

2 курс / Нормальная физиология / Физиология_сердца_Алексеева_Э_А_,_Шантанова_Л_Н_
.pdfПД атипичных кардиомиоцитов (рис. 5):
•Максимальный диастолический потенциал в клетках СА-узла -50…-60 мВ, в клетках АВ-узла – 60…-70мВ;
•Медленно нарастающие ПД;
•Длительность – 100-300 мс;
•Амплитуда – 70-80 мВ.
Синусовый узел
Рис. 5. ПД атипичных кардиомиоцитов (i – проводимость мембраны для ионов)
Фазы ПД:
фаза деполяризации (фаза 0) имеет низкую скорость, обусловлена входящим током Na+ / Ca2+ в связи с открыванием высокопороговых Ca2+ -каналов L- типа;
фаза реполяризации (фаза 3) – инактивация Na+ -каналов, открытие K+ каналов, выход ионов K+ из клетки, в результате достигается максимальный диастолический потенциал (около –60 мВ), который нестабилен, так как мембрана начинает спонтанно деполяризоваться;
фаза медленной диастолической деполяризации (фаза МДД),
фаза 4 является показателем автоматии (пейсмекерным потенциалом) и последовательно формируется:
• сначала Na+ -каналами утечки, по которым Na+ входит в внутрь клетки;
11
•открыванием низкопороговых Сa 2+ -каналов Т- типа и входом Сa 2+ /Na+ внутрь клетки;
•спонтанным снижением проводимости мембраны для К+ ;
•в последней трети МДД активацией высокопороговых Сa 2+ - каналов L- типа.
1.2.3. Проводимость и проведение возбуждения в сердце
Свойство кардиомиоцитов проводить возбуждение от клетки к клетке называется проводимостью. ПД распространяется через участки тесного соприкосновения мембран (нексусы). Скорость, с которой ПД распространяется через участок сердечной мышцы, называется скоростью проведения. Она варьирует в различных участках миокарда и напрямую зависит от:
диаметра вовлеченных мышечных волокон;
скорости фазы деполяризации.
ПД, возникший в клетках SA узла, распространяется по предсердиям со скоростью 1 м/с. Имеется три тракта: короткий перед-
ний Бахмана (Bachmann), средний Венкебаха (Wenckebach) и
длинный задний Тореля (Thorel). Синхронное возбуждение обоих предсердий достигается тем, что импульс быстро распространяется на левое предсердие по межпредсердному пучку Бахмана — ветви переднего межузлового тракта.
АВ узел замедляет передачу возбуждения до 0,02-0,04 м/с, что определяет его основные функции, которые заключаются в следующем:
физиологическая задержка передачи возбуждения от предсердий к желудочкам (0,04 с) обеспечивает десинхронизацию их деятельности – сокращение предсердий предшествует сокращению желудочков;
защита желудочков от возможной (слишком частой) импульсации со стороны предсердий – АВ узел является "фильтром" на пути между предсердиями и желудочками;
защита желудочков от возможных (слишком ранних) предсердных импульсов, которые могли бы застать желудочки в уязвимой фазе;
защита желудочков от возможной длительной асистолии; когда предсердный импульс слишком запаздывает, АВ узел становится генератором желудочкового ритма.
12
Большой размер клеток и быстрая фаза деполяризации клеток пучка Гиса и волокон Пуркинье определяют самую высокую скорость проведения возбуждения в миокарде (4-2 м/с), что обеспечивает синхронность охвата возбуждением всей толщины миокарда желудочков, тем самым повышается мощность и эффективность сокращения сердца.
1.2.4. Сократимость и сокращение сердца
Сокращение рабочих кардиомиоцитов сердечной мышцы ини-
циируется потенциалом действия.
Функциональная структура типичного кардиомиоцита.
•Миофибриллы, саркомеры ответственны за механические процессы укорочения и напряжения;
•сократительные белки: миозин – образует толстые нити, обладает длинной прямой цепью с двумя глобулярными головками, каждая из которых содержит зону связывания с АТФ и актином; актин – образует тонкие нити, состоит из двух винтообразных тяжей с локусами (активными центрами) для взаимодействия с головками поперечных мостиков миозина; тропонин С – связывается с кальцием и инициирует изменение конфигурации регуляторных белков; тропонин Т – обеспечивает связь тропонина и тропомиозина; тропонин I – ингибирует взаимодействие актина и миозина в состоянии покоя; тропомиозин – регуляторный белок, расположенный в области активного центра актина, препятствующий взаимодействию актина и миозина вне сокращения.
•Т- система – впячивание сарколеммы внутрь клетки, соединяется с L-системой, обеспечивает проведение возбуждения внутрь миоцита к саркоплазматическому ретикулуму;
•L- система (саркоплазматический ретикулум) – депонируется кальций внутри клетки при помощи специфического белка – кальсеквестрина;
•Большое количество митохондрий, в которых осуществляются процессы окислительного фосфорилирования, обеспечивающие выработку АТФ для удовлетворения высоких метаболических потребностей сердечной мышцы.
13
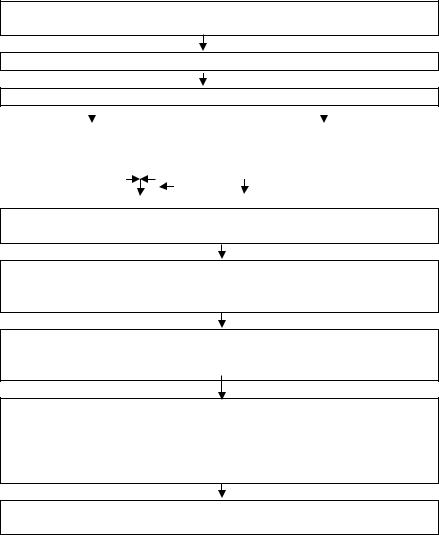
Электромеханическое сопряжение и механизм сокращения кардиомиоцитов (рис. 6)
ПД распространяется по сарколемме вдоль и внутрь по Т-системе
Открываются потенциалзависимые Са2+ каналы L-типа
Са2+ поступает внутрь клетки
Открывание хемоактивируемых |
Открывание метаботропных |
|
(рианодиновых) Са2+каналов |
(ИФ3)- зависимых Са2+каналов |
|
СПР |
|
СПР |
веропамил – |
+ кофеин + норадреналин |
|
Ca2+ поступает из СПР в цитозоль клетки, концентрация Ca2+ повышается с 10-7 до 10-5 М
Са2+ связывается с тропонином С, увеличивается степень спирализации тропомиозина,
открываются активные центры актина
Со свободным активным центром актина связывается головка поперечного мостика миозина, активация АТФ-азы миозина, гидролиз АТФ до АДФ и Pn
Отсоединение АДФ и Рn от головки миозина сопровождается выделением энергии (силовой удар), которая расходуется на поворот шейки на 45˚ «гребок», что вызывает перемещение актина вдоль миозина на 1 % длины саркомера
Присоединение АТФ к головке миозина вызывает разъединение актина и миозина
Значение тока Са2+ внутрь клетки:
увеличение длительности ПД;
увеличение продолжительности рефрактерного периода;
14

влияние на силу сокращения;
пополнение запаса внутриклеточного Са2+.
Рис. 6. Электромеханическое сопряжение, укорочение и расслабление саркомера
Механизм расслабления кардиомиоцитов
Активация Са2+ -АТФ-азы |
3Na+ /Ca2+– ионообменник + |
СПР (80 %) |
Са2+ -насос плазмолеммы |
|
(20%) |
Снижение концентрации Са2+внутри клетки до 10-7 М
Освобождение тропонина, тропомиозин закрывает активные центры актина
Актин и миозин свободно скользят относительно друг друга
15

1.3. Механическая работа сердца
Повторные синхронные сокращения и расслабления миокарда создают силы, необходимые для продвижения крови через большой и малый круги кровообращения.
Сердечный цикл – полная последовательность сокращения и расслабления сердца. При ЧСС 75 уд\мин продолжительность сердечного цикла составляет 0,8 с.
1.3.1. Структура кардиоцикла
Сердечный цикл начинается с систолы предсердий (0,1 с). Систола желудочков начинается одновременно с диастолой предсердий
(рис. 7).
Цикл работы желудочков (Шово и Морели (1861 г.)) состоит из систолы и диастолы желудочков.
Рис. 7. Структура кардиоцикла
16
Систола желудочков: период напряжения и период изгнания
(0,33 с)
Период напряжения осуществляется в 2 фазы:
1)фаза асинхронного сокращения (0,04 с) – неравномерное со-
кращение желудочков. Сокращение мышцы межжелудочковой перегородки и папиллярных мышц. Давление в желудочках растет, достигает 5-8 мм рт.ст., становится выше, чем в предсердиях. Кровь устремляется обратно в предсердия. Эта фаза заканчивается полным закрытием атриовентрикулярных клапанов.
2)фаза изометрического сокращения (0,04 с) – начинается с момента закрытия атриовентрикулярного клапана и протекает при закрытии всех клапанов. Поскольку кровь несжимаема, в эту фазу длина мышечных волокон не изменяется, а увеличивается их напряжение. В результате давление в желудочках растет: в левом желудочке – 120-130 мм рт.ст., в правом – 25 мм рт.ст, становится выше, чем в аорте и легочном стволе, кровь устремляется в сосуды, в итоге – открытие полулунных клапанов.
Период изгнания (0,25 с) состоит из 2-х фаз:
1)фаза быстрого изгнания (0,12 с) – начинается с открытия полулунных клапанов, кровь быстро поступает в аорту и легочной ствол, давление одновременно увеличивается как в желудочках, так
ив сосудах, так как желудочки продолжают сокращаться.
2)фаза медленного изгнания (0,13 с)
Основной фактор – разница давлений, что способствует выбросу крови. В этот период происходит изотоническое сокращение миокарда. В покое в фазу изгнания из желудочков выбрасывается 4050% от общего объема крови (130 мл), т.е. конечносистолический объем составляет 65-70 мл, который является резервным объемом.
Диастола желудочков (0,47 с). Состоит из следующих периодов:
Протодиастолический период (0,04 с) – интервал времени от начала расслабления желудочков до закрытия полулунных клапанов. Миокард желудочков расслабляется, давление в них падает и становится ниже, чем в аорте и легочном стволе, кровь за счёт разности давления возвращается в желудочки, наполняя кармашки полулунных клапанов, закрывает их.
Период изометрического расслабления (0,25 с) – осуществля-
ется при полностью закрытых клапанах. Длина мышечного волокна постоянна, изменяется их напряжение, давление в желудочках
17
уменьшается, становится ниже, чем в предсердиях. В результате движения крови открываются атриовентрикулярные клапаны.
Период наполнения (0,18 с) – осуществляется в общую паузу сердца. Сначала быстрое наполнение, затем медленное – желудочки наполняются на 2/3.
Пресистола – наполнение желудочков кровью за счет систолы предсердий (на 1/3 объёма).
1.4. Энергетика миокарда
Для того чтобы сердце функционировало должным образом, у него должен быть адекватный запас химической энергии в виде аденозинтрифосфата (АТФ). Основные субстраты, из которых в сердце может образовываться АТФ, могут варьировать в зависимости от того, какие вещества в данный момент в наибольшем количестве. Например, после приема пищи, обильной углеводами, ткани сердца будут захватывать и использовать глюкозу и пируват, а между приемами пищи ткани сердца могут переключаться на метаболизм свободных жирных кислот, триглицеридов и кетонов. Кроме того, гликоген запасается в клетках миокарда в качестве резервного источника энергии, он может быть мобилизован с помощью гликогенолиза для обеспечения дополнительного количества субстрата при наличии повышенной симпатической активности. [Катехоламины, взаимодействуя с мембранными β-рецепторами, увеличивают содержание в клетке циклического аденозинмонофосфата (цАМФ), который активирует фосфорилазу, стимулирующую метаболизм гликогена].
Конечным продуктом метаболизма гликогена, глюкозы, жирных кислот, триглицеридов, пирувата и лактата является ацетил Со А, который вступает в цикл трикарбоновых кислот (Кребса) в митохондриях, где в процессе окислительного фосфорилирования молекулы веществ распадаются до С02 и воды, а энергия переходит в АТФ.
Анаэробных источников энергии в сердце (например, процесс гликолиза, креатин-фосфат) недостаточно для удовлетворения метаболических потребностей более чем на несколько минут. То, что в тканях сердца продукция АТФ осуществляется почти полностью с помощью аэробных процессов, доказывается (1) большим количеством митохондрий в клетках сердечной мышцы и (2) наличием вы-
18
сокой концентрации кислородсвязывающего белка, миоглобина, в клетках миокарда. Миоглобин может передать содержащийся в нем кислород в систему митохондриальных цитохромоксидаз при снижении внутриклеточного уровня кислорода.
Процесс снабжения миокарда энергией складывается из 3-х этапов.
I этап – образование макроэргических соединений в митохондриях, в присутствии О2. Наиболее энергетически выгодный процесс
– окисление жирных кислот, 1 молекула жирных кислот даёт 130140 молекул АТФ. Частично используется окисление глюкозы: 1 молекула глюкозы – 30-35 молекул АТФ. При гипоксии образование энергии нарушается. Происходят анаэробные процессы. (1 молекула глюкозы даёт 2 молекулы АТФ). При гипоксии возможен некроз миокарда.
II этап – участвуют ферменты АТФ-, АДФ-транслоказа, который обеспечивает обменную функцию через мембрану митохондрий. Другой фермент – креатинфосфатаза, который находится на наружной поверхности митохондрии и в миофибриллах. На наружную поверхность митохондрии выделяется АТФ, которая взаимодействует с активной единицей креатинфосфотазы, здесь из АТФ и креатина в присутствии Mg2+ образуется АДФ и креатинфосфат, последний поступает к миофибриллам. Креатинфосфат – основной носитель энергии. На поверхности миофибрилл реакция идёт в обратном направлении. АТФ используется миофибриллой.
III этап. 75 % энергии идёт на сокращение и расслабление миокарда; 15 % – на работу Са2+ – насоса; 5 % – работу Nа+-К+ – насоса;
10 % – на синтез различных веществ.
Механическая работа сердца линейно зависит от скорости поглощения О2. В покое сердце потребляет 24-30 мл О2./ мин. При физической нагрузке увеличивается в 3-4 раза.
1.5. Кровоснабжение миокарда
Кровоснабжение миокарда осуществляется коронарными сосудами.
Левая коронарная артерия снабжает левый желудочек, межжелудочковую перегородку, большую часть правого и левого предсердий, синоатриальный узел. Правая коронарная артерия дает ветви к передней стенке правого желудочка, частично к переднему отделу
19
перегородки, обеим папиллярным мышцам правого желудочка, задней стенке правого желудочка и заднему отделу межжелудочковой перегородки; от нее также отходит вторая ветвь к синоаурикулярному узлу.
В норме коронарный кровоток составляет 1/20 минутного объема крови (приблизительно 250 мл в минуту). При нагрузке интенсивность коронарного кровотока может возрастать в 15-20 раз (приблизительно 4-5 л). Основной механизм – вазодилатация на повышенную нагрузку.
Анатомо-физиологические особенности коронарного кровотока:
1.Коронарные сосуды получают самую оксигенированную
кровь.
2.Интенсивность коронарного кровотока в значительной степени автономна и мало меняется при падении систолического давления.
3.Левая коронарная артерия наполняется кровью на 85 % в период диастолы. Правая коронарная артерия – в систолу.
4.Коронарные сосуды имеют анастомозы между мелкими сосудами 4-го и 5-го порядка, между правой и левой коронарной артериями, между коронарными артериями и внутренней грудной артерией. В норме эти анастомозы не функционируют, имеют очень маленький диаметр. Открываться могут только при внезапном закрытии магистрального сосуда.
5.В субэндокардиальной зоне коронарные сосуды узкие, извилистые, почти не имеют анастомозов, поэтому некроз возникает в этой зоне.
6.Из коронарных сосудов миокард экстрагирует до 75 % кислорода. Поэтому увеличение потребления кислорода в сердце обеспечивается в первую очередь увеличением коронарного кровотока (доставки О2).
Регуляция коронарного кровотока
Миогенная саморегуляция.
Симпатическая стимуляция ά-адренорецепторов вызывает кратковременное сужение коронарных сосудов и снижение кровотока, работа сердца увеличивается, что приводит к увеличению об-
20