

Лекция 5. Электрофизиология нервного ствола
Нервный ствол (или нерв) представляет собой пучок множества (тысячи) нервных волокон, одетых общими эпителиальной и соединительнотканной оболочками. Обычно нерв включает в себя волокна различного типа и разного диаметра. Например, в седалищном нерве лягушки имеются двигательные волокна (аксоны мотонейронов), чувствительные (аксоны чувствительных клеток) и вегетативные (аксоны нейронов симпатических ганглиев). Моторные и наиболее толстые чувствительные волокна этого нерва миелинизированы, симпатические - не имеют миелиновой оболочки, но покрыты одним слоем шванновских клеток.
Все эти волокна в покое имеют поляризованную поверхностную мембрану. Как уже говорилось, если нерв перерезать и соединить с электрорегистрирующим устройством его поперечный разрез и продольную поверхность, то регистрируется потенциал повреждения или иначе потенциал покоя (ПП) нормальной точки нерва относительно поврежденной. ПП имеет обычно небольшую величину - 20-30 мВ, он меньше МПП волокон нерва, что объясняется шунтирующим действием межволоконных щелевых пространств. Если в сегменте нерва, расположенном между электродами, заменить (перфузией) межклеточную жидкость на раствор сахарозы ("сахарозный мостик"), то ПП нерва поднимается до -70 мВ. Его величина, по существу, отражает величину МПП наиболее толстых ?-волокон группы А, генерирующих основную часть внешнего тока.
Потенциалы действия различных групп волокон нервных стволов позвоночных исследованы в классических работах Эрлангера и Гассера (1937). Авторы установили, что в нервах лягушки и кошки имеются три основные группы волокон (А, В и С), различающиеся по электровозбудимости, по скорости развития и компенсации ПД и по скорости проведения (все эти показатели в ряду А-В-С падают). Группа А включает наиболее толстые хорошо миелинизированные моторные и чувствительные волокна; В - слабо миелинизированные преганглио-нарные (парасимпатические) волокна; С - немиелинизирован-ные постганглионарные (симпатические) волокна. Группа А неоднородна, в ней по тем же показателям можно выделить подгруппы; в этом ряду названные показатели тоже падают. Заметим, что указанные соотношения порогов электрического раздражения групп волокон не отражают точного соотношения электровозбудимости их мембран. Высокие относительные пороги тонких волокон при их раздражении в нервном стволе определяются в основном тем обстоятельством, что тонкие волокна по сравнению с толстыми обладают более высоким внутренним сопротивлением. В них входит такая малая часть раздражающего тока, что при пороговой силе для Aα-волокон, она совершенно недостаточна для создания на мембране более тонких волокон сколько-нибудь существенной деполяризации. За вычетом этого фактора различия электровозбудимости А, В, и С групп волокон, по-видимому, гораздо менее значительны.

Суммарная (многоволоконная) электрическая активность нерва (нервного ствола) создается его волокнами, каждое из которых генерирует свой стандартный по амплитуде и временным параметрам ПД, распространяющийся в обе стороны от точки, к которой приложено раздражение. Суммарный электрический сигнал нерва зависит от числа активных волокон, синхронности их активности, способа отведения и других обстоятельств.
Рассмотрим случай так называемого однофазного отведения, при котором один отводящий электрод (активный) стоит на нормальном участке ствола, а другой (индифферентный) на поврежденном участке, где волокна деполяризованы полностью (рис.5.1.). Пусть применяется искусственное раздражение и раздражающий электрод (катод) расположен достаточно близко (l==3 мм) от активного отводящего электрода, а нерв находится в непроводящей среде - масле или воздухе. В этой ситуации, когда сильное одиночное раздражение приводит к синхронному возбуждению всех волокон, активным отводящим электродом регистрируется суммарный ПД нерва, по форме приближающийся к ПД отдельного α-волокна, но немного более затянутый во времени. Этот ПД нерва, однако, не подчиняется правилу "все или ничего", но зависит от силы раздражения: на пороге ПД ничтожно мало, с увеличением силы стимулов он постепенно растет, достигая максимума, равного при обычных условиях нескольким милливольтам (5-10 мВ), а в условиях сахарозного мостика 50-100 мВ. При дальнейшем увеличении силы стимулов этот ПД несколько удлиняется во времени. Все эти изменения амплитуды и длительности пика ПД нерва
![]()
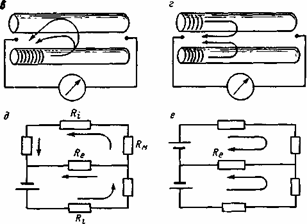
Рис.5.1 . Развитие пика составного потенциала действия нервного ствола.
а - схема раздражения и однофазного отведения ПД нервного ствола, перфузированного высокоомным раствором сахарозы, б - рост амплитуды составного ПД при усилении стимулов. Схемы петления: в - тока одиночного возбужденного волокна на окружающие его покоящиеся волокна, г - токов двух соседних возбужденных волокон, д и е - эквивалентные схемы двух соседних волокон при возбуждении одного из них (д) и при синхронной активации обоих в соседних точках (е).
при усилении стимула определяются ростом числа активных волокон, подключением к низкопороговым и быстрым а-волокнам, более высокопороговых медленных β, γ, Δ волокон группы А, а затем В и, наконец С-группы.
Но рост амплитуды суммарного ("составного") ПД нерва при увеличении числа синхронно активных волокон не имеет в своей основе истинного сложения их ЭДС. Волокна в нерве подключены не последовательно (лишь тогда их ЭДС бы складывались), а параллельно. Причину роста составного ПД при увеличении числа активных волокон можно понять, рассмотрев схему на рис. , из которой видно, что неактивные волокна шунтируют активные. При активизации неактивных волокон этот шунт снимается. Кроме того, при этом падает суммарное внутреннее сопротивление источника .
Рассмотрим случай, когда при тех же прочих условиях существенно (до 80-100 мм) увеличено расстояние между раздражающим электродом (катодом) и активным отводящим электродом (рис.5.2 ). При этом на пороговые раздражения и

Рис.5.2. Схемы дисперсии компонентов составного однофазного потенциала действия нерва (связанных с А (α, β, γ, Δ) В- и С-волокнами) и формирования двухфазного потенциала действия.
а, б - варианты однофазного отведения ПД при малой (а) и большой (б) дистанциях проведения, в, г - формы составных ПД при данных дистанциях и разных силах раздражения (при большой дистанции и сверхпороговом раздражении пик составного потенциала больше и разделен на ряд последовательных зубцов, порождаемых волокнами с разными скоростями проведения), д, е - схемы двухфазного отведения (d) и формирования асимметричной двухфазной кривой (е) из однофазных сигналов двух отводимых точек (1 и 2).
при небольшом превышении порога получается в общем такой же Аα-пик. Но по мере роста предъявляемых стимулов этот Аα--пик не просто увеличивается, а осложняется рядом дополнительных более поздних пиков, следующих за Аα-пиком. Это Аβ-, Аγ и АΔ-, В и С-пики. Такое их распределение -результат отставания от Аα--импульса импульсов Аβ,γ и Δ, В и С волокон, называемое их дисперсией во времени. Степень ее у одиночного залпа первоначально синхронных импульсов тем больше, чем больше дистанция проведения и чем больше различия в скорости проведения у сигналов волокон разных групп (рис.5.2.).
Рассмотрим вариант так называемого двухфазного отведения ПД, при котором оба электрода расположены на нормальных неповрежденных участках нерва. Это отведение применяют в том случае, если нервный проводник почему-либо надо сохранить в целости. При двухфазном отведении картина сложна, даже если дистанция проведения мала и нет существенной временной дисперсии волоконных сигналов. Дело в том, что каждый из электродов при этом отводит свой составной ПД (ПД в своей точке) и эти два ПД алгебраически складываются на входе регистрирующего прибора. На схеме (рис.5.2.) видно, что при этом сложении получается двухфазная волна и что в силу несимметричности пиков ПД первая фаза этой волны выше

Рис.5. 3 . Схема отведения распространяющего пика потенциала действия нерва на некотором расстоянии от него в проводящей среде.
1-у левого края фильтровальной бумажки, 2 - на уровне средней части проводника, 3 - на конце нерва, 4 - у правого края фильтровальной бумажки.
второй, а вторая длиннее первой. При большей дистанции проведения эти фазы, кроме того, осложняются соответствующими дополнительными пиками.
Наконец, рассмотрим так называемое фокальное отведение (рис.5.3.) пика ПД нервного ствола, расположенного в проводящей среде. При таком отведении один электрод приближен к нерву, а второй (индифферентный) "бесконечно" удален. Этот случай моделирует регистрацию электрического сигнала нейрона или группы нейронов в мозге с помощью внеклеточного фокального микроэлектрода. Лоренте де Но проанализировал этот вариант отведения в опыте на седалищном нерве лягушки, расположенном на листе фильтровальной бумаги, смоченном в растворе Рингера. Зарегистрировав фокальным электродом потенциалы в разных точках проводящей среды в определенные моменты возбуждения нерва (например, при достижении вершины а-пика ПД в средней части нерва), автор смог получить картины электрического поля, возникающего в проводящей среде вокруг активного нерва (см. рис.5.3.). Представленная на рис. схема - это изображение электрического поля на плоскости. Но вращая рисунок вокруг оси нерва, можно получить соответствующее объемное поле. Обратим внимание на то, что нерв в тот момент, когда он оказывается возбужденным в средней части, имеет вид генератора с тремя полюсами - двумя плюсами по концам и минусом в средней части. И электрическое поле в среде имеет три соответственные зоны, разделенные на плоскости линиями (а в объеме поверхностями) нулевых сдвигов U. Заметим, что по мере удаления от нерва в любую сторону величины сдвигов U падают, на краях бумажки они близки к 0. Когда возбуждение нерва (его составной ПД) распространяется слева направо, то движется и порождаемое им электрическое поле в среде. При этом стоящий на средней линии фокальный электрод, мимо которого движется ПД,
регистрирует трехфазный сигнал, тем более слабый,чем дальше этот электрод отнесен от нерва (в поперечном к его оси направлении). Если отводящий фокальный электрод помещается в месте зарождения поля ПД, т. е. на нашей схеме на левом краю фильтровальной бумажки, где нерв на нее выходит из масляной ванночки (в которой он контактирует с раздражающими электродами), то при пробеге ПД регистрируется двухфазный сигнал . Если же фокальный электрод помещается на правом краю бумажки, куда нерв не дотягивается и ПД соответственно не доходит, то при пробеге ПД регистрируется однофазный сигнал положительной полярности. Заметим, что эти формы фокального электрического сигнала, порождаемого полем проводящегося ПД,- трехфазная, двухфазная и однофазная характерны соответственно для случаев прохода ПД мимо электрода, зарождения ПД около электрода и ухода от него и, наконец, движения ПД к электроду с остановкой "на полпути".
Все рассмотренные до сих пор варианты записи электронейрограмм получаются при синхронной электрической активности всех волокон нервного ствола в условиях искусственного электрического раздражения нерва.
Но в естественных условиях отдельные волокна нервного ствола получают раздражение (тоже электрическое) из своих индивидуальных источников, например моторные волокна, из тел мотонейронов, а чувствительные - из рецепторов. Волокна нервного ствола при этом работают чаще всего асинхронно, а чувствительные и двигательные, кроме того, проводят ПД в разных направлениях. При этом из-за действия шунтов суммарная электрическая активность нервного ствола оказывается очень ослабленной, ее анализ весьма труден.
В заключение коснемся вопроса об электрических взаимоотношениях работающих соседних волокон в нервных стволах. На схеме (рис.5.1.) видно, что соседние волокна подключены друг к другу, как шунты, через межклеточную щель. И если одно из этих волокон работает, то внешние петли тока, порождаемого его ПД, затекают в соседние волокна, используемые, таким образом, как часть внешней электропроводности (проводимости). Сила тока этих петель при активности малого числа волокон не достаточна для раздражения. Поэтому в условиях асинхронной активности волокна функционируют вполне изолированно друг от друга. Но при синхронной активации значительной части волокон нерва суммарное поле их ПД порождает более сильный ток, приближающийся к пороговому для волокон. При этом возможно, например, ускорение проведения импульсов по более медленным волокнам за счет добавления соседнего электрического влияния от более быстро проводящих. Такой эффект отчетливо показали Катц и Шмидт в 1940 г. на нерве ракообразного, содержащем всего 2 волокна. Если искусственно повысить электровозбудимость волокон нерва, то, как показал еще в 1937 г. Д. Г. Квасов, возбуждение части волокон влечет за собой "соседнее" электрическое раздражение остальных. Такой эффект между прочим лежит в основе уравнительной стадии парабиоза нерва, описанного Н. Е. Введенским.
Если синхронно и в одной точке нерва возбуждается большая часть его волокон, например все А-волокна, то получается еще один эффект. Происходит взаимное вычитание внешних петель биотоков соседних возбужденных волокон. Это приводит к снижению силы токов, связывающих возбужденные участки этих волокон с покоящимися, подлежащими раздражению. В итоге на несколько процентов падает скорость проведения их ПД и ГФ1 этого проведения. В нормальном проводнике такое снижение ГФ! практически не сказывается на функции. Но если нерв локально обработан каким-либо повреждающим агентом и в этом месте (участок парабиоза) его волокна исходно имеют ГФ1∼1, то синхронный залп ПД совокупности его волокон, распространившись до участка парабиоза, может не пройти через этот участок из-за рассмотренного дополнительного снижения ГФ. И чем больше волокон синхронно возбуждено (т. е. чем сильнее приложенное к нерву раздражение), тем вероятнее такой блок. В этом, по нашим данным, состоит основная причина парадоксальной стадии парабиоза нерва.