

Физиология микроорганизмов
Микроорганизмам, как и всем живым существам, присущи процессы питания, дыхания, роста и размножения. Однако эти процессы у микробов характеризуются своеобразием и рядом особенностей. Микробы занимают особое место среди других живых существ: они способны использовать в качестве питательных веществ как неорганические, так и разнообразные органические соединения; могут существовать и размножаться в аэробных и анаэробных условиях; длительно сохраняются во внешней среде с помощью спор; обладают исключительной приспособляемостью к меняющимся факторам окружающей среды.
Дыхание бактерий
Сущность процесса дыхания бактерий заключается в совокупности биохимических реакций, в ходе которых идет образование АТФ, без которого невозможен процесс метаболизма, протекающего с затратой энергии. АТФ является универсальным переносчиком химической энергии между процессами, выделяющими энергию, и реакциями, их использующими. При дыхании — процессе биологического окисления бактерий — потребляются те же соединения, что и на построение отдельных структурных компонентов клетки, но в первую очередь — сахара, спирты, органические кислоты, жиры и т. д.
Большая часть бактерий использует в процессе дыхания свободный кислород. Такие микроорганизмы получили название аэробные (от аег — воздух). Аэробный тип дыхания характеризуется тем, что окисление органических соединений происходит при участии кислорода воздуха с освобождением большого количества калорий. Молекулярный кислород выполняет роль акцептора водорода, образующегося при аэробном расщеплении этих соединений.
Примером может служить окисление глюкозы в аэробных условиях, которое приводит к выделению большого количества энергии:
С 6Н12О6
+ 602
6С02+6Н20
+ 688,5 ккал.
6Н12О6
+ 602
6С02+6Н20
+ 688,5 ккал.
Процесс анаэробного дыхания микробов заключается в том, что бактерии получают энергию при окислительно-восстановительных реакциях, при которых акцептором водорода является не кислород, а неорганические соединения — нитрат или сульфат.
Многие бактерии могут существовать в аэробных и анаэробных условиях. Такие микроорганизмы получили название факультативных (необязательных) анаэробов.
Например, стафилококки, кишечная палочка и другие факультативные анаэробы имеют полный набор дыхательных ферментов, обеспечивающих им существование как в кислородной, так и в бескислородной среде. Факультативные анаэробы обладают так называемым нитратным дыханием, так как образующийся при окислении органических соединений нитрат (акцептор водорода) восстанавливается до молекулярного азота и аммиака.
Обязательные анаэробы
Облигатные (обязательные) анаэробы могут существовать лишь в строго анаэробных условиях. Среди патогенных — это возбудители столбняка, газовой гангрены, ботулизма. Облигатные анаэробы при окислении органических соединений образуют сульфат, который восстанавливается до сероводорода, поэтому облигатное дыхание называют еще сульфатным.
Для нейтрализации токсичных форм кислорода микроорганизмы, способные существовать в его атмосфере, имеют защитные механизмы. У облигатных аэробов и факультативных анаэробов накоплению закисного радикала О2- препятствует фермент супероксиддисмутаза, расщепляющая закисный радикал на перекись водорода и молекулярный кислород. Перекись водорода у этих бактерий разлагается каталазой на воду и молекулярный кислород.
В присутствии кислорода рост облигатных анаэробов прекращается. Это связано с тем, что жизнь в аэробных условиях приводит к тому, что конечным продуктом окисления органических соединений оказывается перекись водорода, а поскольку анаэробы не продуцируют фермент каталазу, расщепляющую перекись водорода, то она накапливается и оказывает токсическое действие на анаэробные бактерии.
При выращивании анаэробов в лабораторных условиях используют специальные приборы — анаэростаты, из которых кислород воздуха удаляется или заменяется другим инертным газом. Бескислородные условия можно создать также кипячением среды или химическими веществами, активно поглощающими кислород из пространства, куда помещены чашки и пробирки с посевами.
Образование энергии (АТФ) наблюдается также при процессах брожения, осуществляемых разнообразными микроорганизмами. Особенность брожения заключается в том, что органические соединения одновременно служат как донаторами электронов (при их окислении), так и акцепторами (при их восстановлении). Брожение происходит в отсутствие кислорода, в строго анаэробных условиях. Основными соединениями брожения являются углеводы. В зависимости от участия определенного микроба и от конечных продуктов расщепления углеводов различают спиртовое, молочнокислое, уксуснокислое, маслянокислое и другие виды брожения.
Освобождение энергии при анаэробных процессах значительно меньшее, например, при брожении глюкозы дрожжами образуется спирт и всего 31,2 ккал.
Спиртовое брожение встречается, в основном, у дрожжей. Конечными продуктами являются этанол и СО2. Сбраживание глюкозы происходит в анаэробных условиях. При доступе кислорода процесс брожения ослабевает, на смену ему приходит дыхание. Подавление спиртового брожения кислородом называется эффектом Пастера. Спиртовое брожение используется в пищевой промышленности : хлебопекарной, виноделии.
Молочнокислое брожение. Различают два типа: гомоферментативное и гетероферментативное.
При гомоферментативном типе расщепление глюкозы происходит гликолитическим распадом (ФДФ-путь) Водород от восстановленного НАД передается на пируват при помощи лактатдегидрогеназы, при этом образуется молочная кислота. Гомоферментативное брожение происходит у S.pyogenes, E.faecalis, S.salivarius у некоторых видов рода Lactobacillus.
Гетероферментативное молочнокислое брожение присутствует у бактерий, у которых отсутствуют ферменты ФДФ-пути : альдолаза и триозофосфатизомераза. Расщепление глюкозы происходит с образованием фосфоглицеринового альдегида (ПФ-путь), который превращается в пируват по ФДФ-пути и в последующем восстанавливается в лактат. Дополнительными продуктами этого типа брожения являются также этанол, уксусная кислота. Гетероферментативное молочнокислое брожение встречается у представителей родов Lactobacillus и Bifidobacterium.
Муравьинокислое (смешанное) брожение встречается у представителей семейств Enterobacteriaceae, Vibrionaceae. В зависимости от продуктов брожения, различают два типа процессов:
Расщепление пирувата с образованием ацетилкофермента А и муравьиной кислоты, которая в свою очередь, может расщепляться на двуокись углерода и молекулярный водород. Другими продуктами брожения, образующимися через цепь реакций, являются этанол, янтарная и молочная кислота. Сильное кислотообразование можно выявить реакцией с индикатором метил-рот, который меняет окраску в сильно кислой среде.
Образуется целый ряд кислот, однако главным продуктом брожения являются ацетоин и 2,3-бутандиол. Ацетоин образуется из двух молекул пирувата с последующим двукратным декарбоксилированием. При последующем восстановлении ацетоина образуется 2,3-бутандиол. Эти вещества при взаимодействии с а-нафтолом в щелочной среде вызывают образование окраски бурого цвета, что выявляется реакцией Фогеса-Проскауэра, используемой при идентификации бактерий.
Маслянокислое брожение. Масляная кислота, бутанол,ацетон,изопропанол и ряд других органических кислот, в часности уксусная, капроновая, валерьяновая, пальмитиновая, являются продуктами сбраживания углеводов сахаролитическими строгими анаэробами.
Аэротолерантные микроорганизмы не используют кислород для получения энергии, но могут существовать в его атмосфере. К этой группе относятся молочно-кислые бактерии, получающие энергию гетероферментативным молочнокислым брожением.
Химический состав микробов
Клетки микробов состоят из воды, белков, углеводов, жиров и минеральных веществ.
Вода является основным по содержанию компонентом бактериальной клетки (до 80—90%). Она находится в свободном состоянии как самостоятельное соединение и связана с другими компонентами клетки. Свободная вода необходима бактериальной клетке для осуществления биохимических процессов. Она является универсальной дисперсионной средой для коллоидов и растворителем для кристаллоидов. Высушивание — удаление воды из клетки — ведет к замедлению жизненных процессов.
Белки составляют 40—80% сухой массы бактерий, большая часть которых представляет собой сложные белки — нуклеопротеиды, хромопротеиды. Бактерии могут содержать до 2000 различных белков, составляющих структуру клетки и участвующих в метаболических реакциях. Количественное и качественное разнообразие белковых соединений придает бактериям видовую специфичность, определяет отношение к окрашиванию, обеспечивает вирулентность, токсигенность, антигенные и иммуногенные свойства. Большая часть белков выполняет ферментативные функции клетки.
Нуклеиновые кислоты в бактериях выполняют те же функции, что и в клетках животного происхождения: молекула ДНК (нуклеоид) обеспечивает наследственные свойства, рибонуклеиновые кислоты (информационная, транспортная и рибосомальная) выполняют соответствующие функции. На долю последней приходится около 80% всей бактериалыюй РНК.
Углеводы в бактериальной клетке находятся в виде простых веществ (моно- и дисахариды) и комплексных соединений. Полисахариды выполняют пластическую функцию, входя в структуру клетки; играют основную роль в обеспечении энергией процессов клеточного метаболизма. Часть внутриклеточных полисахаридов — крахмал, гликоген и др. — являются запасными питательными веществами.
Липиды являются необходимыми компонентами цитоплазматической мембраны и клеточной стенки. В некоторых бактериях они выполняют роль запасных питательных веществ.
Колебания содержания липидов в бактериях очень велики — от 1,5 до 40% — и связаны с видовой принадлежностью. Количество липидов зависит и от физиологического состояния клетки, от возраста культур. Наиболее богаты липидами микобактсрии туберкулеза — до 40%. Бактериальные липиды представлены в основном фосфолипидами, жирными кислотами, глицеридами.
Органические вещества бактерий не находятся в клетке в виде отдельных компонентов, а представляют собой сложные соединения с большой молекулярной массой.
Минеральные вещества — фосфор, калий, магний, сера, железо, кальций, йод, цинк, молибден и др. — входят в состав различных клеточных структур бактерий. Они необходимы для регулирования осмотического давления, рН, окислительно-восстановительного потенциала, для активации ферментов. Общее содержание минеральных веществ составляет от 2 до 30% сухой массы бактериальной клетки.
В практических бактериологических лабораториях широко применяют микро- и экспресс-методы для ориентировочного изучения биохимических свойств микроорганизмов. Для этой цели существует множество тест-систем. Наиболее часто используют систему индикаторных бумаг (СИБ). СИБы представляют из себя диски фильтровальной бумаги, пропитанные растворами сахаров или других субстратов в сочетании с индикаторами. Такие диски опускают в пробирку с выросшей в жидкой питательной среде культурой. По изменению цвета диска с субстратом судят о работе фермента. Микро-тест системы для изучения идентификации энтеробактерий представлены одноразовыми пластиковыми контейнерами со средами, содержащими различные субстраты, с добавлением индикаторов. Посев чистой культуры микроорганизмов в такие тест-системы позволяет быстро выявить способность бактерий утилизировать цитраты, глюкозу, сахарозу, выделять аммиак, индол, разлагать мочевину, лизин, фенилаланин и т.д.
Питание бактерий
Своеобразие процесса питания бактерий состоит в том, что
поступление питательных веществ в клетку происходит по всей поверхности, которая очень велика по сравнению с общей величиной бактерии.
Второй особенностью является необыкновенная быстрота метаболических процессов
третьей — высокая адаптация к меняющимся условиям среды.
Типы питания. Разнообразие условий существования микробов обусловливает различные типы питания. Они определяются на основании усвоения двух из четырех необходимых органогенов — углевода и азота. Источником водорода и кислорода служит вода.
По усвоению углерода бактерии можно разделить на два типа:
аутотрофы (литотрофы)
гетеротрофы. (органотрофы)
Аутотрофы (от autos — сам, trophe — пища) способны получать углерод из неорганических соединений и даже из углекислоты. Энергию, необходимую для синтеза органических веществ, аутотрофы получают при окислении минеральных соединений. К аутотрофным бактериям относятся нитрифицирующие (находящиеся в почве), серобактерии (живущие в теплых источниках с содержанием сероводорода), железобактерии (размножающиеся в воде с закисным железом) и др.
Гетеротроф ы (от heteros— другой, trophe — пища) используют в качестве источника углерода органические соединения. Универсальным источником углерода служат различные углеводы (их часто добавляют в питательные среды), белки и др. Гетеротрофы играют значительную роль в уничтожении различных мертвых органических остатков. Такие бактерии называются сапрофитами (от sapros — гнилой, phyton — растение). Микробы, способные существовать за счет органических соединений организма животных и в клетках растений, получили название паразитических (parasitos — нахлебник). Среди патогенных микроорганизмов выделяют так называемые облигатные паразиты, которые способны жить только в живых клетках или тканях. К таким микробам относятся риккетсии, вирусы и некоторые простейшие.
По способности усваивать азот бактерии делятся также на две группы:
аминоаутотрофы (аминолитотрофы)
аминогетеротрофы. (аминоорганотрофы)
Аминоаутотрофы используют молекулярный азот воздуха. Бактерии этой группы — азотфиксирующие почвенные и клубеньковые бактерии — единственные живые существа, усваивающие свободный азот, принимают активное участие в круговороте азота в природе.
Аминогетеротрофы получают азот из органических соединений — сложных белков. К аминогетеротрофам относятся все патогенные микроорганизмы и большинство сапрофитов.
В настоящее время для аутотрофов применяется также название литотрофы, а для гетеротрофов — органотрофы.
По источникам энергии различают
фототрофы — бактерии, для которых источником энергии является солнечный свет,
хемотрофы — бактерии, которые получают энергию за счет химического окисления веществ.
В зависимости от окисляемого субстрата среди хемотрофных организмов выделяют хемолитотрофы и хемоорганотрофы .
Однако далеко не все соединения, которые необходимы бактериям в биологических процессах, клетка может синтезировать сама. При составлении питательных сред необходимо добавлять вещества, получившие название факторов роста. Это различные витамины, аминокислоты (без которых невозможен синтез белка), пиридиновые и пиримидиновые основания (предшественники нуклеиновых кислот) и др.
Микроорганизмы, нуждающиеся в каком-то одном или нескольких факторах роста, называются ауксотрофными в отличие от прототрофных бактерий, которые в данных соединениях не нуждаются и способны сами их синтезировать.
Механизм питания бактерий
Поступление в бактериальную клетку питательных веществ представляет собой сложный физико-химический процесс, которому способствует ряд факторов: разница в концентрации веществ, величина молекул, их растворимость в воде или липидах, рН среды, проницаемость клеточных мембран и т. д. В проникновении питательных веществ в клетку различают четыре возможных механизма.
Наиболее простой способ — пассивная диффузия, при которой поступление вещества в клетку происходит из-за различия градиента концентрации (разницы концентрации по обе стороны цитоплазматической мембраны). Решающее значение имеет величина молекулы. Очевидно, в мембране есть участки, через которые и возможно проникновение веществ небольших размеров. Одним из таких соединений является вода.
Большинство питательных веществ попадает в бактериальную клетку против градиента концентрации, поэтому в таком процессе должны принимать участие ферменты и может расходоваться энергия.
Одним из таких механизмов является облегченная диффузия, которая происходит при большей концентрации вещества вне клетки, чем внутри. Облегченная диффузия — процесс специфический и осуществляется особыми мембранными белками, переносчиками, получившими название п е р м е а з, так как они выполняют функцию ферментов и обладают специфичностью. Они связывают молекулу вещества, переносят в неизмененном виде к внутренней поверхности цитоплазматической мембраны и высвобождают в цитоплазму. Так как перемещение вещества происходит от более высокой концентрации к более низкой, этот процесс протекает без затраты энергии.
Третий возможный механизм транспорта веществ поучил название активного переноса. Этот прессе наблюдается при низких концентрациях субстрата в окружающей среде и перенос растворенных веществ также в неизмененном виде осуществляется против градиента концентрации. В активном переносе веществ участвуют пермеазы. Поскольку концентрация вещества в клетке может в несколько тысяч раз превышать ее во внешней среде, активный перенос обязательно сопровождается затратой энергии. Расходуется аденозинтрифосфат (АТФ), накапливаемый бактериальной клеткой при окислительно-восстановительных процессах.
при четвертом возможном механизме переноса питательных веществ наблюдается транслокация радикалов — активный перенос химически измененных молекул, которые в целом виде не способны проходить через мембрану. В переносе радикалов участвуют пермеазы.
Синтезируемые в бактериальных клетках соединения выходят из них тремя путями:
Фосфотрансферазная реакция. Происходит при фосфорилировании переносимой молекуды.
Контрансляционная секреция. В этом случае синтезируемые молекулы должны иметь особую лидирующую последовательность аминокислот, чтобы прикрепиться к мембране и сформировать канал, через который молекулы белка смогут выйти в окружающую среду. Таким образом выходят из клетки соответствующих бактерий токсины столбняка, дифтерии и др. молекулы.
Почкование мембраны. Молекулы, образующиеся в клетке, окружаются мембранным пузырьком, который отшнуровывается в окружающую среду.
Ферменты бактерий.
В бактериальной клетке происходят многочисленные реакции, как биосинтетические, направленные на синтез соединений, необходимых для организации структуры бактерии, так и производящие энергию, процессы ассимиляции и диссимиляции. Все эти реакции катализируются соответствующими ферментами. Ферменты являются белками и обладают специфичностью при распознавании соответствующего вещества и последующем превращении его. Большая часть ферментов связана с определенными структурами бактериальной клетки. Так, в цитоплазматической мембране находятся окислительно-восстановительные ферменты, которым принадлежит основная роль в дыхании клетки, ферменты, обеспечивающие доставку питательных веществ, и др. Ферменты, связанные с делением клетки, обнаруживаются в мезосомах, клеточной стенке, в месте образования перегородки.
У бактерий по характеру вызываемых ими превращений обнаруживаются следующие основные группы ферментов:
г и д р о л а з ы, вызывающие расщепление протеинов, углеводов, липидов путем присоединения молекул воды;
оксидоредуктазы, катализирующие окислительно-восстановительные реакции;
трансфера з ы, осуществляющие перенос отдельных атомов, от молекулы к молекуле;
л и а з ы, отщепляющие химические группы негидролитическим путем;
изомеразы, участвующие в углеводном обмене;
л и г а з ы, способствующие биосинтетическим реакциям клетки.
Ферменты бактерий классифицируются на экзоферменты и эндоферменты. Экзоферменты выделяются бактериальной клеткой в окружающую среду для внеклеточного переваривания. Этот процесс осуществляется с помощью гидролаз, которые расщепляют макромолекулы питательных веществ до простых соединений — глюкозы, аминокислот, жирных кислот. Такие соединения могут свободно проходить через оболочку клетки и с помощью пермеаз передаваться в цитоплазму клетки для участия в метаболизме, являясь источниками углерода и энергии. Некоторые экзоферменты выполняют защитную функцию, например, пенициллиназа, выделяемая многими бактериями, делает клетку недосягаемой для антибиотика — пенициллина.
Эндоферменты катализируют метаболические реакции, происходящие внутри клетки.
Ферменты бактерий классифицируются также на конститутивные и индуцибельные. Конститутивными называются такие ферменты, которые синтезируются клеткой независимо от наличия субстрата в среде, индуцибельные ферменты образуются бактериями только при наличии в среде соответствующего индуцирующего соединения, т. е. субстрата данного фермента. Например, в геноме кишечной палочки заложена способность разлагать лактозу, но только при наличии в среде лактозы клеткой синтезируется фермент, катализирующий ее гидролиз.
Известны также ферменты, которые получили название аллостерических . Кроме активного центра у них имеется регуляторный или аллостерический центр, который в молекуле фермента пространственно разделен с активным центром. Аллостерическим (от греч. allos - иной, чужой) он называется потому, что молекулы, связывающиеся с этим центром, по строению ( стерически ) не похожи на субстрат, но оказывают влияние на связывание и превращение субстрата в активном центре, изменяя его конфигурацию. Молекула фермента может иметь несколько аллостерических центров. Вещества, связывающиеся с аллостерическим центром, называют аллостерическими эффекторами. Они влияют через аллостерический центр на функцию активного центра: или облегчают ее, или затрудняют. Соответственно аллостерические эффекторы называются положительными (активаторы) или отрицательными (ингибиторы). Аллостерические ферменты играют важную роль в тонкой регуляции метаболизма бактерий. Поскольку практически все реакции в клетке катализируются ферментами, регуляция метаболизма сводится к регуляции интенсивности ферментативных реакций.
Патогенные бактерии обладают наряду с ферментами обмена также ферментами агрессии, являющимися факторами вирулентности. К таким ферментам относятся
гиалуронидаза,
дезоксирибонуклеаза,
коллагеназа,
н е й р а м и и и д аза, и др.
Гиалуронидаза стрептококков, например, расщепляет гиалуроновую кислоту в мембранах клеток соединительных тканей макроорганизма , что способствует распространению возбудителей и их токсинов в организме, обуславливая высокую инвазивность этих бактерий.
Плазмокоагулаза является главным фактором патогенности стафилококков, так как участвует в превращении протромбина в тромбин, который вызывает образование фибриногена, в результате чего каждая бактерия покрывается пленкой, предохраняющей ее от фагоцитоза.
Ферменты бактерий обладают высокой специфичностью, и именно это свойство широко используется при идентификации и определении видов микроорганизмов. Наибольшее значение имеет определение сахаролитических (ферментация сахаров) и протеолитических (разложение белков) свойств.
Споры
Одной из особенностей микроорганизмов является их способность к спорообразованию. Споры образуются при неблагоприятных условиях существования (высушивание, недостаток питательных веществ, изменение рН среды и т. д.), причем из одной клетки формируется только одна спора. Таким образом, образование спор не связано с процессом размножения, а является своеобразным приспособлением к переживанию в неблагоприятных условиях. По принятой номенклатуре спорообразующие аэробы носят название бацилл, а спорообразующие анаэробы — клостридии.
Процесс спорообразования проходит ряд стадий, в течение которых в определенном месте клетки цитоплазма, нуклеоид, рибосомы концентрируются, уплотняются, покрываются мембраной, а затем плотной, плохо проницаемой многослойной оболочкой, включающей кальциевые соли дипиколиновой кислоты, обусловливающей термоустойчивость спор. Споры длительное время могут сохраняться в покое, оставаясь жизнеспособными. Так, в почве споры патогенных микроорганизмов (возбудителя сибирской язвы, столбняка и др.) могут сохраняться десятками лет. При попадании в благоприятную среду споры очень быстро прорастают — из 1 споры возникает 1 бактериальная клетка, которая начинает размножаться.
Спорообразование — видовое свойство палочек, а форма и расположение формирующейся споры по отношению к вегетативной части клетки является дифференциально-диагностическим признаком. Форма спор может быть овальной или круглой, расположение центральное (возбудитель сибирской язвы), субтерминальное — ближе к концу палочки (возбудители газовой гангрены, ботулизма) и терминальное — на конце (возбудитель столбняка).

В зрелой споре различимы: центральный, плохо окрашиваемый участок (спороплазма), двухслойная ЦПМ и оболочка споры.
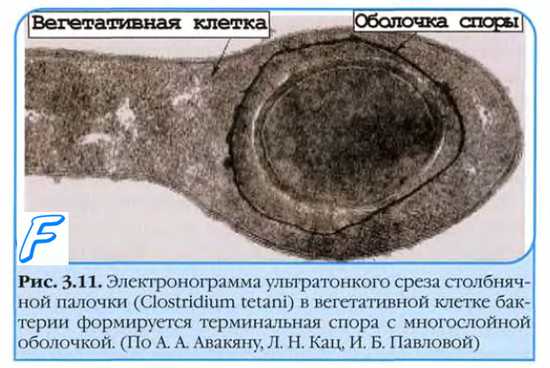
Спороплазма (протопласт споры) включает цитоплазму, бактериальную хромосому, системы белкового синтеза и некоторые другие (например, анаэробного энергообразования).
Оболочка споры двухслойная: пространство между слоями заполняют гликопептидные полимеры, сходные с пептидогликанами, образующие сетчатую структуру (кортекс), проявляющую высокую чувствительность к лизоциму. Внутренний слой (стенка споры) образован пептидогликанами, аналогичными таковым \ вегетирующей клетки. Внешний слой (собственно оболочка) образуют кератиноподобные белковые структуры с низкой проницаемостью.