

Экологические системы
Экологическая система.
По определению английского ботаника А. Тенсли (1935) экосистемой называют любую устойчивую совокупность организмов и неорганических компонентов, в которой может осуществляться круговорот веществ, т.е. живое сообщество образует с неорганической средой определенную систему, в которой поток атомов, вызываемый жизнедеятельностью организмов, имеет тенденцию замыкаться в круговорот. А. Тенсли подчеркивал, что неорганические и органические факторы выступают как равноправные компоненты.
Биотическая структура экосистемы.
Экосистема имеет определенную функциональную структуру, включающую группы организмов, различаемые по способу питания – автотрофы и гетеротрофы.
Автотрофы (самопитающие) — организмы, образующие органическое вещество своего тела из неорганических веществ посредством фотосинтеза и хемосинтеза. Фотосинтез осуществляют фотоавтотрофы — все хлорофиллоносные (зеленые) растения и микроорганизмы. Хемосинтез наблюдается у некоторых бактерий, использующих в качестве источника энергии окисление водорода, серы, сероводорода, аммиака, железа. Хемоавтотрофы в природных экосистемах играют относительно небольшую роль, за исключением чрезвычайно важных нитрифицирующих бактерий. Автотрофы составляют основную массу всех живых существ, отвечают за образование всего нового органического вещества в любой экосистеме, являются производителями продукции — продуцентами экосистем.
Гетеротрофы (питающиеся другими) — организмы, потребляющие органическое вещество других организмов и продуктов их жизнедеятельности (животные, грибы и большая часть бактерий). Гетеротрофы в экосистемах выступают как потребители – консументы и деструкторы – редуценты (разрушители) органических веществ.
Для поддержания круговорота веществ в экосистеме необходимо соблюдение условий:
наличие запаса неорганических веществ (молекул) в усвояемой форме;
3 функционально различные экологические группы организмов: (продуцентов, консументов, редуценты).
Биотический круговорот веществ и энергии в природных экосистемах
Запасы биогенных элементов, из которых строят тела живые организмы, на Земле в целом и на каждом конкретном участке на ее поверхности небезграничны. Лишь система круговоротов могла придать этим запасам свойство бесконечности, необходимое для продолжения жизни. Поддерживать и осуществлять круговорот могут только функционально различные группы организмов. Главный участник биотического круговорота – углерод, как основа органических веществ.
Круговорот углерода.
Наибольшее количество углерода содержится в литосфере в виде двух больших резервуаров, имеющих разное химическое происхождение:
в составе осадочных карбонатов, всех форм МСО3;
в составе ископаемых топлив, большая часть которых представляет собой биогенное и биокосное вещество — фоссилизированную, т.е. погребенную и преобразованную органику (от англ, fossil — окаменелый, ископаемый) — уголь, нефть, газ, нефтеносные сланцы, битуминозные пески, асфальт.
Небольшое количество углерода содержится в литосфере в виде графита и алмазов.
В гидросфере углерод находится в основном в виде гидрокарбоната НСО3, растворенного диоксида СО2 и некоторого количества растворимой органики.
В атмосфере – в виде СО2 и небольшой доли примесных газов, пыли и аэрозолей, содержащих углерод.
Вся мертвая органика, задействованная в биотическом круговороте, все формы детрита, органика почв и илов отнесены к биосфере.
Биомасса живых организмов содержит меньше 0,001% углерода Земли, но практически полностью определяет его планетарный круговорот.
Основная часть круговорота углерода происходит между биотой суши и океана и атмосферой. По сравнению с этими потоками остальные потоки малы. Но так как их действие охватывало огромные по длительности геологические эпохи, связанные с этими потоками, утечки привели к образованию гигантских масс ископаемых топлив и осадочных карбонатов.
Фоссилизация биогенного углерода, пик которой относится к каменноугольному периоду палеозойской эры (карбон — 350 — 290 млн. лет назад), в настоящее время не происходит. Идет противоположный процесс, в тысячи раз более мощный — техногенное сжигание ископаемых топлив.
Круговорот кислорода
В отличие от углерода, резервуары доступного для биоты кислорода по сравнению с его потоками огромны. Поэтому отпадает проблема глобального дефицита О2 и замкнутости его круговорота. Биотический круговорот кислорода составляет 270 Гт/год. Кислород на Земле — первый по распространенности элемент , его содержание (вес. %): в атмосфере — 23,1; в биосфере (в составе сухой органики) — 44,8; в литосфере — 47,2; в гидросфере (в составе воды) — 86,9. Для водных организмов нужен растворенный в воде кислород. Его среднее содержание в фотическом слое гидросферы составляет 4,5 мг/л и значительно колеблется.
Содержание кислорода в атмосфере во много раз больше — 288 мг/л — и на протяжении длительной геологической эпохи постоянно. Наземные животные довольно чувствительны к отклонениям от этого уровня. Некоторый дефицит кислорода для животных и человека возникает только в высокогорье, в зонах интенсивного потребления и в искусственных устройствах. Биота биосферы, сыгравшая решающую роль в оксигенизации атмосферы, подвела его концентрацию в воздухе до черты, за которой уровень окислительной способности среды становится уже опасным для биоты.
С круговоротом кислорода тесно связано образование озона. В высоких слоях атмосферы под влиянием жесткой ультрафиолетовой части солнечного спектра происходит ионизация и диссоциация части молекул кислорода, образуется атомарный кислород, который немедленно присоединяется к возбужденным молекулам кислорода, образуя озон — трехатомный кислород:
На образование озона тратится около 5 % поступающей к Земле солнечной энергии – около 8,6∙1015 Вт. Реакции легко обратимы. При распаде озона эта энергия выделяется, за счет чего в верхних слоях атмосферы поддерживается высокая температура. Средняя концентрация озона в атмосфере составляет около 10-6 объемных процентов; максимальная концентрация О3 – до 4∙10-6 объемных процентов достигается на высотах 20 – 25 км. Поглощая при своем образовании значительную часть жестких ультрафиолетовых лучей, озон играет большую защитную роль для всей экосферы.
Для некоторых экосистем вынос вещества за их пределы настолько велик, что их стабильность поддерживается в основном за счет притока такого же количества вещества извне, тогда как внутренний круговорот малоэффективен. Таковы проточные водоемы, реки, ручьи, участки на крутых склонах гор. Другие экосистемы имеют значительно более полный круговорот веществ и относительно автономны (леса, луга, озера и т. п.). Но ни одна, даже самая крупная экосистема Земли не имеет полностью замкнутого круговорота веществ. Материки обмениваются веществом с океанами, литосферой (при участии в этих процессах атмосферы), а вся наша планета часть материи и энергии получает из космоса, а часть отдает в космос.
Поток энергии через экосистемы.
В конечном счете, вся жизнь на земле существует за счет энергии солнечного излучения, которая переводится фотосинтезирующими организмами в химическую энергию органических соединений:
|
|
Процессы фотосинтеза и дыхания в экосистемах ΔGi — энергия солнечного света, потребленная в процессе фотосинтеза; ΔHe — энергия окисления органических веществ (дыхания), выделенная в виде теплоты |
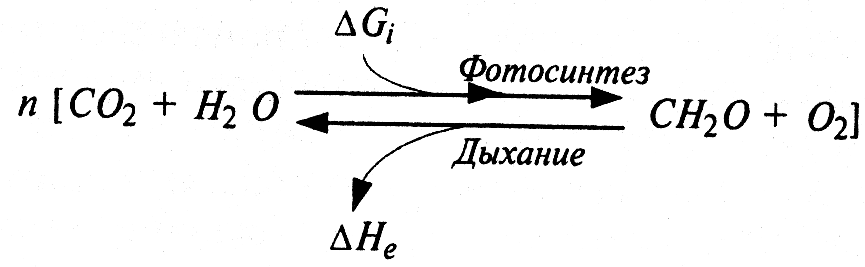
Количества поглощенной и выделенной энергии в реакции равны: ΔGi = ΔHe ≈ 478 кДж/моль. Общий множитель n определяет масштаб преобразований вещества и энергии в экосистеме. Прямая реакция полностью, а обратная на 80—90% обеспечиваются растениями. Остальное количество органического вещества потребляется гетеротрофными организмами — животными, грибами, бактериями — и расходуется в процессе их дыхания.
Принципиальное различие между потоками вещества и энергии в экосистеме заключается в том, что биогенные элементы, составляющие органическое вещество, могут многократно участвовать в круговороте веществ, тогда как поток энергии однонаправлен и необратим. Каждая порция энергии используется только однократно. В соответствии со вторым началом термодинамики на каждом этапе переноса и трансформации энергоносителей обязательно происходит сток части энергии: значительная ее часть неизбежно теряется, рассеивается в виде теплоты.
Пищевые (трофические) цепи и сети
Пищевые (трофические) цепи в сообществах – это механизмы передачи энергии от одного организма к другому. Типы пищевых цепей:
Пастбищные пищевые цепи, или цепи эксплуататоров, начинаются с продуцентов; для таких цепей при переходе с одного трофического уровня на другой характерно увеличение размеров особей при одновременном уменьшении плотности популяций, скорости размножения и продуктивности по биомассе. Например, «трава → полевки → лисица» или «трава → кузнечик → лягушка → цапля → коршун». Это наиболее распространенные цепи питания.
Цепи паразитов («яблоня → щитовка → наездник»; «корова → слепень → бактерии → фаги») характеризуются уменьшением размеров особей при увеличении численности, скорости размножения и плотности популяций.
Детритные цепи, включающие только редуцентов («опавшие листья → плесневые грибы → бактерии»), сходны с цепями паразитов.
Пищевые сети образуются потому, что практически любой член какой-либо пищевой цепи одновременно является звеном и в другой пищевой цепи: он потребляет и его потребляют несколько видов других организмов.
Совокупности трофических уровней различных экосистем моделируются с помощью трофических пирамид чисел (численностей), биомасс и энергий.
|
|
Простая трофическая пирамида (по Ю. Одуму, 1975) А – пирамида чисел; Б – пирамида биомасс; В – пирамида энергий.
|
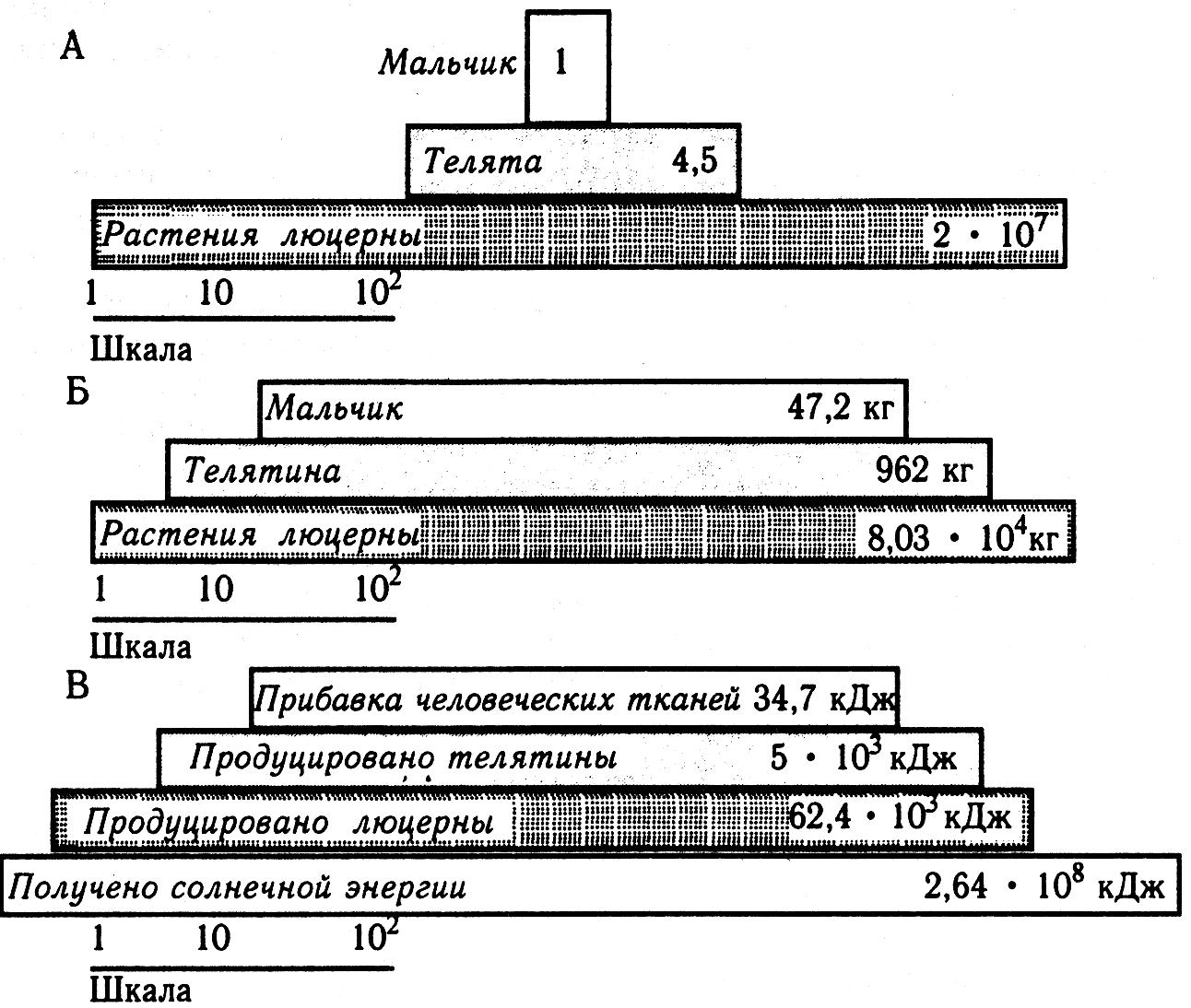
В пирамиде чисел отображают число особей на каждом из трофических уровней экосистемы. Для пастбищных цепей пирамиды имеют очень широкое основание (большое число продуцентов) и резкое сужение к конечным консументам, при этом числа «ступеней» различаются на 1–3 порядка. Но это справедливо только для травяных сообществ – луговых или степных биоценозов. Картина резко искажается, если рассматривать лесное сообщество (на одном дереве могут кормиться тысячи фитофагов) или если на одном трофическом уровне оказываются такие разные фитофаги, как тля и слон. Это искажение можно преодолеть с помощью пирамиды биомасс. В наземных экосистемах биомасса растений всегда существенно больше биомассы животных, а биомасса фитофагов всегда больше биомассы зоофагов. Для водных, особенно морских экосистем: биомасса животных обычно намного больше биомассы растений. Эта «неправильность» обусловлена тем, что пирамидами биомасс не учитываются продолжительность существования поколений особей на разных трофических уровнях и скорость образования и выедания биомассы. Главным продуцентом морских экосистем является фитопланктон. В океане за год может смениться до 50 поколений фитопланктона. За то время, пока хищные рыбы (и более крупные моллюски и киты) накопят свою биомассу, сменится множество поколений фитопланктона, суммарная биомасса которых намного больше. Поэтому универсальным способом выражения трофической структуры экосистем являются пирамиды скоростей образования живого вещества, продуктивности. Их обычно называют пирамидами энергий, имея в виду энергетическое выражение продукции, хотя правильнее говорить о мощности. Человек может участвовать в данной трофической цепи на любом уровне, начиная со второго.
Правило 10%: с каждого уровня трофической цепи на последующий переходит около 10% энергии.
Правило 1%: соотношение конечных консументов и их вклад в деструкцию (< 1%) является важным условием стабильности биосферы. Для биосферы в целом доля возможного конечного потребления чистой первичной продукции в энергетическом выражении не должна превышать одного процента. Для отдельных экологических систем порог нарушения стационарного состояния эмпирически оценивается на уровне не выше 5 % отклонения от нормального протока энергии (Н.Ф. Реймерс, 1994).
Следствие №1: трофические цепи не бывают длинными (обычно – 4–5 звеньев).
Следствие №2: эффект накапливания в трофических цепях вредных веществ, трудновыводимых из живых организмов (тяжелые металлы, ДДТ и др.)
Развитие и устойчивость экосистем. Сукцессия.
Изменения в любом сообществе, подразделяется на циклические и поступательные:
Циклические изменения сообществ отражают суточную, сезонную и многолетнюю периодичность внешних условий и проявления эндогенных ритмов организмов.
Поступательные изменения сообществ происходят под действием внешних, длительно действующих в одном направлении факторов.
Различают поступательные изменения:
экзогенетические смены (под действием внешнего фактора), например – дигрессия;
эндогенетические смены (под действием внутреннего фактора), например – сукцессия.
Дигрессия – вид экзогенетической смены биоценоза, при котором упрощается, обедняется его состав. Пример – превращение степей Нижнего Днепра в сыпучие пески под влиянием неумеренного выпаса скота.
Сукцессия – процесс саморазвития сообщества (от лат. successio — преемственность, последовательность). Причина сукцессии – неполнота биологического круговорота в биоценозе, в результате которого живое меняет вокруг себя среду, изымая из нее ряд веществ и насыщая ее продуктами метаболизма. Результат сукцессии – изменение окружающей среды в неблагоприятную стороны и вытеснение вида популяциями других видов, для которых изменение экологической среды является выгодными (конкуренция). Обычно наблюдается сукцессионная серия – развитие сообществ, в котором виды последовательно сменяют друг друга.
Различают виды сукцессий:
первичная сукцессия — постепенное заселение организмами появившейся девственной суши, оголенной материнской породы (отступившее море или ледник, высохшее озеро, песчаные дюны, голые скалы и застывшая лава после вулканического извержения и т.п.);
вторичная сукцессия имеет характер постепенного восстановления свойственного данной местности сообщества после нанесенных повреждений (последствий бури, пожара, вырубки, наводнения, выпаса скота, запуска полей). Возникшая в результате вторичной сукцессии система может существенно отличаться от первоначальной, если изменились некоторые характеристики ландшафта или климатические условия.
Все более или менее регулярные изменения экосистем, не выходящие за границы устойчивости экосистемы – ее обычного размера, видового состава, биомассы, продуктивности, характеризуют климаксное состояние экосистемы.
Постоянство важнейших экологических параметров обозначают как гомеостаз экосистемы.
Устойчивость экосистемы, как правило, тем больше, чем больше она по размеру и чем богаче и разнообразнее ее видовой и популяционный составы.
Длительное существование биоценоза возможно лишь в том случае, если изменения среды, вызванные деятельностью одних организмов, точно компенсируются деятельностью других. Антропное воздействие на среду – мощнейший фактор изменения условий обитания, он может привести к невозможности существования человека во вновь созданных им же условиях.
Агроэкосистемы – особый вид экосистем, создаваемых человеком для получения высокой чистой продукции автотрофов, отличаются от природных экосистем рядом особенностей:
Резко снижено разнообразие организмов, как растительных, так и животных. Видовое разнообразие разводимых человеком животных ничтожно мало по сравнению с природным видами.
Культивируемые человеком виды поддерживаются искусственным отбором в состоянии, неспособном выдерживать борьбу за существование с дикими видами без поддержки человека
Агроэкосистемы получают дополнительный поток энергии и веществ кроме солнца и природных веществ. Продукция удаляется из экосистемы и не поступает в цепи питания. Вредители – уничтожаются.
В настоящее время около 30% суши занято пахотными землями и пастбищами, деятельность людей по поддержанию этих систем становится глобальным экологическим фактором.
Условия идеальной агроэкосистемы: высокопродуктивность и стабильность с экологической точки зрения несовместимы. Искусственная регуляция численности вредителей – необходимое экономическое условие содержания агроэкосистем. Применение мощных ядохимикатов, гербицидов, пестицидов и т.д. приводит к «бумеранг–эффекту» – часто еще более мощной вспышке нашествия вредителей в результате подавления, уничтожения не только самого вредителя, но и его естественных врагов.