

6.12.2. Аккомодация
Для ясного видения предмета необходимо, чтобы лучи от отдельных точек его были сфокусированы на сетчатке. Эту функцию и выполняют глазные мышцы, поворачивающие глаз, и преломляющие среды глаза.В обычных условиях преломляющая сила глаза молодого человека обеспечиваетфокусировку лучей,поступающих от далеко расположенного предмета на сетчатке. Близкие предметы при этом видны расплывчато, так как лучи от них сфокусированы за сетчаткой. Для того, чтобы ясно видеть близкие предметы, необходимо увеличить преломляющую силу глаза. Отсюда становится понятным, почему одновременно нельзя ясно видеть далекие и близкие предметы. Приспособление глаза к ясному видению различно удаленных предметов называетсяаккомодацией.
О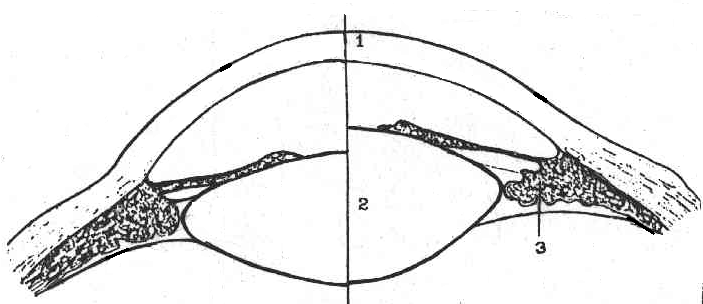 птическая
система имеет два основных приспособительных
механизма, позволяющих активно
регулировать интен-сивность светового
потока и его направленность - зрачок и
хрусталик.
птическая
система имеет два основных приспособительных
механизма, позволяющих активно
регулировать интен-сивность светового
потока и его направленность - зрачок и
хрусталик.
Рис. 6.19. Механизм аккомодации: 1 - роговица, 2 - выпуклый хрусталик при сокращении цилиарной мышцы - 3.
Аккомодация обеспечивается хрусталиком, кривизна которого может меняться (рис. 6.19). У молодого человека рефракционная возможность хрусталика может меняться от 15 D до 29 D. Хрусталик заключен в тонкую капсулу, переходящую по краям в циннову связку, радиальные волокна которой прикрепляются с другого конца к ресничному телу. Кривизна хрусталика зависит от взаимодействия сил эластичности его структур и упругости, возникающей в цилиарном аппарате и склере, к которой прикреплена циллиарная связка. Механическое натяжение склеры в свою очередь зависит от внутриглазного давления. Так как обычно волокна связки натянуты, то форма хрусталика менее выпуклая, чем свойственно его эластическим элементам.
Регуляция аккомодации
В регуляции натяжения циллиарного аппарата ведущее значение принадлежит цилиарной мышце. В ней имеется два типа волокон: меридианные и циркулярные. Меридианные волокна тянутся от места перехода роговицы в склеру до цинновой связки и при своем сокращении "снимают" натяжение цинновой связки. Активно уменьшается натяжение связки и при сокращении циркулярных волокон. В результате под влиянием эластических сил хрусталика кривизна его увеличивается.Диапазон аккомодации(интервал изменения преломляющей способности) хрусталика наибольший в молодом возрасте, когда максимальная величина диапазона аккомодации достигает 14 D. При максимальной кривизне хрусталика точка ближайшего максимума видения глаза находится на расстоянии 7 см. С возрастом хрусталик теряет эластичность и после 45-50 лет диоптрическая сила его начинает снижаться так, что в старости диапазон аккомодации может составлять лишь около 2 D. При этом ближайшая точка нормального видения постепенно отодвигается, и способность ясно видеть близкие предметы утрачивается. Это явление именуетсяпресбиопией (старческой дальнозоркостью).
Цилиарная мышца иннервируется парасимпатическими волокнами глазодвигательного нерва и при их возбуждении глаз начинает ясно видеть близко расположенные предметы. Стимулируется рефлекс с нейронов фовеальной области коры (зрительное поле 18). Если закапать в глаз лекарственные препараты, блокирующие медиаторную пере-дачу сигналов парасимпатического нерва (атропин), то глаз перестает видеть близкие предметы. В связи с тем, что для видения предметов, находящихся недалеко, цилиарная мышца должна сокращаться, то, например, при продолжительном чтении глаза начинают "уставать".