
|
напротив стационара |
|
|
18,44 |
|
|
ЛИН СО РАН, глуби- |
|
|
|
|
|
íà 1,0—1,5 ì |
|
|
|
|
26 ноября |
Зал. Лиственничный, |
89 ± 23,6 |
317 ± 143 |
56,62 ± |
|
|
напротив Байкальско- |
|
|
22,67 |
|
|
го музея, глубина |
|
|
|
|
|
1,0—1,5 ì |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
жено, что в составе макроводорослей встечались лишь единичные стерильные нити р. Spirogyra. В истоке р. Ангары (возле пирса Байкальского музея в пос. Листвянке) постоянное присутствие небольшого количества этих водорослей мы регистрировали с 2008 г. Все это свидетельствует о том, что развитие представителей р. Spirogyra в литорали оз. Байкал приобрело массовый характер лишь за последние несколько лет.
Еще одна нехарактерная для макрофитов Байкала особенность распределения была обнаружена в ноябре 2011 г. в пади Жилище (бух. Большие Коты). Большинство камней, расположенных в зоне уреза, были на 100% покрыты ярко-зеленым обрастанием [3]. Внешне эта зона ничем не отличалась от типичного для открытого Байкала 1-го пояса, состоящего из U. zonata (ñì. ðèñ. 1, à). Микроскопический анализ показал, что обрастание представляет собой нити зеленой водоросли, определенной как Stigeoclonium tenue (ñì. ðèñ. 1, á, â).
В ноябре 2012 г. аналогичные работы проводились в Южном Байкале, в том числе бух. Большие Коты в районе р. Черной до падей Варначки и Сенной. Микроскопическое исследование камней, расположенных в зоне уреза показало, что обрастание на 90—100% состояло из нитчатых водорослей р. Stigeoclonium и на 10% и менее — из Ulothrix. Средняя сырая биомасса первого составляла 322 ± 58 г/м2. Пробы, взятые в урезовой зоне пади Жилище бух. Большие Коты в 2013 г., также содержали значительное количество нитей стигеоклониума.
Таким образом, можно сделать вывод о смене доминирующего вида первого растительного пояса открытого Байкала, происходящей поздней осенью, как минимум в пределах целой бухты в период 2011—2013 гг. Массовые находки стигеоклониума в пробах из урезовой зоны зал. Лиственничного, а также его значительное количество в летне-осенних сборах 2013 г. в урезовой зоне западного побережья Малого Моря и северной оконечности Байкала (участок между городами Северобайкальском и Нижнеангарском) значительно расширяют масштабы этого нового для Байкала явления.
20
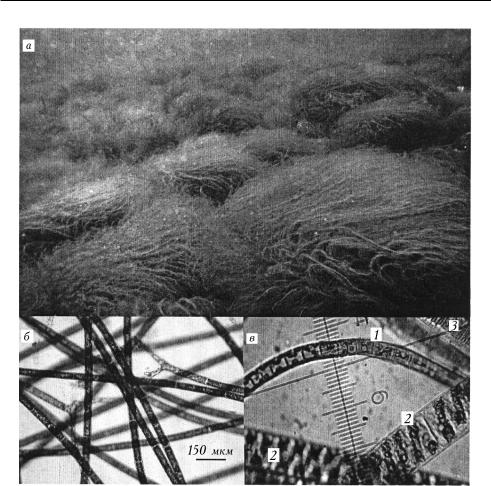
Общая гидробиология
2. Подводная фотосъемка и микрофотографии донных водорослей: à — массовое развитие спирогиры на каменистом субстрате в бух. Большие Коты, ноябрь 2012 г., глубина 1,0—1,5 м; áâ — примеры микрофотографий с разным относительным обилием водорослей на препаратах: á — нитчатые водоросли рода Spirogyrac — смешанные обрастания: 1 — Ulothrix zonata; 2 — Spirogyra sp.; 3 — Fragilaria sp.
Что же вызвало сложившуюся ситуацию в Байкале? Один из вариантов ответа в отношении стигеоклониума может быть весьма прост: маловероятно, но возможно, что предыдущими исследованиями не охватывался позд- не-осенний период. Также не исключено, что эти водоросли просто принимались за улотрикс — ведь без микроскопического исследования оба типа обрастания практически неразличимы.
Возможные причины массового развития несвойственных для озера видов легче всего свести к антропогенному фактору. Авторы [7], наблюдавшие такое явление в районе пос. Листвянка (зал. Лиственничный), связали его с возрастанием биогенной нагрузки на прибрежную зону в результате увели- чения потока туристов. Но, как следует из гидрохимических исследований, ни в районе м. Березового, ни в бух. Большие Коты не отмечено значительных изменений в биогенной нагрузке по сравнению с таковой в 1950-е и 1960-е годы, служащие фоновыми для дальнейших исследований [4, 13].
21

Общая гидробиология
2. Концентрация химических компонентов в прибрежной воде (1 м от берега)
зал. Лиственничного и бух. Большие Коты
|
|
Бух. Большие Коты |
Зал. Лиственничный |
||||
Äàòà |
|
|
|
|
|
|
|
Pìèí, |
Сумма Nìèí, |
ÏÎ, ìã |
Pìèí, |
Сумма Nìèí, |
ÏÎ, ìã |
||
|
|||||||
|
ìêã/äì3 |
ìã/äì3 |
Î/äì3 |
ìêã/äì3 |
ìã/äì3 |
Î/äì3 |
|
|
|
|
2011 ã. |
|
|
|
|
èþíü |
0—4 |
0,01—0,01 |
0,93—0,94 |
|
|
|
|
август |
0—4 |
0,03—0,08 |
0,63—1,1 |
|
|
|
|
сентябрь |
1—7 |
0,01—0,05 |
1,01 |
10 |
0,10—0,11 |
1,24—1,86 |
|
ноябрь |
4—8 |
0,04 |
0,92 |
8 |
0,12 |
0,92 |
|
|
|
|
2012 ã. |
|
|
|
|
èþíü |
3—7 |
0,03—0,07 |
0,92—1,77 |
|
|
|
|
èþëü |
1—7 |
0,04—0,1 |
0,69—1,58 |
5 |
0,13 |
0,85 |
|
сентябрь |
0—7 |
0,02—0,04 |
0,69—1,27 |
2—5 |
0,17—0,32 |
1,7—1,9 |
|
7 ноября |
6—11 |
0,03—0,1 |
0,69—1,27 |
10 |
0,14—0,16 |
0,88—1,00 |
|
26 ноября* |
|
|
|
8—10 0,09—0,18 0,69-2,00 |
|||
|
|
|
|
4—10 |
0,08—0,62 |
1,33—4,61 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
П р и м е ч а н и е. ПО — перманганатная окисляемость; — исследования не проводили; * отбор придонных проб воды шприцами вблизи камней с обрастаниями.
Было показано, что содержание фосфатного фосфора в прибрежных водах озера и в 100 м от уреза было невысоким — 1—6 мкг/дм3 [13] и только в воде зал. Лиственничного оно возрастало до 10,5 мкг P/дм3
Наше исследование химии воды показало, что концентрация минерального фосфора в прибрежной воде в бух. Большие Коты в летние месяцы снижалась до нулевых значений, а осенью достигала максимальных — 11 мкг P/дм3 (òàáë. 2).
В прибрежной воде зал. Лиственничного концентрация фосфатов летом не опускалась ниже 2 мкг P/дм3, а осенью не превышала 10 мкг P/дм3, содержание минерального азота было в 2—8 раза выше, чем в бухте. В первом случае 70—95% суммарного количества составлял нитратный азот, во втором эта форма преобладала только в летние месяцы. В начале июня и сентябре — ноябре в прибрежной воде бух. Большие Коты доля аммонийного увеличивалась до 60% общего содержания минерального азота. Содержание органических веществ в прибрежной воде заливов в летне-осенний период не превышало 2 мг О/дм3 (по ПО). Однако осенью, при отмирании и разложении макроводорослей, в придонной воде (пробы, отобранные шприцами) этот показатель возрастал до 4,6 мг О/дм3, а содержание минерального азота — до 0,62 мг N/дм3.
22

Общая гидробиология
У западного берега от бух. Песчаной до зал. Лиственничного существуют области с устойчивыми течениями. Подо льдом скорость достигает 9—10 см/сек, в период открытой воды она на порядок выше [2], то есть превышает оптимальные (от 12 до 15 см/сек) для успешного роста водорослей. Тем не менее в Южном Байкале отмечена интенсивная вегетация S. fluviatilis. В заливах влияние циклонических течений выражено слабее, спирогиры развиваются в прибрежной полосе шириной 20—30 м.
На современном этапе оценить причины ситуации, сложившейся в литорали озера Байкал, и дать прогноз непросто. Можно предположить две основные причины массового развития чужеродных для озера нитчатых водорослей. Во-первых, это может быть следствием естественной цикличности в их развитии под воздействием глобальных факторов, например климатических. Во-вторых, массовое развитие спирогиры зачастую приурочено к местам повышенной концентрации биогенных элементов, обусловленной, в частности, сбросом недостаточно очищенных сточных вод [20, 22]. Населенные пункты, располагающиеся в прибрежной зоне оз. Байкал, как правило, не имеют централизованной системы очистки бытовых сточных вод. Коли- чество частных гостиниц и туристов за последние годы выросло в несколько раз. Грунты надводной и заплесковой зон являются своеобразным буфером, фильтром, защищающим чистоту озер, однако их фильтрационная способность явно ограничена. В зал. Лиственничном предел этой способности, по-видимому, уже превышен. По нашим данным, в поровых водах зоны заплеска, а также в пробах придонной воды на мелководье бух. Большие Коты и зал. Лиственничного в летний период 2010—2013 гг. регулярно обнаруживались следы фекального загрязнения, обусловленного прежде всего пассивной фильтрацией сточных вод прибрежных поселков. Например, в ноябре 2012 г. количество термотолерантных колиформных бактерий (ТКБ) и энтерококков в пос. Листвянка, в лунке, выкопанной на пляже, было очень высоким (соответственно 15 000 и 3700 КОЕ/100 мл). Значительное количество общих колиформных бактерий, ТКБ и энтерококков в это же время было обнаружено и в пробах придонной воды, взятых шприцами: соответственно 210, 80 и 66 КОЕ/100 мл.
Заключение
Многолетние (2003, 2008—2013 гг.) исследования структуры и количественных характеристик макроводорослей прибрежной зоны западного берега южной котловины Байкала показали, что на глубинах от 0,3 до 3,0 м (и более) во второй половине лета — осенью доминируют не отмечаемые ранее в озере виды р. Spirogyra. Изучение биологии водорослей в природной среде и в условиях культуры позволило идентифицировать один из доминирующих видов как S. fluviatilis Hilse. Этот вид приводится для флоры озера впервые и пока в притоках исследуемого района не обнаружен. В период максимального развития (ноябрь) биомасса водорослей может достигать 317 ± 143 г/м2 при 100%-ном проективном покрытии каменистого субстрата дна. В зоне уреза бух. Большие Коты и зал. Лиственничного отмечено массовое развитие и другой нитчатой водоросли — Stigeoclonium tenue (C. Agardh) Kutz., которая ранее встречалась в озере на глубине 1—2,5 м, а в заметном количестве развивалась только в р. Большой Котинке. Осенью при отмирании и разложении макроводорослей в придонной воде проис-
23

Общая гидробиология
ходит значительное увеличение содержания органических веществ (в 2,0—2,5 раза). Полученные результаты, наряду с литературными сведениями, свидетельствуют о начале евтрофикации участков прибрежной зоны Байкала, приуроченных к ряду населенных пунктов.
**
У статт³ наведено матер³али щодо масового розвитку у прибережн³й смуз³ п³вденно¿ частини оз. Байкал нитчастих водоростей р. Spirogyra, як³ ран³ше в озер³ не зустр³чались, та Stigeoclonium tenue у друг³й половин³ л³та — восени. Обговорюються причини цього явища.
**
Paper deals with unusual mass vegetation of filamentous microalga of the gen. Spirogyra and Stigeoclonium tenue in late summer — autumn in the coastal zone of the southern section of Lake Baikal over the years 2008—2013. Probable reasons of this phenomenon are discussed.
**
1.Вассер С.П., Кондратьева Н.В., Масюк Н.П. и др. Водоросли. Справоч- ник. — Киев: Наук. думка, 1989. — 605 с.
2.Верболов В.И. Течения и водообмен в Байкале // Вод. ресурсы. — 1996.
—Ò. 23, ¹ 4. — Ñ. 413—423.
3.Вишняков В.С., Тимошкин О.А., Ижболдина Л.А., Волкова Е.А. Таксономи- ческий список макроводорослей прибрежной зоны бухты Большие Коты и залива Лиственничный (Южный Байкал) // Изв. Иркут. óí-òà. Сер. Биология. Экология. — 2012. — Т. 5, ¹ 3. — С. 147—159.
4.Голобокова Л.П., Сакирко М.В., Онищук Н.А. и др. Гидрохимическая характеристика вод литорали северо-западного участка Южного Байкала // Аннотированный список фауны озера Байкал и его водосборного бассейна. — Новосибирск: Наука, 2009. — Т. 2: Водоемы и водотоки юга Восточной Сибири и Северной Монголии. Справочники и определители по фауне и флоре озера Байкал. — С. 760—784.
5.Ижболдина Л.А. Атлас и определитель водорослей бентоса и перифитона озера Байкал (мейо- и макрофиты) с краткими очерками по их экологии.
—Новосибирск: Наука-центр, 2007. — 248 с.
6.Кожов М.М. О видообразовании в озере Байкал // Бюл. МОИП. Отд. биол. — 1960. — Т. 65, ¹ 6. — С. 39—47.
7.Кравцова Л.С., Ижболдина Л.А., Ханаев И.В. и др. Нарушение вертикальной зональности зеленых водорослей в прибрежной части залива Лиственничный озера Байкал // Докл. РАН. — 2012. — Т. 447, ¹ 2. — С. 227—229.
8.Мейер К.И. Введение во флору водорослей оз. Байкал // Бюл. МОИП. — 1930. — Т. 39, ¹ 3—4. — С. 179—392.
9.Окунева Г.Л. Сезонные изменения мезобентоса на каменистой литорали (район пос. Большие Коты) // Продуктивность Байкала и антропогенные изменения его природы. — Иркутск: Изд-во Иркут. ун-та, 1974. — С. 137—152.
24