

СТРОЕНИЕ И ФУНКЦИИ КЛЕТКИ
.pdfСТРОЕНИЕ И ФУНКЦИИ КЛЕТКИ
Кле́тка— элементарная единица строения и жизнедеятельности всех организмов (кроме вирусов, о которых нередко говорят как о неклеточных формах жизни), обладающая собственным обменом веществ, способная к самостоятельному существованию, самовоспроизведению и развитию. Все живые организмы либо состоят из множества клеток (многоклеточные животные, растения и грибы), либо являются одноклеточными организмами (многие простейшие и бактерии). Раздел биологии, занимающийся изучением строения и жизнедеятельности клеток, получил название цитологии. В последнее время принято также говорить о биологии клетки, или клеточной биологии.
Обычно размеры растительных и животных клеток колеблются в пределах от 5 до 20 мкм в поперечнике. Типичная бактериальная клетка значительно меньше – ок. 2 мкм, а наименьшая из известных – 0,2 мкм.
Некоторые свободноживущие клетки, например такие простейшие, как фораминиферы, могут достигать нескольких сантиметров; они всегда имеют много ядер. Клетки тонких растительных волокон достигают в длину одного метра, а отростки нервных клеток достигают у крупных животных нескольких метров. При такой длине объем этих клеток небольшой, а поверхность очень велика.
Самые крупные клетки – это неоплодотворенные яйца птиц, заполненные желтком. Наибольшее яйцо (и, следовательно, наибольшая клетка) принадлежало вымершей громадной птице – эпиорнису (Aepyornis). Предположительно его желток весил ок. 3,5 кг. Самое крупное яйцо у ныне живущих видов принадлежит страусу, его желток весит ок. 0,5 кг
Одно время клетка рассматривалась как более или менее гомогенная капелька органического вещества, которую называли протоплазмой или живой субстанцией. Этот термин устарел после того, как выяснилось, что клетка состоит из множества четко обособленных структур, получивших название клеточных органелл («маленьких органов»).
Первым человеком, увидевшим клетки, был английский учѐный Роберт Гук (известный нам благодаря закону Гука). В 1665 году, пытаясь понять, почему пробковое дерево так хорошо плавает, Гук стал рассматривать тонкие срезы пробки с помощью усовершенствованного им микроскопа. Он обнаружил, что пробка разделена на множество крошечных ячеек, напомнивших ему соты в ульях медоносных пчел, и он назвал эти ячейки клетками (по-английски cell означает «ячейка, клетка»).

В 1675 году итальянский врач М. Мальпиги, а в 1682 году — английский ботаник Н. Грю подтвердили клеточное строение растений. О клетке стали говорить как о «пузырьке, наполненном питательным соком». В 1674 году голландский мастер Антоний ван Левенгук (Anton van Leeuwenhoek, 1632—1723) с помощью микроскопа впервые увидел в капле воды «зверьков» — движущиеся живые организмы (инфузории, амѐбы, бактерии). Также Левенгук впервые наблюдал животные клетки — эритроциты и сперматозоиды. Таким образом, уже к началу XVIII века учѐные знали, что под большим увеличением растения имеют ячеистое строение, и видели некоторые организмы, которые позже получили название одноклеточных. В 1802—1808 годах французский исследователь Шарль-Франсуа Мирбель установил, что все растения состоят из тканей, образованных клетками. Ж. Б. Ламарк в 1809 году
распространил идею Мирбеля о клеточном строении и на животные организмы. В 1825 году чешский учѐный Я. Пуркине открыл ядро яйцеклетки птиц, а в 1839 ввѐл термин «протоплазма». В 1831 году английский ботаник Р. Броун впервые описал ядро растительной клетки, а в 1833 году установил, что ядро является обязательным органоидом клетки растения. С тех пор главным в организации клеток считается не мембрана, а содержимое.
Методы исследования клеток
Впервые клетки удалось увидеть только после создания световых микроскопов, с того времени и до сих пор микроскопия остается одним из важнейших методов исследования клеток. Световая (оптическая) микроскопия, несмотря на своѐ сравнительно небольшое разрешение, позволяла наблюдать за живыми клетками. В ХХ веке была изобретена электронная микроскопия, давшая возможность изучить ультраструктуру клеток.
В изучении клеточной формы и структуры первым инструментом был световой микроскоп. Его разрешающая способность ограничена размерами, сравнимыми с длиной световой волны (0,4–0,7 мкм для видимого света). Однако многие элементы клеточной структуры значительно меньше по размерам.
Другая трудность состоит в том, что большинство клеточных компонентов прозрачны и коэффициент преломления у них почти такой же, как у воды. Для улучшения видимости часто используют красители, имеющие разное сродство к различным клеточным компонентам. Окрашивание применяют также для изучения химии клетки. Например, некоторые красители связываются преимущественно с нуклеиновыми кислотами и тем самым выявляют их локализацию в клетке. Небольшая часть красителей
– их называют прижизненными – может быть использована для окраски живых клеток, но обычно клетки должны быть предварительно зафиксированы (с помощью веществ, коагулирующих белок) и только после этого могут быть окрашены.
Перед проведением исследования клетки или кусочки ткани обычно заливают в парафин или пластик и затем режут на очень тонкие срезы с помощью микротома. Такой метод широко используется в клинических лабораториях для выявления опухолевых клеток. Помимо обычной световой микроскопии разработаны и другие оптические методы изучения клетки: флуоресцентная микроскопия, фазово-контрастная микроскопия, спектроскопия и рентгеноструктурный анализ.

Оптическая микроскопия
В оптическом микроскопе увеличение объекта достигается благодаря серии линз, через которые проходит свет. Максимальное увеличение, которое можно достичь благодаря оптическому микроскопу, составляет около 1000. Еще одной важной характеристикой является
разрешение — расстояние между |
двумя |
|
точками, которые еще распознаются |
||
отдельно, другими словами, разрешение |
||
характеризует чѐткость изображения. Эта |
||
величина ограничивается длиной световой |
||
волны, и даже при использовании самого |
||
коротковолнового |
света — |
|
ультрафиолетового — |
можно |
достичь |
разрешения только около 200 нм; такое разрешение было получено еще в конце
XIX века. Таким образом, малейшие структуры, которые можно наблюдать под оптическим микроскопом, это митохондрии и бактерии, линейный размер которых составляет примерно 500 нм. Однако объекты размером меньше 200 нм видны в световом микроскопе только тогда, если они сами излучают свет. Эта особенность используется в флуоресцентной микроскопии, когда клеточные структуры или отдельные белки связываются со специальными флуоресцентными белками или антителами с флуоресцентными метками. На качество изображения, полученного с помощью оптического микроскопа, влияет также контрастность — еѐ можно увеличить, используя различные методы окраски клеток. Для изучения живых клеток используют фазовоконтрастную, дифференциальную интерференционно-контрастную и темнопольную микроскопию. Конфокальные микроскопы позволяют улучшить качество флуоресцентных изображений.
Электронная микроскопия
В 30-х годах XX века был сконструирован электронный микроскоп, в котором вместо света через объект пропускается пучок электронов. Теоретический предел разрешения для современных электронных микроскопов составляет около 0,002 нм, однако из практических причин для биологических объектов достигается разрешение только около 2 нм. С помощью электронного микроскопа можно изучать ультраструктуру клеток. Различают два основных типа электронной микроскопии:
сканирующую и трансмиссионную.
Сканирующая (растровая) электронная микроскопия (РЭМ) используется для изучения поверхности объекта. Образцы зачастую покрывают тонкой пленкой золота. РЭМ
позволяет получать объемные изображения. Трансмиссионная (просвечивающая) электронная микроскопия (ПЭМ) — используется для изучения внутреннего
строения клетки. Пучок электронов пропускается через объект, предварительно обработанный тяжелыми металлами, которые накапливаются в определенных структурах, увеличивая их электронную плотность. Электроны рассеиваются на участках клетки с большей электронной плотностью, в результате чего на изображениях эти области выглядят темнее.
Фракционирование клеток. Для установления функций отдельных компонентов клетки важно выделить их в чистом виде, чаще всего это делается с помощью метода дифференциального центрифугирования. Разработаны методики, позволяющие получить чистые фракции любых клеточных органелл. Получение фракций начинается с разрушения плазмалеммы и образования гомогената клеток. Гомогенат последовательно центрифугируется при различных скоростях, на первом этапе можно получить четыре фракции: (1) ядер и крупных обломков клеток, (2) митохондрий, пластид, лизосом и пероксисом, (3) микросом — пузырьков аппарата Гольджи и эндоплазматического ретикулума, (4) рибосом, в супернатанте останутся белки и более мелкие молекулы. Дальнейшее дифференциальное центрифугирование каждой из смешанных фракций позволяет получить чистые препараты органелл, к которым можно применять разнообразные биохимические и микроскопические методы.
Строение клеток
Все клеточные формы жизни на Земле можно разделить на два надцарства на основании строения составляющих их клеток:
прокариоты (доядерные) — более простые по строению;
эукариоты (ядерные) — более сложные. Клетки, составляющие тело человека, являются эукариотическими.
Несмотря на многообразие форм, организация клеток всех живых организмов подчинена единым структурным принципам.
Прокариотическая клетка
Прокариоты (от лат. pro — перед, до и греч. κάρῠον — ядро, орех) — организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов — линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелѐные водоросли), и археи. Основное содержимое клетки, заполняющее весь еѐ объѐм, — вязкая зернистая
цитоплазма.
Эукариотическая клетка
Эукариоты (эвкариоты) (от греч. ευ — хорошо, полностью и κάρῠον — ядро, орех)
— организмы, обладающие, в отличие от прокариот, оформленным клеточным ядром, отграниченным от цитоплазмы ядерной оболочкой. Генетический материал заключѐн в нескольких линейных двухцепочных молекулах ДНК (в зависимости от вида организмов их число на ядро может колебаться от двух до нескольких сотен), прикреплѐнных изнутри к мембране клеточного ядра и образующих у подавляющего большинства комплекс с белками-гистонами, называемый хроматином.
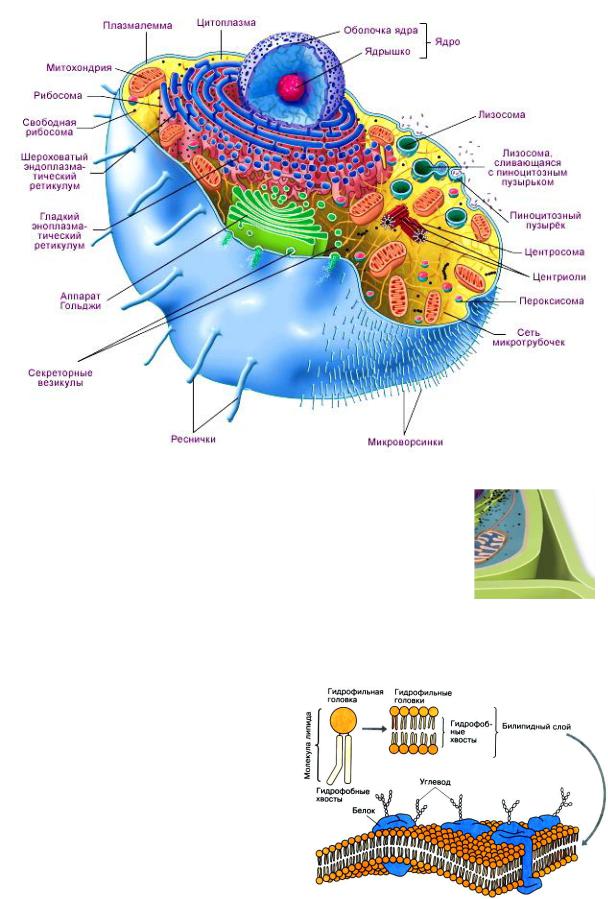
Строение эукариотической клетки. Схематическое изображение животной клетки.
Некоторые клетки, в основном растительные и бактериальные, имеют наружную клеточную стенку. У высших растений она состоит из целлюлозы. Клеточная стенка играет исключительно важную роль: она представляет собой внешний каркас, защитную оболочку, обеспечивает тургор растительных клеток: через клеточную стенку проходит вода, соли, молекулы многих органических веществ.. У клеток животных клеточные стенки, как правило, отсутствуют.
Под клеточной стенкой растений расположена плазматическая мембрана или плазмалемма. Толщина плазматической мембраны около 10 нм, изучение ее строения и функций возможно только с помощью электронного микроскопа.
Внутри клетка заполнена цитоплазмой, в которой расположены различные органоиды и клеточные включения, а также генетический материал в виде молекулы ДНК. Каждый из органоидов клетки выполняет свою особую функцию, а в совокупности все они определяют жизнедеятельность клетки в целом.
Плазматическая мембрана обеспечивает в первую очередь разграничительную функцию по отношению к внешней для
клетки среде. Она представляет собой двойной слой молекул (бимолекулярный слой, или бислой). В основном это молекулы фосфолипидов и других близких к ним веществ. Липидные молекулы имеют двойственную природу, проявляющуюся в том, как они ведут себя по отношению к воде. Головы молекул гидрофильные, т.е. обладают сродством к воде, а их углеводородные хвосты гидрофобны. Поэтому при смешивании с водой липиды образуют на ее поверхности пленку, аналогичную пленке масла; при этом все их молекулы ориентированы одинаково: головы молекул – в воде, а углеводородные хвосты – над ее поверхностью.
Вклеточной мембране два таких слоя, и в каждом из них головы молекул обращены наружу, а хвосты – внутрь мембраны, один к другому, не соприкасаясь таким образом с водой.
Кроме основных липидных компонентов, она содержит крупные белковые молекулы, которые способны «плавать» в липидном бислое и расположены так, что одна их сторона обращена внутрь клетки, а другая соприкасается с внешней средой. Некоторые белки находятся только на наружной или только на внутренней поверхности мембраны или лишь частично погружены в липидный бислой.
Основная функция клеточной мембраны заключается в регуляции переноса веществ в клетку и из клетки.
Существует несколько механизмов транспорта веществ через мембрану:
Диффузия — проникновение веществ через мембрану по градиенту концентрации (из области, где их концентрация выше, в область, где их концентрация ниже). Диффузный транспорт веществ осуществляется при участии белков мембраны, в которых имеются молекулярные поры (вода, ионы), либо при участии липидной фазы (для жирорастворимых веществ).
Облегченная диффузия — специальные мембранные белки-переносчики избирательно связываются с тем или иным ионом, или молекулой и переносят их через мембрану.
Активный транспорт. Этот механизм сопряжен с затратами энергии и служит для переноса веществ против их градиента концентрации. Он осуществляется специальными
белками-переносчиками, образующими так называемые ионные насосы. Наиболее изученным является Nа+/К+-насос в клетках животных, активно выкачивающий ионы Nа наружу, поглощая при этом ионы К+ .
Всочетании с активным транспортом ионов в клетку через цитоплазматическую мембрану проникают различные сахара, нуклеотиды, аминокислоты.
Такая избирательная проницаемость физиологически очень важна, и ее отсутствие
–первое свидетельство гибели клетки. Это легко проиллюстрировать на примере свеклы. Если живой корень свеклы погрузить в холодную воду, то он сохраняет свой пигмент; если же свеклу кипятить, то клетки погибают, становятся легко проницаемыми и теряют пигмент, который и окрашивает воду в красный цвет.
Крупные молекулы типа белковых клетка может «заглатывать». Под влиянием некоторых белков, если они присутствуют в жидкости, окружающей клетку, в клеточной мембране возникает впячивание, которое затем смыкается, образуя пузырек – небольшую вакуоль, содержащую воду и белковые молекулы; после этого мембрана вокруг вакуоли разрывается, и содержимое попадает внутрь клетки. Такой процесс называется пиноцитозом (буквально «питье клетки»), или эндоцитозом.
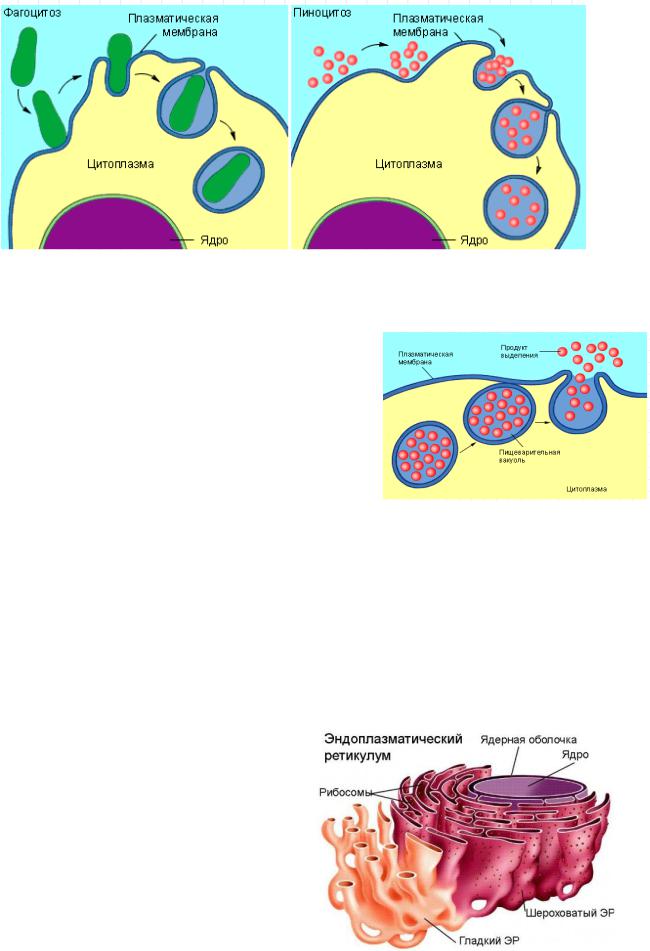
Более крупные частички, например частички пищи, могут поглощаться аналогичным образом в ходе т.н. фагоцитоза. Как правило, вакуоль, образующаяся при фагоцитозе, крупнее, и пища переваривается ферментами лизосом внутри вакуоли до разрыва окружающей ее мембраны. Такой тип питания характерен для простейших, например для амеб, поедающих бактерий.
Экзоцитоз (экзо — наружу), благодаря нему, клетка выводит внутриклеточные продукты или непереваренные остатки, заключенные в вакуоли, или пузырьки. Пузырек подходит к цитоплазматической мембране, сливается с ней, а его содержимое выделяется в окружающую среду. Так выделяются пищеварительные ферменты, гормоны, гемицеллюлоза и др..
Структура цитоплазмы.
Жидкую составляющую цитоплазмы также называют цитозолем. Под световым микроскопом казалось, что клетка заполнена чем-то вроде жидкой плазмы или золя, в котором «плавают» ядро и другие органоиды. На самом деле это не так. Внутреннее пространство эукариотической клетки строго упорядочено. Передвижение органоидов координируется при помощи специализированных транспортных систем, так называемых микротрубочек, служащих внутриклеточными «дорогами», и специальных белков динеинов и кинезинов, играющих роль «двигателей». Отдельные белковые молекулы также не диффундируют свободно по всему внутриклеточному пространству, а направляются в необходимые компартменты при помощи специальных сигналов на их поверхности, узнаваемых транспортными системами клетки.
Эндоплазматический ретикулум
В эукариотической клетке существует система переходящих друг в друга мембранных отсеков (трубок и цистерн),
которая называется эндоплазматическим ретикулумом (или эндоплазматическая сеть, ЭПР или ЭПС). Ту часть ЭПР, к мембранам которого прикреплены рибосомы, относят к гранулярному (или шероховатому) эндоплазматическому
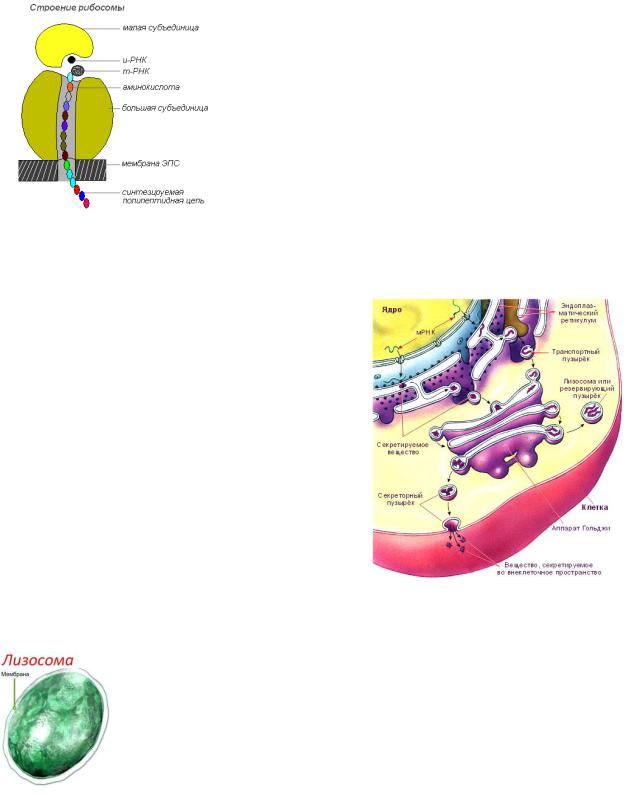
ретикулуму, на его мембранах происходит синтез белков. Те компартменты, на стенках которых нет рибосом, относят к гладкому ЭПР, принимающему участие в синтезе липидов. Внутренние пространства гладкого и гранулярного ЭПР не изолированы, а переходят друг в друга и сообщаются с просветом ядерной оболочки. Канальцы открываются и на поверхности клетки, и эндоплазматический ретикулум, таким образом, играет роль аппарата, через который внешняя среда может непосредственно взаимодействовать со всем содержимым клетки.
Крошечные тельца, называемые рибосомами, покрывают поверхность шероховатого эндоплазматического ретикулума, особенно вблизи ядра. Диаметр рибосом около 15 нм. Каждая рибосома состоит из двух неодинаковых по размерам частиц, малой и большой Их основная функция – синтез белков; к их поверхности прикрепляются матричная (информационная) РНК и аминокислоты, связанные с транспортными РНК. Синтезированные белки сначала накапливаются в каналах и полостях эндоплазматической сети, а затем транспортируются к органоидам и участкам клетки, где они потребляются.
Аппарат Гольджи
Аппарат Гольджи (комплекс Гольджи)
представляет собой стопку плоских мембранных мешочков, несколько расширенных ближе к краям. В цистернах аппарата Гольджи созревают некоторые белки, синтезированные на мембранах гранулярного ЭПР и предназначенные для секреции или образования лизосом. Аппарат Гольджи асимметричен — цистерны располагающиеся ближе к ядру клетки (цис-Гольджи) содержат наименее зрелые белки, к этим цистернам непрерывно присоединяются мембранные пузырьки — везикулы, отпочковывающиеся от эндоплазматического ретикулума. По-видимому, при помощи таких же пузырьков происходит дальнейшее перемещение созревающих белков от одной цистерны к другой. В конце концов от противоположного конца органеллы
(транс-Гольджи) отпочковываются пузырьки, содержащие полностью зрелые белки.
Лизосомы
Лизосомы (греч. «лизео» — растворяю, «сома» — тело) представляют собой небольшие округлые тельца. Эти мембранные органоиды клетки имеют овальную форму и диаметр 0,5 мкм Они отпочковываются от аппарата Гольджи и, возможно, от эндоплазматического ретикулума. Лизосомы содержат разнообразные ферменты, которые расщепляют крупные молекулы: белки, жиры, углеводы, нуклеиновые кислоты. Из-за своего разрушительного действия эти ферменты как бы «заперты» в лизосомах и высвобождаются только по мере надобности. Но если лизосома
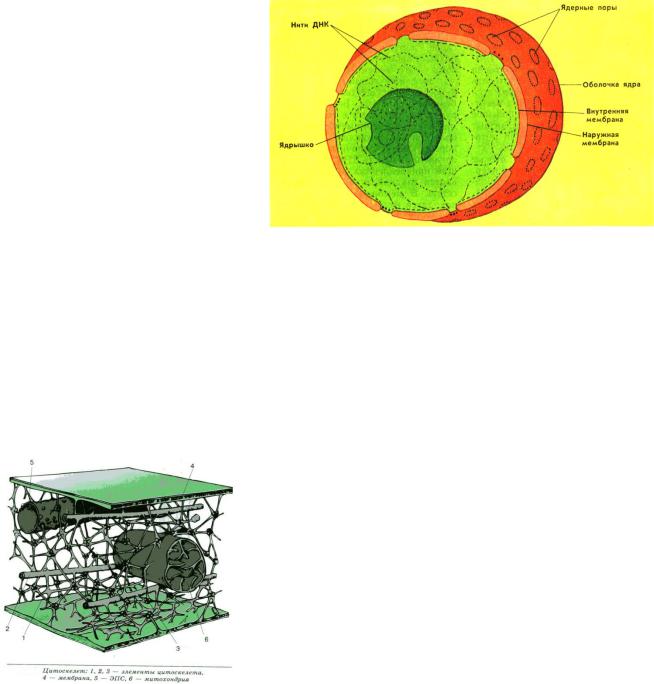
повреждается от каких-либо внешних воздействий, то разрушается вся клетка или часть ее.
При внутриклеточном пищеварении ферменты выделяются из лизосом в пищеварительные вакуоли.
При голодании клетки лизосомы переваривают некоторые органоиды, не убивая клетку. Такое частичное переваривание обеспечивает клетке на какое-то время необходимый минимум питательных веществ.
Обладая способностью к активному перевариванию пищевых веществ, лизосомы участвуют в удалении отмирающих в процессе жизнедеятельности частей клеток, целых клеток и органов. Например, исчезновение хвоста у головастика лягушек происходит под действием ферментов лизосом.. В данном случае это нормально и полезно для организма, но иногда такое разрушение клеток носит патологический характер. Например, при вдыхании асбестовой пыли она может проникнуть в клетки легких, и тогда происходит разрыв лизосом, разрушение клеток и развивается легочное заболевание.
Ядро
Информационным центром клетки, местом хранения и воспроизводства наследственной информации, которая определяет все признаки данной клетки и организма в целом, является ядро. Удаление ядра из клетки, как правило, ведет к ее быстрой гибели. Форма и размеры ядра клетки очень изменчивый зависят от вида организма, а также от типа, возраста и функционального состояния клетки. Общий план
строения ядра одинаков у всех клеток эукариот. Клеточное ядро состоит из ядерной оболочки, ядерного матрикса (нуклеоплазмы), хроматина и ядрышка (одного или нескольких). От цитоплазмы содержимое ядра отделено двойной мембраной или так называемой ядерной оболочкой. Наружная мембрана в некоторых местах переходит в каналы эндоплазматического ретикулума; к ней прикреплены рибосомы. Клеточное ядро содержит молекулы ДНК, на которых записана генетическая информация организма. . Этим определяется ведущая роль клеточного ядра в наследственности. В ядре происходит репликация — удвоение молекул ДНК, а также транскрипция— синтез молекул РНК на матрице ДНК. Сборка рибосом также происходит в ядре, в специальных образованиях, называемых ядрышками. Ядерная оболочка пронизана множеством пор, диаметр которых около 90 нм. Благодаря наличию пор, обеспечивающих избирательную проницаемость, ядерная оболочка контролирует обмен веществ между ядром и цитоплазмой.
|
Цитоскелет |
К элементам |
цитоскелета относят белковые |
фибриллярные структуры, расположенные в цитоплазме клетки: микротрубочки, актиновые и промежуточные филаменты. Микротрубочки принимают участие в транспорте органелл, входят в состав жгутиков, из микротрубочек строится митотическое веретено деления. Актиновые филаменты необходимы для поддержания
формы клетки, псевдоподиальных реакций. Роль промежуточных филаментов, повидимому, также заключается в поддержании структуры клетки. Белки цитоскелета составляют несколько десятков процентов от массы клеточного белка.
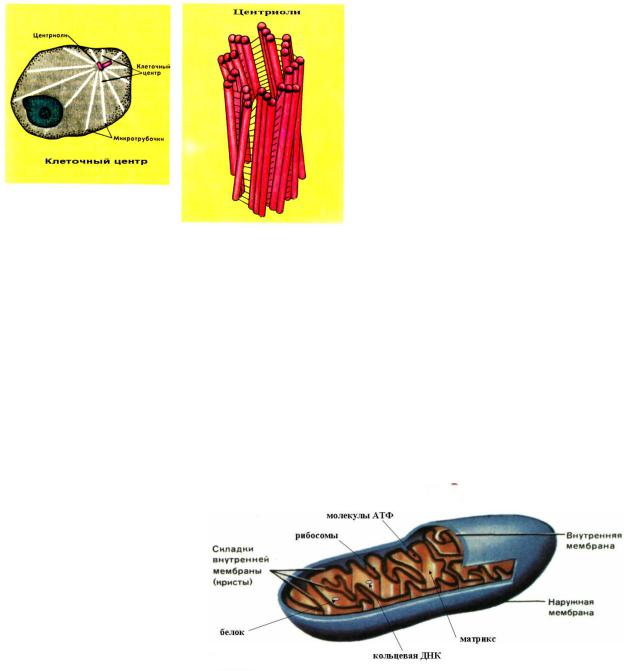
Центриоли
Центриоли представляют собой цилиндрические белковые структуры, расположенные вблизи ядра клеток животных (у растений центриолей нет, за исключением низших водорослей). Центриоль представляет собой цилиндр, боковая поверхность которого образована девятью наборами микротрубочек. Количество микротрубочек в наборе может
колебаться для разных организмов от 1 до 3.
Вокруг центриолей находится так называемый центр организации цитоскелета, район в котором группируются минус концы микротрубочек клетки.
Перед делением клетка содержит две центриоли, расположенные под прямым углом друг к другу. В ходе митоза они расходятся к разным концам клетки, формируя полюса веретена деления. После цитокинеза каждая дочерняя клетка получает по одной центриоли, которая удваивается к следующему делению. Удвоение центриолей происходит не делением, а путѐм синтеза новой структуры, перпендикулярной существующей.
Митохондрии
Митохондрии — особые органеллы клетки, основной функцией которых является синтез АТФ — универсального носителя энергии. В митохондриях протекает окисление органических веществ, сопряженное с синтезом
аденозинтрифосфата (АТФ). Распад АТФ с образованием аденозиндифосфата (АДФ) сопровождается выделением энергии, которая расходуется на различные процессы жизнедеятельности, например на синтез белков и нуклеиновых кислот, транспорт веществ внутрь клетки и из нее, передачу нервных импульсов или мышечное сокращение.
Митохондрии, таким образом, являются энергетическими станциями, перерабатывающими «топливо» – жиры и углеводы – в такую форму энергии, которая может быть использована клеткой, а следовательно, и организмом в целом.